БИОЛОГИЯ Том 3 - руководство по общей биологии - 2004
23. НЕПРЕРЫВНОСТЬ ЖИЗНИ
23.8. Синтез белка
«ДНК создает РНК, а РНК создает белок»
Из всего того, что было сказано в предыдущих разделах, можно видеть, что единственные молекулы, которые синтезируются под непосредственным контролем ДНК, это белки. Белки могут быть структурными, как кератин и коллаген, или функциональными, как инсулин, фибриноген и, главное, ферменты, ответственные за регуляцию клеточного метаболизма. Именно набор содержащихся в данной клетке ферментов определяет, к какому типу клеток она будет относиться. Таким способом ДНК контролирует жизнедеятельность клетки.
«Инструкции», необходимые для синтеза ферментов и всех других белков, заключены в ДНК, которая почти вся находится в ядре. Однако, как было показано в начале 50-х годов XX в., синтез фактически происходит в цитоплазме и в нем участвуют рибосомы. Стало ясно, что должен существовать какой-то механизм, переносящий генетическую информацию из ядра в цитоплазму. В 1961 г. два французских биохимика Жакоб и Моно, постулировали существование особой формы РНК, которую они назвали матричной РНК (мРНК). Их идея оказалась верной. Последовательность событий, происходящих в процессе синтеза белка, сформулирована в короткой фразе: «ДНК создает РНК, а РНК создает белок».
23.8.1. Роль РНК
РНК содержится во всех живых клетках в виде одноцепочечных молекул. Она отличается от ДНК тем, что содержит в качестве пентозы рибозу (вместо дезоксирибозы), а в качестве одного из пиримидиновых оснований — урацил (вместо тимина). Анализ РНК, содержащейся в клетках, показал, что существуют три типа РНК, участвующих в синтезе белковых молекул. Это матричная, или информационная, РНК (мРНК), транспортная РНК (тРНК) и рибосомная РНК (рРНК). Все три РНК синтезируются непосредственно на ДНК, а количество РНК в каждой клетке находится в прямой зависимости от количества вырабатываемого этой клеткой белка.
23.8.2. Матричная РНК
Как показали анализы, мРНК составляет 3—5% всей РНК клетки. Это одноцепочечная молекула, образующаяся на одной из цепей ДНК в процессе, называемом транскрипцией. При синтезе мРНК копируется только одна цепь молекулы ДНК. Синтез мРНК будет описан позднее. Последовательность оснований в мРНК представляет собой комплементарную копию соответствующей цепи ДНК; длина ее варьирует в зависимости от длины полипептидной цепи, которую она кодирует. Большая часть мРНК находится в клетке лишь короткое время. У бактерий это может быть всего несколько минут, тогда как в развивающихся эритроцитах мРНК может служить матрицей для синтеза гемоглобина в течение нескольких дней.
23.8.3. Рибосомная РНК
Рибосомная РНК составляет примерно 80% всей РНК клетки. Она кодируется генами, находящимися в ДНК нескольких хромосом, расположенных в участке ядрышка, известном под названием ядрышкового организатора. Последовательность оснований в рРНК сходна у всех организмов — от бактерий до высших растений и животных. рРНК содержится в цитоплазме, где она связана с белковыми молекулами, образуя вместе с ними клеточные органеллы, называемые рибосомами (разд. 5.10.4).
На рибосомах происходит синтез белка. Здесь «код», заключенный в мРНК, транслируется в аминокислотную последовательность полипептидной цепи.
23.8.4. Транспортная РНК
Существование транспортной РНК (тРНК) было постулировано Криком и продемонстрировано Хоглендом в 1955 г. У каждой аминокислоты имеется собственная семья молекул тРНК. Они доставляют содержащиеся в цитоплазме аминокислоты к рибосоме. Таким образом, тРНК служит промежуточной молекулой между триплетным кодом в мРНК и аминокислотной последовательностью полипептидной цепи. На долю тРНК приходится примерно 15% всей клеточной РНК; у этих РНК самая короткая полинуклеотидная цепь — в нее входит в среднем 80 нуклеотидов. В каждой отдельной клетке содержится более 20 различных молекул тРНК (идентифицировано уже 60). Все молекулы тРНК имеют сходную основную структуру (см. рис. 23.25).
