БИОЛОГИЯ Том 1 - руководство по общей биологии - 2004
2. РАЗНООБРАЗИЕ ЖИЗНИ НА ЗЕМЛЕ
2.8. Царство Animalia (животные)
2.8.5. Тип Annelida (аннелиды, или кольчатые черви)
Систематика и характерные признаки кольчатых червей приведены в табл. 2.16, примеры различных аннелид — на рис. 2.54. Две из них, Nereis и Lumbricus рассматриваются ниже более подробно. Аннелиды — целомические животные.
Таблица 2.16. Систематика типа Annelida (аннелиды, или кольчатые черви)
Тип Annelida |
||
Характерные признаки Тело трехслойное, имеется целом Двусторонняя симметрия Метамерия Перед ротовым отверстием расположен простомиум — напоминающий губы вырост переднего сегмента Хорошо выражена кутикула (наружная оболочка) На теле имеются щетинки — волосовидные структуры, образованные хитином и расположенные посегментно (за исключением пиявок)* |
||
Класс Polychaeta (многощетинковые) |
Класс Oligochaeta (малощетинковые, или дождевые черви) |
Класс Hirudinea (пиявки) |
Морские животные |
Обитают в пресных водоемах или во влажной почве |
Эктопаразиты; на переднем и заднем концах тела имеются присоски |
Четко обособлена голова |
Голова обособлена слабо |
Голова обособлена слабо |
На боковых выростах тела, называемых параподиями, имеются многочисленные щетинки* |
Щетинок мало; они расположены парами или по одной; параподий нет* |
Сегментов тела немного и их число постоянное; щетинок и параподий нет* |
Пояска нет |
Имеется поясок, секретируюший материал для кокона, в который откладываются яйца |
Пояска нет |
ПРИМЕРЫ: Arenicola (пескожил) Nereis (нереис) |
ПРИМЕР: Lumbricus (дождевой червь) |
ПРИМЕР: Hirudo (пиявка) |
* Диагностические признаки |
||
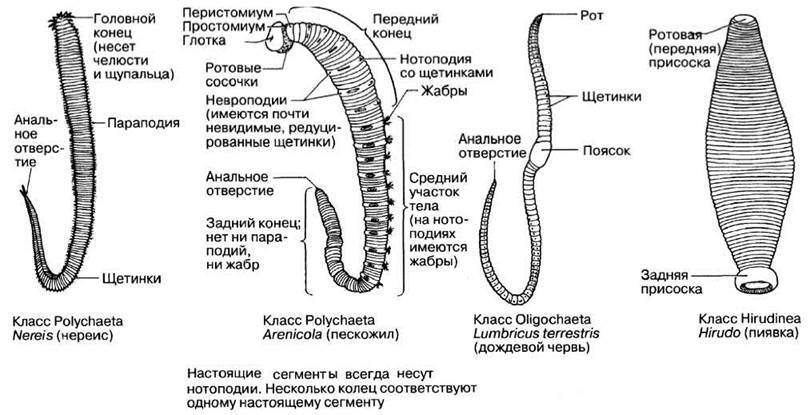
Рис. 2.54. Разнообразие аннелид.
Целомический тип строения тела
Как мы уже видели, у плоских червей мезодерма полностью заполняет пространство между листками эктодермы и энтодермы, образуя плотный средний слой. Строение тела, при котором нет целома, называют ацеломическим (рис. 2.55, A).
У аннелид и более высокоорганизованных групп животных развивается полость тела, называемая целомом. Он возникает как щель в мезодерме в процессе эмбрионального развития. Образующаяся при этом полость заполняется целомической жидкостью, разделяя мезодерму на два листка — соматический, обращенный наружу, и внутренностный, обращенный внутрь (рис. 2.55, Б). Соматическая мезодерма, соединившись с эктодермой, образует стенку тела. Внутренностная мезодерма, соединяясь с энтодермой, образует мускульную стенку кишечника. Таким образом, целом отделяет стенку тела от стенки кишечника.
