БИОЛОГИЯ Том 1 - руководство по общей биологии - 2004
2. РАЗНООБРАЗИЕ ЖИЗНИ НА ЗЕМЛЕ
2.3. Прокариоты
2.3.3. Размножение
Индивидуальный рост и бесполое размножение
Соотношение поверхность/объем у бактериальных клеток очень велико, что способствует быстрому поглощению питательных веществ из окружающей среды за счет диффузии и активного транспорта. Поэтому в благоприятных условиях бактерии способны расти очень быстро. Рост бактериальных клеток в большой степени зависит от таких факторов среды, как температура, наличие питательных веществ, pH среды и концентрация ионов. Кроме того, облигатным аэробам необходим кислород, а облигатным анаэробам необходимо, чтобы его не было.
Достигнув определенных размеров, диктуемых соотношением объемов ядра и цитоплазмы, бактерии переходят к бесполому размножению путем простого деления, т. е. путем деления на две идентичные дочерние клетки (рис. 2.11). Клеточному делению предшествует репликация ДНК, причем до тех пор, пока процесс репликации не завершится, мезосомы могут удерживать ДНК в определенном положении (рис. 2.5 и 2.6, В). Мезосомы могут прикрепляться и к новым перегородкам, образующимся между дочерними клетками, участвуя каким-то образом в синтезе материала клеточной стенки. У самых быстрорастущих бактерий деление происходит через каждые 20 мин.
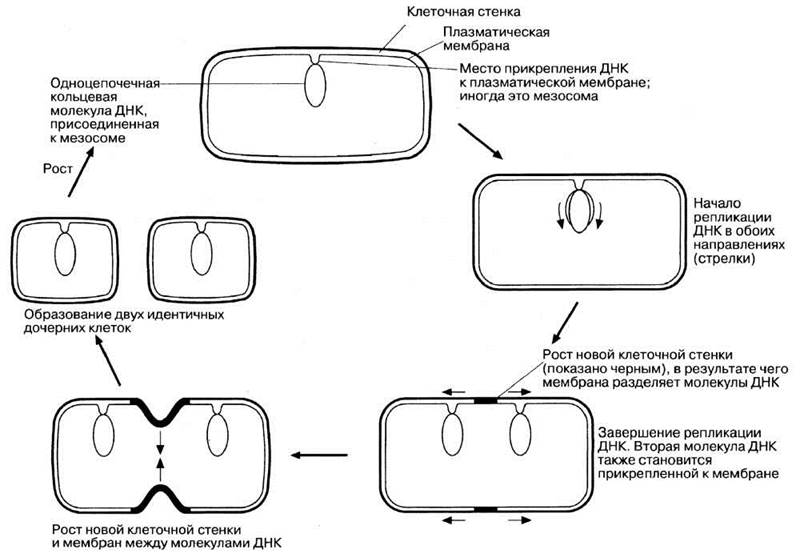
Рис. 2.11. Бесполое размножение бактерий (например, Е.coli) простым делением.
Половое размножение
В 1946 г. у бактерий было обнаружено половое размножение, но в самой примитивной форме. Гамет в данном случае не образуется, однако наиважнейшее событие полового размножения, а именно обмен генетическим материалом, происходит и в этом случае. Этот процесс называется генетической рекомбинацией. Генетическая рекомбинация впервые была обнаружена при изучении E.coli. В норме при наличии в среде достаточного количества глюкозы и неорганических солей E.coli сама синтезирует все необходимые ей аминокислоты. В результате облучения этих бактерий у них иногда возникают случайные мутации. Были выделены два типа мутантов: один, неспособный синтезировать биотин (витамин) и аминокислоту метионин, и другой — не способный синтезировать аминокислоты треонин и лейцин. В среду, не содержавшую всех четырех факторов роста, помещали по 108 клеток каждого мутантного штамма. Теоретически клетки не должны были расти на этой среде. Однако все же было получено несколько сотен колоний (каждая колония возникает из одной исходной клетки), причем оказалось, что в таких клетках имеются все гены, необходимые для образования этих четырех факторов роста. Следовательно, в клетках каким-то образом произошел обмен генетической информацией, но выделить вещество, ответственное за этот процесс, в то время не удалось. В конце концов было установлено (при помощи электронного микроскопа), что клетки E.coli могут непосредственно контактировать друг с другом, т. е. у них может происходит конъюгация (рис. 2.12).