На 5'-конце молекулы тРНК всегда находится гуанин, а на 3'-конце — последовательность оснований ЦЦА. Последовательность нуклеотидов в остальной части молекулы варьирует и может содержать «необычные» основания, такие как инозин и псевдоурацил. Последовательность оснований в триплете антикодона (рис. 23.25) строго соответствует той аминокислоте, которую переносит данная молекула тРНК. Каждая аминокислота присоединяется к одной из своих специфичных тРНК при участии фермента аминоацил-тРНК-синтетазы. В результате образуется комплекс аминокислота-тРНК, известный как аминоацил-тРНК, в котором энергия связи между концевым нуклеотидом А в триплете ЦЦА и аминокислотой достаточна, чтобы в дальнейшем могла образоваться связь с соседней аминокислотой. Таким образом синтезируется полипептидная цепь.
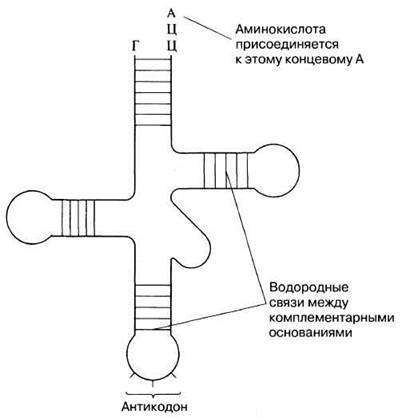
Рис. 23.25. Одна из предложенных моделей строения транспортной РНК (тРНК). Молекула состоит из 80 нуклеотидов, но пар, образованных в результате комплементарного связывания оснований, только 21.
23.8.5. Резюме
Процесс синтеза белка слагается из двух этапов, схематически представленных на рис. 23.26.
1. Транскрипция — синтез мРНК на ДНК. При этом некоторый участок ДНК (ген) копируется с образованием мРНК.
2. Трансляция — перевод последовательности оснований в молекуле мРНК в последовательность аминокислот в молекуле белка.
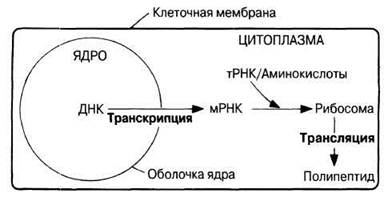
Рис. 23.26. Схема главных этапов белкового синтеза.
23.8.6. Транскрипция
Транскрипцией называют процесс, посредством которого последовательность оснований в одном из участков цепи ДНК «переписывается» в комплементарную последовательность оснований мРНК. Двойная спираль ДНК раскручивается в результате разрыва относительно слабых водородных связей между основаниями двух одиночных цепей ДНК. Только одна из этих цепей может быть выбрана в качестве матрицы для образования комплементарной одиночной цепи мРНК. Эта молекула образуется в результате связывания друг с другом свободных рибонуклеотидов под действием PH К-полимеразы в соответствии с правилами спаривания оснований ДНК и РНК (табл. 23.5 и рис. 23.27).
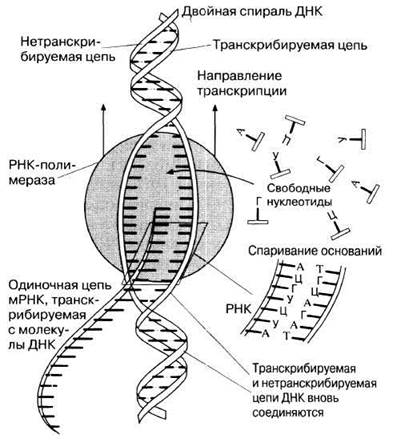
Рис. 23.27. Схема механизма транскрипции. В присутствии РНК-полимеразы двойная спираль ДНК раскручивается в результате разрыва водородных связей между комплементарными основаниями. На образовавшейся одиночной цепи ДНК из свободных рибонуклеотидов строится полинуклеотидная цепь мРНК. Основания мРНК выстраиваются против комплементарных оснований ДНК. (По Е. J. Ambrose, D. М. Easty, Cell Biology, 1977, 2nd ed. Nelson.)
Как именно происходит транскрипция оснований ДНК в основания РНК, было продемонстрировано в опытах с синтетической ДНК, состоявшей из нуклеотидов только одного типа — тимидина (ТТТ...). При введении этой ДНК в бесклеточную систему, содержавшую PHК-полимеразу и все четыре нуклеотида (А, У, Г и Ц), синтезировалась мРНК, содержавшая только комплементарные аденинуклеотиды.
Синтезированные молекулы мРНК выходят из ядра через ядерные поры и несут генетическую информацию рибосомам. После того как в процессе транскрипции образовалось достаточное количество молекул мРНК, РНК-полимераза покидает ДНК, и две цепи ДНК вновь соединяются («молния» застегивается) — восстанавливается двойная спираль.