Основная часть мезодермы, выстилающая целом, развивается в мышцы; мышцы, составляющие стенку тела, обеспечивают локомоцию животного, а мышцы стенки кишечника — перистальтику последнего. Транспорт веществ от стенки тела к стенке кишечника (и наоборот) осуществляется по сосудам хорошо развитой кровеносной системы. Обратите внимание, что полость кишечника находится в энтодерме. Выстилка целома называется брюшиной. Участки брюшины, соединяющие стенку кишечника со стенкой тела называются брыжейками. Любые органы, выдающиеся в целом, например репродуктивные или выделительные органы, удерживаются брюшиной (рис. 2.55, Б).
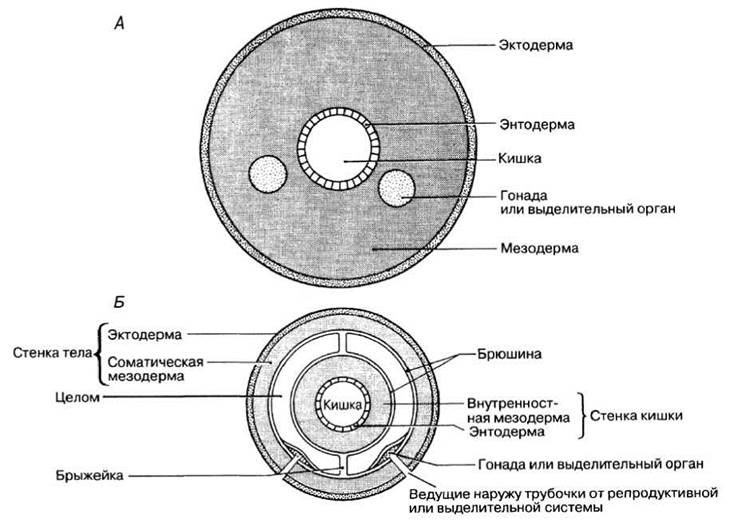
Рис. 2.55. А. Поперечный срез обобщенного ацеломического животного. Б. Поперечный срез обобщенного целомического животного.
Биологическое значение целома
1. Поскольку целом отделяет кишечник от стенки тела, движения мышц стенки тела, ассоциированные с локомоцией животного, способны происходить независимо от движений кишечной стенки (перестальтика), проталкивающих пищу через кишечник и способствующих ее перемешиванию. В результате интенсивность локомоции животного возрастает, и происходит дифференцировка различных частей кишечника, способных выполнять различные функции. В желудке, например, происходит перемешивание пищи, что в свою очередь дает возможность разнообразить диету.
2. Целом — полость, в которой те или иные органы могут расти, развиваться и функционировать независимо друг от друга.
3. Наличие целома способствует увеличению размеров тела и сложности его строения, что порождает дополнительные проблемы, связанные с процессами транспорта и координации. Пища, например, переваривается в кишечнике, но клетки тела отделены от кишечника целомом. Сходным образом, кишечник находится на некотором расстоянии от поверхности тела, через которую происходит газообмен. По мере увеличения размеров тела увеличиваются и перечисленные проблемы, как уже говорилось в разд. 2.8.3, и в результате возникает необходимость в системе транспорта. Поэтому у всех целомических животных появилась сосудистая кровеносная система, представляющая собой систему трубок. Кровь — это жидкая ткань, циркулирующая по всему телу животного за счет сокращения мышечных волокон, расположенных в стенках кровеносных сосудов или за счет работы сердца. Благодаря наличию в сосудах и в сердце клапанов кровь течет лишь в одном направлении. Более высокая сложность требует и более сложной координации, а следовательно, и более сложной нервной системы. Одним из последствий этого направления эволюции явилась цефализация, т. е. развитие головы (см. также ниже — размеры тела и соотношение площадь его поверхности: объем).
4. У аннелид целом несет и дополнительную специализированную функцию — функцию гидростатического, т. е. жидкого скелета. Скелет выполняет три главные функции, а именно функции опоры, защиты и локомоции. Поскольку целомическая жидкость, как и всякая другая жидкость, не сжимается, сокращения мышц могут приводит к изменению формы червя, но не его объема. При локомоции определенные части тела становятся попеременно то длинными и тонкими, то короткими и толстыми — в зависимости от того, какая группа мышц оказывает давление на целомическую жидкость. Защитная роль целома обеспечивается способностью жидкости быстро и равномерно распределять внешнее давление по всем направлениям.
