БИОЛОГИЯ Том 1 - руководство по общей биологии - 2004
3. ХИМИЧЕСКИЕ КОМПОНЕНТЫ ЖИВОГО
3.6. ДНК и РНК - нуклеиновые кислоты
3.6.3. Структура ДНК
Нуклеиновые кислоты, подобно белкам, обладают первичной структурой (под которой подразумевается их нуклеотидная последовательность) и трехмерной структурой. Интерес к структуре ДНК усилился, когда в начале XX в. возникло предположение, что ДНК, возможно, представляет собой генетический материал. С данными, подтверждающими эту роль ДНК, мы познакомимся в гл. 23.
В начале пятидесятых годов американский химик лауреат Нобелевской премии Лайнус Полинг (Linus Pouling), уже изучивший к тому времени а-спиральную структуру, характерную для многих фибриллярных белков, обратился к исследованию структуры ДН К. которая, по имеющимся в то время сведениям, также представлялась фибриллярной молекулой. Одновременно в Королевском колледже в Лондоне Морис Уилкинс и Розалинда Франклин (Maurice Wilkins, Rosalind Franklin) пытались решить ту же проблему методом рентгеноструктурного анализа. Их исседования требовали долгой и трудоемкой работы по приготовлению чистых препаратов солей ДНК, для которых удавалось получать сложные дифракционные картины (рис. 3.41). С помощью этих картин можно было, однако, выявить лишь общую структуру молекулы ДН К, не столь детализованную, как та, которую давали возможность получить чистые кристаллы белка.
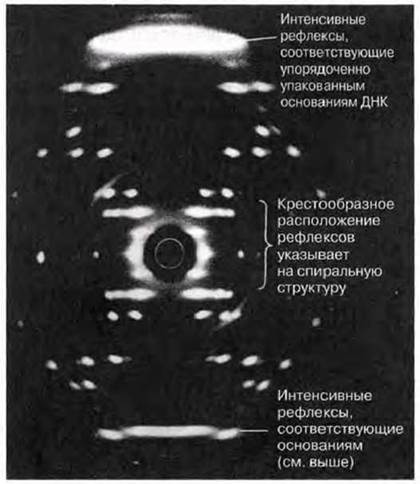
Рис. 3.41. Рентгенограмма нити ДНК. По таким рентгенограммам было впервые сделано заключение о двухспиральной структуре ДНК. (С разрешения J. М. Squire.)
Тем временем Джеймс Уотсон и Фрэнсис Крик (James Watson, Francis Crick) в Кавендишской лаборатории Кембриджского университета избрали иной подход, который в конечном счете и обеспечил успешное решение проблемы. Используя все физические и химические данные, какие оказались в их распоряжении. Уотсон и Крик стали строить пространственные модели ДНК в надежде на то, что рано или поздно им удастся получить достаточно убедительную структуру, согласующуюся со всеми этими данными. История их поисков увлекательно описана Уотсоном в его книге «Двойная спираль».
Два обстоятельства оказались для Уотсона и Крика решающими. Во-первых, они имели возможность регулярно знакомиться с результатами работ Уилкинса и, сопоставляя с его рентгенограммами свои модели, могли таким образом проверять эти модели. Рентгенограммы же Уилкинса убедительно свидетельствовали в пользу спиральной структуры (рис. 3.41) с периодичностью 0,34 нм вдоль оси. Во-вторых, Уотсон и Крик отдавали себе отчет в важном значении закономерностей, касающихся соотношения различных оснований в ДНК. Обнаружил эти закономерности и сообщил о них в 1951 г. Эрвин Чаргафф (Erwin Chargaff). Это открытие, однако, при всей своей важности не привлекло к себе должного внимания. В табл. 3.10 мы приводим некоторые из данных Чаргаффа, дополнив их результатами более поздних исследований.
Таблица 3.10. Относительные количества оснований в ДНК разных организмов
Нуклеотидный состав, мол. % |
||||
Организм |
аденин |
гуанин |
тимин |
цитозин |
Человек Овца Курица Черепаха Лосось Морской еж Саранча Пшеница Дрожжи Eschericha соlli (бактерия) Бактериофаг фХ174 (вирус) |
30,9 29,3 28,8 29,7 29,7 32,8 29,3 27,3 31,3 24,7 24,6 |
19,9 21,4 20,5 22,0 20,8 17,7 20,5 22,7 18,7 26,0 24,1 |
29,4 28,3 29,2 27,9 29,1 32,1 29,3 27,1 32,9 23,6 32,7 |
19,8 21,0 21,5 21,3 20,4 17,3 20,7 22,8 17,1 25,7 18,5 |
3.13. Ознакомьтесь с таблицей. Какой вывод она позволяет сделать о соотношении различных оснований в молекулах ДНК?
