БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 16. ЕНЕРГЕТИЧНИЙ ОБМІН
16.2.Окисне фосфорилювання АДФ
16.2.1.Механізм спряження окиснення і фосфорилювання
Яким же чином здійснюється спряження цих двох процесів? Найобґрунтованішу відповідь на це питання дає хеміосмотична теорія П. Мітчелла, запропонована ним у 1961 р. Основні положення були підтверджені й розроблені детально спільними зусиллями багатьох дослідників у наступні роки.
Протонний градієнт і електрохімічний потенціал. Перенесення електронів по дихальному ланцюгу від НАДН до кисню супроводжується викачуванням протонів із матриксу мітохондрій через внутрішню мембрану в міжмембранний простір. На цю роботу витрачається частина енергії електронів, які переносяться по ЛПЕ.
Протони, що переносяться з матриксу в міжмембранний простір, не можуть повернутися назад у матрикс, оскільки внутрішня мембрана непроникна для протонів. Отже, створюється протонний градієнт, за якого концентрація протонів у міжмемб- ранному просторі більша, а рН менше, ніж у матриксі. Крім того, кожен протон несе позитивний заряд, і внаслідок цього з'являється різниця потенціалів по обидва боки мембрани: негативний заряд на внутрішньому боці та позитивний - на зовнішньому. У сукупності електричний і концентраційний градієнт Н+ складають градієнт електрохімічного потенціалу ΔμH+ - джерело енергії для синтезу АТФ. Найактивніший транспорт протонів у міжмембранному просторі, який є необхідним для утворення ΔμH, відбувається на ділянках ЛПЕ, що відповідають розміщенню комплексів I, III і IV. Ці ділянки називають пунктами спряження дихання та фосфорилювання (рис. 16.9).
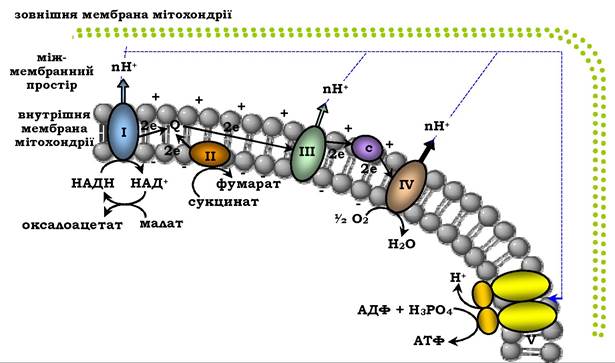
Рис. 16.9. Спряження дихання та синтезу АТФ у мітохондріях:
І - НАДН-дегідрогеназа; ІІ - сукцинатдегідрогеназа;
ІІІ - OН2-дегідрогеназа; IV - цитохрооксидаза; V - АТФ-синтаза. Енергія протонного потенціалу (електрохімічного потенціалу ΔμH) використовується для синтезу АТФ, якщо протони повертаються до матриксу крізь іонні канали АТФ-синтази
Механізм транспорту протонів через мембрану мітохондрій у пунктах спряження недостатньо ясний. Однак установлено, що важливу роль у цьому процесі відіграє KoQ. Детальніше механізм перенесення протонів за участю KoQ вивчений на рівні комплексу III (рис. 16.10).
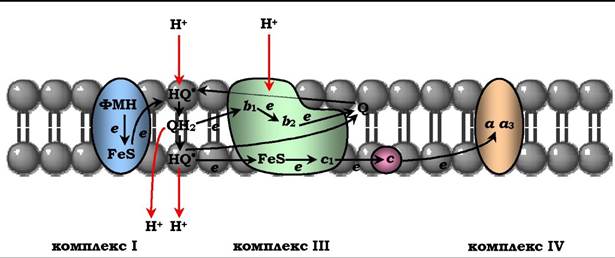
Рис. 16.10. Сполучення перенесення електронів крізь дихальний комплекс ІІІ з транспортом Н+ крізь мембрану: відновлений убіхінон (QH2) взаємодіє з Fe3+ гему b1, і відновлюючи його, вивільняє протон у водну фазу й перетворюється в семихінон (HQ.). Електрон від гему b1 переноситься на Fе3+ гему b2. НQ. віддає другий електрон на FеS-центр, розташований ближче до зовнішньої поверхні мембрани, при цьому другий протон опиняється в міжмембранному просторі, електрон надходить до цитохрому с1, а далі до цитохрому с. Окиснений Q дифундує до внутрішньої поверхня мембрани, де отримує електрон від гему b2 і протон із матриксу, перетворюючись у НQ.. НQ.отримує електрон від комплексу І і протон із матриксу; у мембрані утворюється QH, і весь процес повторюється
KoQ переносить електрони від комплексу I до комплексу III і протони з матриксу в міжмембранний простір, тобто відбуваються своєрідні циклічні перетворення, названі Q-циклами. Донором електронів для комплексу III є відновлений убіхінон (QH2), а акцептором - цитохром с. Останній міститься із зовнішнього боку внутрішньої мембрани мітохондрій; там же розташовується активний центр цитохрому сі, з якого електрони переносяться на цитохром с.