23.8.7. Трансляция
Трансляцией называют процесс, посредством которого последовательность оснований в молекуле РНК переводится в последовательность аминокислот в полипептидной цепи. Этот процесс происходит на рибосомах. Несколько рибосом присоединяются к молекуле мРНК в виде нити бус, образуя структуру, названную полирибосомой или полисомой. Полисомы можно увидеть с помощью электронного микроскопа (рис. 23.28). Преимущество полисомной организации заключается в том, что при этом становится возможным одновременный синтез нескольких полипептидов (разд. 23.8.3). Каждая рибосома состоит из двух субчастиц — большой и малой — и напоминает по форме грибок (рис. 5.27). Первые два кодона мРНК (в сумме 6 оснований) входят в рибосому, как показано на рис. 23.29, А. Первый кодон связывает молекулу аминоацил-тРНК, содержащую комплементарный ему антикодон и несущую первую аминокислоту (обычно это метионин) синтезируемого полипептида. Затем второй кодон присоединяет к себе молекулу аминоацил-тРНК, содержащую комплементарный ему антикодон (рис. 23.29, А и Б). Функция рибосомы заключается в том, чтобы удерживать в нужном положении мРНК, тРНК и ферменты, участвующие в процессе трансляции, до тех пор, пока между соседними аминокислотами не образуется пептидная связь.
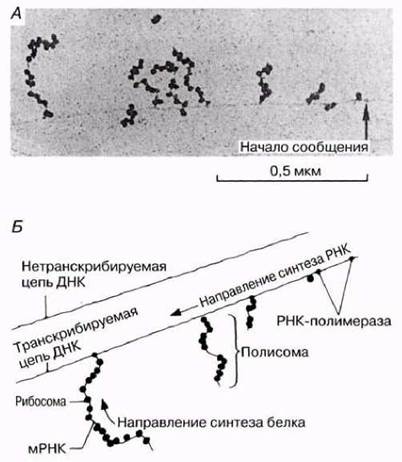
Рис. 23.28. Процесс транскрипции и образования полисомы у бактерий. Обратите внимание на то, что, поскольку у бактерий нет ядра, PHК не должна у них отделяться от ДНК. А. Электронная микрофотография участка бактериальной ДНК, на которой можно видеть стадии образования мРНК и присоединение рибосом. Б. Схематическое изображение структуры, показанной на микрофотографии А.
Как только новая аминокислота присоединилась к растущей полипептидной цепи, рибосома перемещается по мРНК на один кодон. Молекула тРНК, которая перед этим была связана с полипептидной цепью, покидает рибосому и возвращается в цитоплазму, чтобы образовать новый комплекс аминоацил-тРНК (рис. 23.29, В).
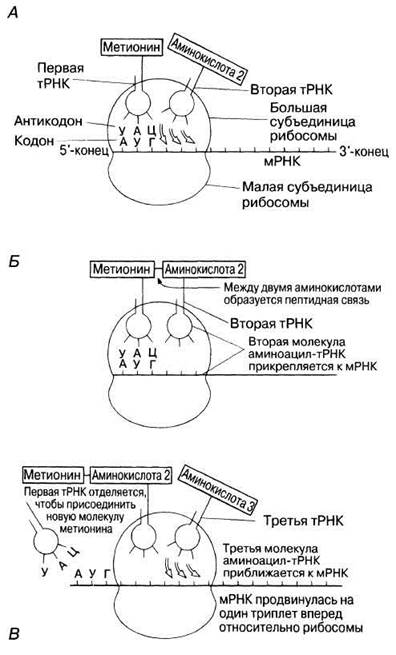
Рис. 23.29. А и Б. Стадии процесса прикрепления молекул аминоацил-тРНК своими антикодонами к кодонам на мРНК и образование пептидной связи между соседними аминокислотами. В. Относительные перемещения мРНК и рибосомы, в результате которых открывается новый триплет (рамка) для присоединения молекулы аминоацил-тРНК. Первая молекула тРНК отделяется от рибосомы и возвращается в цитоплазму, где ферменты реактивируют ее, делая возможным образование нового комплекса аминоацил-тРНК.
Такое последовательное «считывание» и «трансляция» рибосомой заключенного в мРНК кода продолжается до тех пор, пока процесс не доходит до одного из стоп-кодонов (терминирующих кодонов). Такими кодонами служат триплеты УАА, УАГ и УГА. На этом этапе полипептидная цепь, первичная структура которой была детерминирована ДНК, покидает рибосому, и трансляция завершается. Главные этапы процесса трансляции:
1) присоединение мРНК к рибосоме;
2) активация аминокислоты и ее присоединение к тРНК;
3) инициация (начало синтеза) полипептидной цепи;
4) элонгация (удлинение) цепи;
5) терминация (окончание синтеза) цепи;
6) дальнейшее использование мРНК или ее разрушение.