5. Целомическая жидкость может участвовать в транспорте питательных веществ, продуктов обмена и респираторных газов, хотя главную роль в выполнении этих функций играет система кровеносных сосудов.
Метамерия
Еще одним эволюционным преимуществом в развитии целомических животных явилась метамерия, или сегментация, — уровень организации, при котором тело животного поделено поперечными перегородками на несколько одинаковых частей, или сегментов. Иными словами, тело животного представляет собой длинный ряд расположенных друг за другом одинаковых сегментов. Сегментация начинается в мезодерме, но обычно охватывает и мезодермальные, и эктодермальные участки тела.
Наиболее четко метамерия проявляется у кольчатых червей (аннелид), у которых это деление хорошо видно снаружи (перетяжки по всей длине тела). Внутренние сегменты отделяются друг от друга перегородками (септами), проходящими через целом. В каждом сегменте имеется свой собственный набор мышц, кровеносных сосудов, нервных клеток, а у некоторых групп и репродуктивных органов. Однако даже у аннелид сегменты не полностью независимы друг от друга, поскольку и нервная и особенно кровеносная системы должны проходить по всей длине тела животного.
Как только сегментация и основной план строения каждого сегмента эволюционно закрепились, появилась возможность для дальнейших эволюционных изменений в пределах отдельных сегментов или их небольших групп, а также возможность для более узкой специализации и разделения труда у различных частей тела. Это происходит несколькими путями. Различные функции могут выполняться разными сегментами; возможно также слияние сегментов, как это произошло при цефализации, когда несколько сегментов сливаются для формирования головы; возможна даже потеря некоторых сегментов, что наблюдается у членистоногих. Как мы увидим далее (разд. 2.8.6), у членистоногих число сегментов уменьшилось, а сегментация внешне проявляется не так отчетливо, например в головогруди у ракообразных. Однако внутренняя сегментация у членистоногих выражена также отчетливо, как и у аннелид. У хордовых (разд. 2.8.9) наружная сегментация утрачена, но некоторые системы все еще сохраняют четкую сегментацию, например, миотомы (мышечные блоки) у зародышей и спинальные нервы.
Размеры и соотношение площадь поверхности: объем
Организмы с относительно высоким отношением площадь поверхности: объем могут полагаться на диффузию для удовлетворения своих транспортных нужд. Кислород, питательные вещества и продукты обмена, такие как диоксид углерода, могут диффундировать достаточно быстро, чтобы обеспечить выживание организма без каких- либо специальных транспортных систем. Диффузия, однако, может обеспечить перенос веществ лишь на короткие расстояния. Между тем по мере увеличения размеров какого-либо тела объем увеличивается быстрее, чем площадь его поверхности, и в результате отношение площадь поверхности: объем уменьшается. Это легко проиллюстрировать на примере куба (рис. 2.56), но тот же принцип действует и в случае сферических тел, клеток и целых живых организмов. У плоских живых организмов при увеличении объема площадь поверхности остается высокой, что характерно для плоских червей. И у этих организмов диффузия вполне может удовлетворить все транспортные потребности. Однако целомическим животным необходимы специализированные системы для газообмена и транспортные системы.
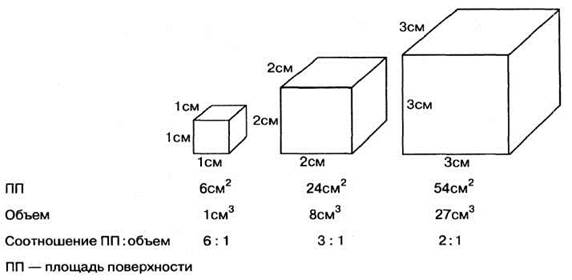
Рис. 2.56. Влияние увеличивающихся размеров тела на соотношение площадь поверхности: объем.