У мембрані існує стаціонарний загальний фонд Q/QH2, з якого кожна молекула QH2 в одному циклі забезпечує перенесення протонів із матриксу в міжмембранний простір і електронів, які врешті-решт надходять до кисню. На роботу, спричинену при викачуванні протонів, витрачається частина вільної енергії, що вивільняється при перенесенні електронів по градієнту редокспотенціалу. Енергія електрохімічного потенціалу (∆μН+) використовується для синтезу АТФ, якщо протони повертаються в матрикс через іонні канали АТФ-синтази.
Будова АТФ-синтази та синтез АТФ. АТФ-синтаза (Н+-АТФаза) - інтегральний білок внутрішньої мембрани мітохондрій. Він розташований у безпосередній близькості до дихального ланцюга. АТФ-синтаза складається з двох білкових комплексів, що позначаються як F0 і F1 (рис. 16.11).
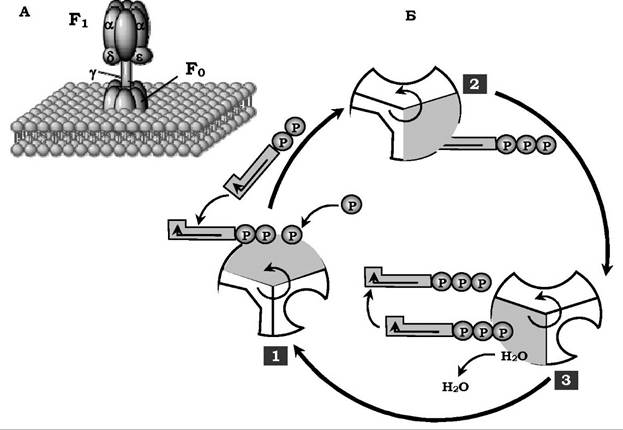
Рис. 16.11. Будова й механізми дії АТФ-синтази:
А - комплекси АТФ-синтази. Б - каталітичний цикл синтезу АТФ:
1 - зв'язування АДФ і Н3РО4; 2 - утворення фосфоангідридного зв'язку АТФ;
3 - вивільнення кінцевого продукту. При кожному перенесенні протонів крізь канал F0 до матриксу всі 3 активні центри каталізують чергову фазу циклу. Енергія електрохімічного потенціалу витрачається на обернення стрижня, у результаті відбуваються циклічні зміни конформації а- і β-субодиниць і синтез АТФ
Гідрофобний комплекс F0 занурений у мембрану. Він виконує роль основи, що фіксує АТФ-синтазу в мембрані. Комплекс F0 складається з декількох субодиниць, котрі утворюють канал, яким протони переносяться в матрикс.
Комплекс F1 виступає в мітохондріальний матрикс. Він складається з дев'яти субодиниць (3а, 3β, γ, ε, δ). Субодиниці а і β укладаються попарно й утворюють "голівку"; між а- і β-субодиницями розташовуються три активні центри, в яких відбувається синтез АТФ; γ-, ε-, δ-субодиниці зв'язують комплекс F1 з F0.
Підвищення концентрації протонів у міжмембранному просторі активує АТФ-синтазу. Електрохімічний потенціал ΔμΗ+ змушує протони рухатися по каналу АТФ-синтази в матрикс. Паралельно під дією ΔμΗ+ відбуваються конформаційні зміни в парах а, β-субодиниць білка F1, у результаті з АДФ і Фн утворюється АТФ. Електрохімічний потенціал, який генерується в кожному з трьох пунктів спряження в ЛПЕ, використовується для синтезу однієї молекули АТФ.
Коефіцієнт окисного фосфорилювання. Окиснення молекули НАДН у ЛПЕ супроводжується утворенням трьох молекул АТФ; електрони від ФАД-залежних дегідрогеназ надходять у ЛПЕ на KoQ, минаючи перший пункт спряження. Тому утворюються тільки дві молекули АТФ. Співвідношення кількості фосфорної кислоти (Р), використаної на фосфорилювання АДФ, і атома кисню (О), поглиненого в процесі дихання, називають коефіцієнтом окисного фосфорилювання і позначають Р/ О. Отже, для НАДН Р/О = 3, для сукцинату Р/ О = 2. Ці величини відображають теоретичний максимум синтезу АТФ, фактично ця величина менше.
Дихальний контроль. Окиснення субстратів і фосфорилювання АДФ у мітохондріях міцно сполучені. Швидкість використання АТФ регулює швидкість потоку електронів у ЛПЕ. Якщо АТФ не використовується і його концентрація в клітинах зростає, то припиняється й потік електронів до кисню. З іншого боку, витрата АТФ і перетворення його в АДФ збільшує окиснення субстратів і поглинання кисню. Залежність інтенсивності дихання мітохондрій від концентрації АДФ називають дихальним контролем. Механізм дихального контролю характеризується високою точністю й має важливе значення, оскільки в результаті його дії швидкість синтезу АТФ відповідає потребам клітини в енергії. Запасів АТФ у клітині не існує. Відносні концентрації АТФ/ АДФ у тканинах змінюються у вузьких межах, тоді як споживання енергії клітиною, тобто частота оборотів циклу АТФ-АДФ, може змінюватися значною мірою (у десятки разів).
Загальний вміст АТФ в організмі становить 30-50 г, але кожна молекула АТФ у клітині "живе" менше хвилини. За добу в людини синтезується 40-60 кг АТФ і стільки ж розпадається. Збільшення концентрації АДФ моментально приводить до прискорення дихання та фосфорилювання.