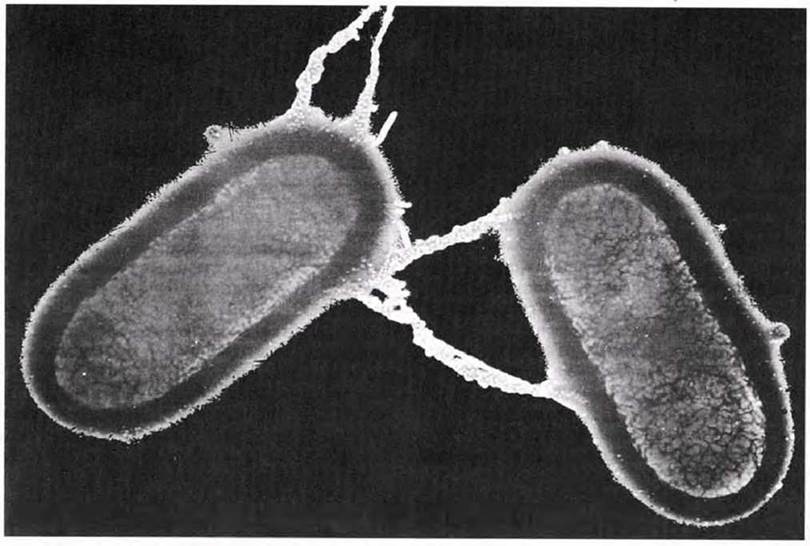
Рис. 2.12. Конъюгация у бактерии. Одна мужская метка (слева) конъюгирует с двумя «женскими» клетками (х 19 475). Второй «женской» клетки на фотографии не видно, так как она находится сверху за пределами фотографии.
Таким образом, при конъюгации происходит перенос ДН К между клетками в результате прямого контакта. Одна клетка в этом случае служит донором («мужская» клетка), другая — реципиентом («женская» клетка). Способность клетки служить донором определяется генами, содержащимися в особой плазмиде, называемой половым фактором или F-фактором (F от англ. fertility — плодовитость). В этих генах закодирован белок специфических пилей, называемых F-пилями или половыми пилями. F-пили участвуют в межклеточном контакте при конъюгации. Пили — структуры полые и предполагается, что именно по этим пилям осуществляется перенос ДНК от донора (F+) к реципиенту (F-). Процесс этот показан на рис. 2.13.
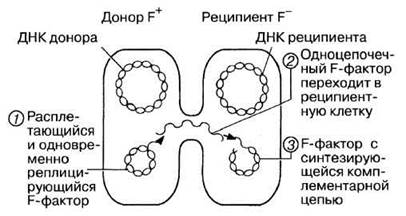
Рис. 2.13. Конъюгация двух бактериальных клеток. Цифрами 1, 2 и 3 обозначены последовательные этапы переноса F-фактора.
Обратите внимание на то, что донорная клетка сохраняет F-фактор, а реципиентная клетка его приобретает и становится F+. Процесс этот протекает медленно, и поэтому прежде чем перенос F-плазмиды завершится, клетка, бывшая изначально F-, успевает реплицироваться один или несколько раз, и в результате в популяции всегда сохраняются F--клетки.
F-фактор вызывает особенно большой интерес и потому, что время от времени, примерно в 1 случае из 100 000, он встраивается в молекулу основной ДНК клетки-хозяина. Тогда при конъюгации происходит перенос не только F-фактора, но и всей остальной ДНК. Этот процесс занимает примерно 90 мин, но клетки могут расходиться и раньше, чем произойдет полный обмен ДНК. Такие штаммы постоянно передают всю или большую часть своей ДНК другим клеткам. Эти штаммы называют Hfr-штаммами (от англ. Н — High — высокая, f — frequency — частота, r — recombination — рекомбинация), потому что донорная ДНК таких штаммов рекомбинирует с ДНК реципиента.