Класс Polychaeta
Пескожил (Arenicola) обитает в приливно-отливной зоне, прорывая норы в песчаном или мягком илистом грунте (рис. 2.54). Нереис (Nereis) (рис. 2.57) обитает в эстуариях либо под камнями либо роет норки в илистом или илисто-песчаном грунте.
Сегментация тела Nereis отчетливо видна снаружи. Все сегменты, за исключением самых передних и задних, одинаковы. По бокам каждого сегмента расположены выросты, называемые параподиями. Верхняя ветвь параподии называется нотоподией, нижняя — невроподией (рис. 2.57). От обеих этих структур (нотоподии и невроподии) отходит по две группы волосковидных щетинок. На каждой параподии имеются еще два дополнительных выроста — игловидные структуры, называемые спинным и брюшным усиками. Через поверхность параподии, обильно снабжаемой кровью, осуществляется газообмен. Nereis ползает, используя параподии как ласты. Этот червь может также и плавать благодаря согласованной работе параподий и мышц кожномускульного мешка, способного изгибаться. Тело Nereis окружено тонкой кутикулой. Пищеварительный канал тянется вдоль всего тела — от ротового отверстия до анального. Глотка у Nereis выдвижная. После того как червь проглотит жертву, глотка снова втягивается.
У Nereis имеется оформленная голова (рис. 2.57); такая четко выраженная цефализация типична для многощетинковых червей, но не для других аннелид. Голова состоит из двух отделов переднего и заднего. Передний отдел, простомиум, образован первым сегментом, а задний — перистомиум, — вторым сегментом. На простомиуме расположена пара чувствительных щупалец (на дорсальной стороне) и две пары глаз, а от вентрально-латеральных участков отходит пара мясистых «щупиков», наделенных чувством осязания. Рот расположен между двумя отделами головы. От одного из них — перистомиума — отходят четыре пары длинных гибких усиков, также наделенных чувством осязания и несущих, кроме того, хеморецепторы, воспринимающие различные химические вещества и заменяющие животному органы вкуса и обоняния.
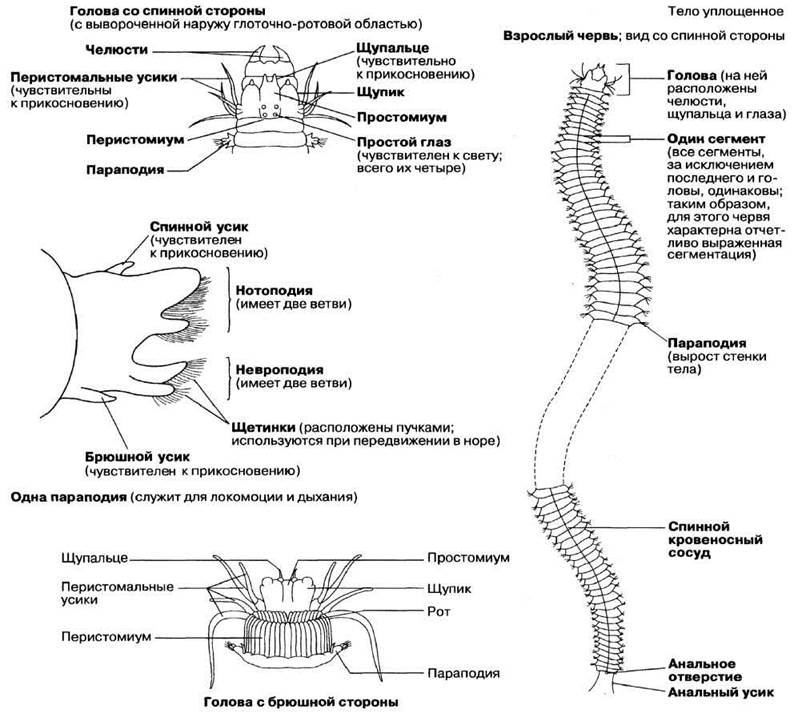
Рис. 2.57. Hepeuc (Nereis), многощетинковый червь.