Уотсон и Крик задались целью проверить предположение, что молекула ДНК состоит из двух спиральных полинуклеотидных цепей, удерживаемых вместе благодаря спариванию оснований, принадлежащих соседним цепям. Основания удерживаются вместе водородными связями.
3.14. Если эта модель верна, можно ли на основе данных Чаргаффа предсказать, какие основания должны соединяться друг с другом в пары?
Как основания соединяются в пары с помощью водородных связей, показано на рис. 3.42. Аденин спаривается с тимином, а гуанин — с цитозином; АТ-пара соединяется двумя водородными связями, а ГЦ-пара — тремя. Уотсон попытался представить себе такой порядок спаривания оснований и позже вспоминал об этом так: «От радости я почувствовал себя на седьмом небе, ибо тут я уловил возможный ответ на мучившую нас загадку: почему число остатков пуринов в точности равно числу остатков пиримидинов?»1 Уотсон увидел, что при таком сочетании основания оказываются очень точно подогнанными друг к другу и что общий размер и форма двух этих пар оснований одинаковы, так как обе пары содержат по три кольца (рис. 3.42). Водородные связи при других сочетаниях оснований в принципе возможны, но они гораздо слабее. После того как все эти обстоятельства выяснились, можно было наконец приступить к созданию достоверной модели ДНК, той, которая изображена на рис. 3.43-3.45.
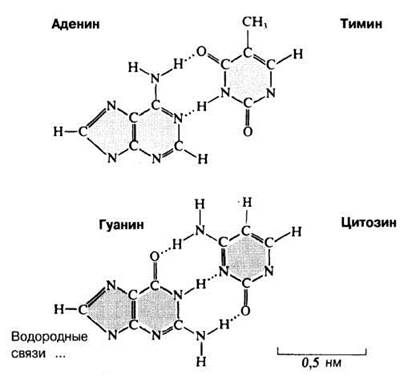
Рис. 3.42. Спаривание оснований — аденина с тимином и гуанина с цитозином.
Строение молекулы ДНК
Уотсон и Крик показали, что ДНК состоит из двух полинуклеотидных цепей. Каждая цепь закручена в спираль вправо, и обе они свиты вместе, т. е. закручены вправо вокруг одной и той же оси, образуя двойную спираль (рис. 3.43). Цепи антипараллельны, т. е. направлены в противоположные стороны. Каждая цепь состоит из сахарофосфатного остова, вдоль которого перпендикулярно длинной оси двойной спирали располагаются основания; находящиеся друг против друга основания двух противоположных цепей двойной спирали связаны между собой водородными связями (рис. 3.44). Сахарофосфатные остовы двух цепей двойной спирали хорошо видны на пространственной модели ДНК (рис. 3.45). Расстояние между сахарофосфатными остовами двух цепей постоянно и равно расстоянию, занимаемому парой оснований, т. е. одним пурином и одним пиримидином. Два пурина занимали бы слишком много места, а два пиримидина — слишком мало для того, чтобы заполнить промежутки между двумя цепями. Вдоль оси молекулы соседние пары оснований располагаются на расстоянии 0,34 нм одна от другой, чем и объясняется обнаруженная на рентгенограммах периодичность. Полный оборот спирали приходится на 3,4 нм, т. е. на 10 пар оснований. Никаких ограничений относительно последовательности нуклеотидов в одной цепи не существует, но в силу правила спаривания оснований эта последовательность в одной цепи определяет собой последовательность нуклеотидов в другой цепи. Поэтому мы говорим, что две цепи двойной спирали комплементарны друг другу.
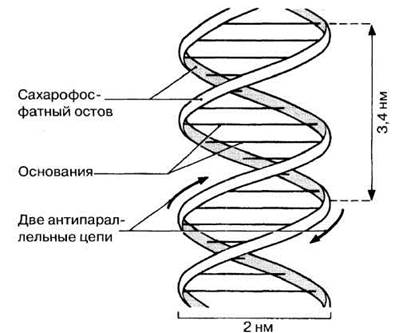
Рис. 3.43. Схематическое изображение структуры ДНК. На один полный оборот спирали приходится 10 пар оснований (расстояние между соседними парами оснований равно 0,34 нм).