Класс Oligochaeta
Дождевой червь (Lumbricus terrestris) — животное с длинным цилиндрическим телом, достигающее в длину приблизительно 12—18 см (рис. 2.58). Передний его конец конусообразный, а задний сплющен в дорсо-вентральном направлении. И хотя дождевой червь живет на суше, он не смог до конца преодолеть все проблемы, связанные с наземным образом жизни. Спасаясь от высыхания, он живет под землей, прорывая нору во влажной почве, и покидает ее лишь по ночам, отправляясь на поиски пищи или полового партнера.
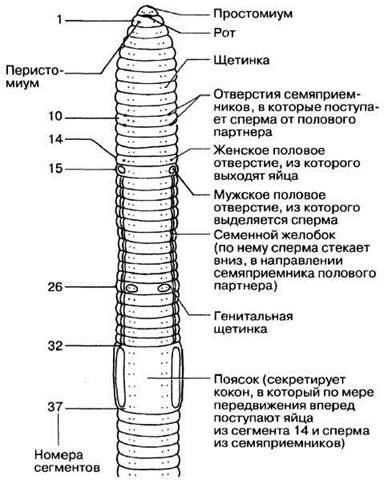
Рис. 2.58. Передний конец дождевого червя (вид с брюшной стороны).
Различия во внешнем строении тела Nereis и дождевого червя обусловлены приспособлением последнего к наземному образу жизни. Тело дождевого червя имеет обтекаемую форму и лишено каких-либо выростов, наличие которых могло бы препятствовать свободному передвижению червя в почве. Простомиум, расположенный над ртом, небольшой и округлый, чувствительных придатков на нем нет. Все сегменты, за исключением первого и последнего, несут по четыре пары щетинок; две из них расположены вентрально (на брюшной стороне), а две дорсо-латерально. Щетинки выходят из расположенных в стенке тела щитинковых мешков. Они могут втягиваться внутрь или выводиться наружу под действием специальных мышц. Щетинки облегчают передвижение червя по земле. Щетинки используются также и для защиты: цепляясь ими за стенки норы, он прочно удерживается в ней. Самые длинные щетинки, расположенные на сегментах 10—15, 26 и 32—37, используются при копуляции. Еще одна структура, используемая при копуляции, — поясок, расположенный на сегментах 32—37. Эпидермис пояска содержит множество железистых клеток, образующих на спинной и боковых поверхностях некоторое утолщение, напоминающее по форме седло. Поясок участвует в процессах копуляции и формирования кокона.
Ротовое и анальное отверстия червя расположены на противоположных концах тела. Дождевой червь питается детритом (фрагментами разлагающегося органического материала), который червь заглатывает вместе с почвой. Подавляющая часть поглощенной почвы проходит через кишечник и затем выбрасывается на поверхность почвы в виде характерных кучек.
Выделяющаяся через спинные поры целомическая жидкость и слизь, секретируемая железистыми клетками эпидермиса, постоянно увлажняют тонкую кутикулу. В результате предотвращается высыхание кутикулы, облегчается газообмен и создается своего рода смазка, облегчающая передвижение червя в почве. Именно через кутикулу происходит газообмен путем диффузии — процесс, зависимый от наличия в эпидермальном слое разветвленной сети капилляров.
Все сегменты, за исключением трех первых и одного последнего, несут по паре нефридиев — канальцев, выполняющих функции выделения и осморегуляции. Они открываются на поверхности тела порами.
Репродуктивная система дождевого червя, как и поведение, связанное с размножением, отличается большой сложностью, что, возможно, объясняется наземным образом жизни и необходимостью предохранять от высыхания гаметы и оплодотворенные яйца. Lumbricus гермафродит (т. е. у каждой особи имеются и мужские и женские репродуктивные органы). Контакт дождевых червей друг с другом происходит очень редко, но когда это происходит, спариваться могут две любые особи, поскольку обе они гермафродиты. В результате происходит взаимообмен спермой и оба червя оплодотворяются.
Половые органы дождевых червей сосредоточены на переднем конце тела. Внешние признаки, связанные с половым размножением, приведены на рис. 2.58. Спаривание и последующая откладка оплодотворенных яиц — процесс, очень сложный. Спаривающиеся особи располагаются рядом друг с другом во взаимно противоположных направлениях, так что голова одного из партнеров оказывается направленной в сторону хвостового отдела другого и наоборот.