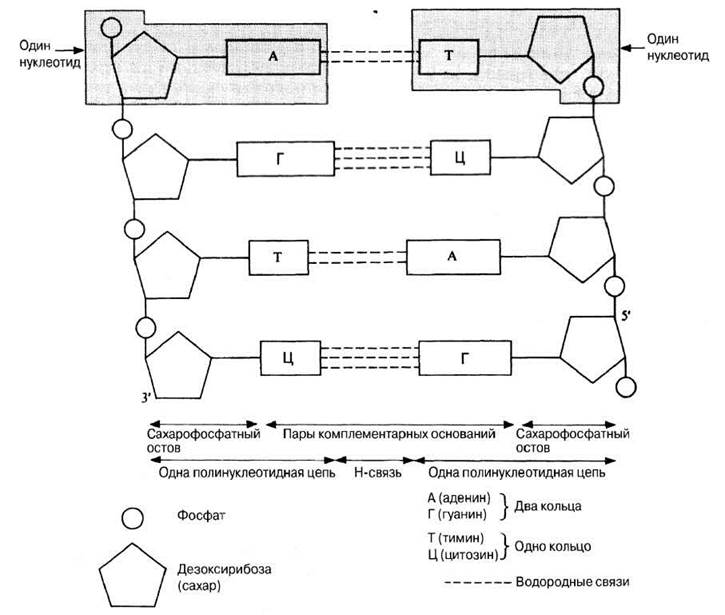
Рис. 3.44. ДНК (схематическое изображение развернутых цепей).
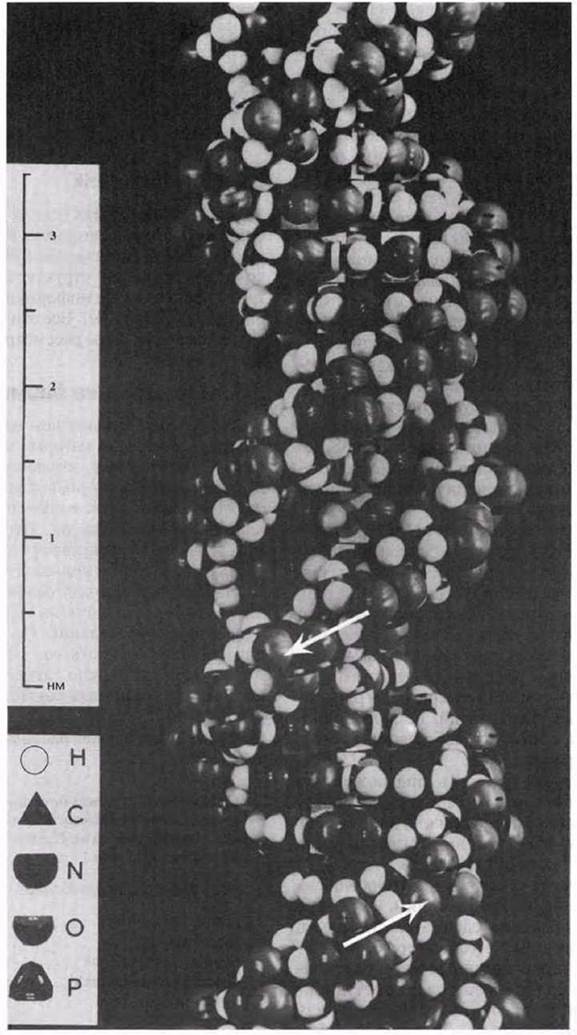
Рис. 3.45. Пространственная модель ДНК. Стрелки указывают направление антипараллельных сахарофосфатных остовов двух полинуклеотидных цепей.
Уотсон и Крик опубликовали сообщение о своей модели ДНК в журнале «Nature» в 1953 г., а в 1962 г. они вместе с Морисом Уилкинсом были удостоены за эту работу Нобелевской премии. В том же году получили Нобелевскую премию Кендрью и Перуц за свои работы по определению трехмерной структуры белков, также выполненные методом рентгеноструктурного анализа. Розалинду Франклин, умершую от рака ранее присуждения этих премий, не включили в число лауреатов, поскольку Нобелевская премия посмертно не присуждается.
Для того чтобы признать предложенную структуру генетическим материалом, требовалось показать, что она способна: 1) нести в себе закодированную информацию и 2) точно воспроизводиться (реплицироваться). Уотсон и Крик отдавали себе отчет в том, что их модель удовлетворяет этим требованиям. В конце своей первой статьи они сдержанно отметили: «От нашего внимания не ускользнуло, что постулированное нами специфическое спаривание оснований сразу же позволяет постулировать и возможный механизм копирования для генетического материала»2. Во второй статье, опубликованной в том же 1953 г., они обсудили выводы, которые следовали из их модели, в генетическом плане (о них мы будем говорить в гл. 23). Это открытие, показавшее, сколь явно структура может быть связана с функцией уже на молекулярном уровне, дало мощный толчок развитию молекулярной биологии.
1 Из «The Double Helix» James D. Watson, Weidenfeld, Nicolson, 1968.
2 Watson J. D., Crick F. Н. С. (1953) Nature, 171, 737.