Процесс трансляции схематически представлен на рис. 23.30.
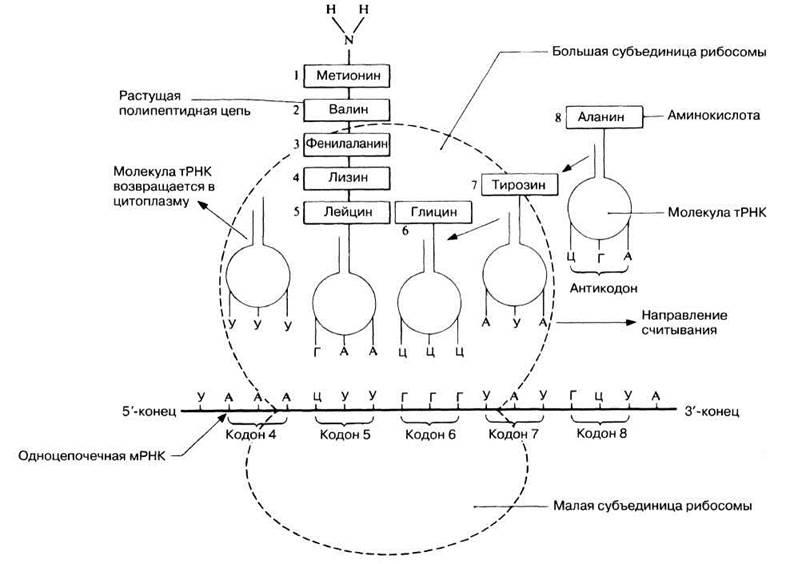
Рис. 23.30. Схематическое изображение процесса трансляции. Антикодон каждой отдельной молекулы аминоацил-тРНК спаривается с комплементарным ему кодоном мРНК на рибосоме. В изображенном здесь случае за этим последует образование пептидной связи между лейцином и глицином и таким образом к растущей полипептидной цепи добавится еще одна аминокислота.
После того, как полипептидные цепи отделились от рибосомы, они могут тотчас же приобретать свойственную им вторичную, третичную или четвертичную структуру (разд. 3.5.3). Если рибосома прикреплена к ЭР, то белок поступает в ЭР, откуда затем траспортируется к месту назначения. Первая часть растущей полипептидной цепи состоит при этом из «сигнальной последовательности» аминокислот, которая соответствует определенному рецептору в мембране ЭР, что обеспечивает присоединение рибосомы к ЭР. Растущая белковая молекула через рецептор поступает в ЭР (рис. 23.31). После того как белок оказывается внутри ЭР, сигнальная последовательность отделяется от него, и белковая молекула свертывается, приобретая свою окончательную форму.
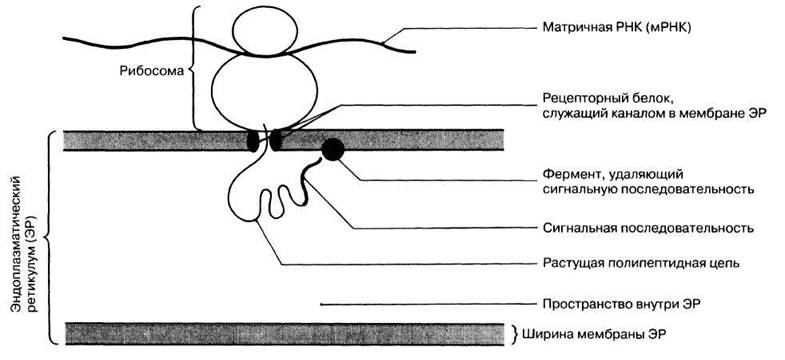
Рис. 23.31. Поступление новообразованного белка в эндоплазматический ретикулум.
Доказательства того, что включение аминокислоты в полипептидную цепь определяется комплементарным спариванием оснований между кодоном мРНК и антикодоном тРНК, а не самой аминокислотой, были получены в следующем эксперименте. Молекула тРНК-цистеин обычно спаривается при помощи своего антикодона АЦА с кодоном УГУ мРНК. При воздействии на этот комплекс катализатора — никеля Ренея — цистеин превращается в аминокислоту аланин. Когда такой новый комплекс тРНК-аланин (несущий антикодон тРНК-цистеина) помещали в бесклеточную систему, содержавшую поли(УГУ)-мРНК, синтезированная полипиптидная цепь состояла только из аланина. Этот эксперимент подтвердил важную роль механизма образования комплекса мРНК-кодон — тРНК-антикодон в трансляции генетического кода.
Вся последовательность событий, из которых складывается процесс белкового синтеза, схематически представлена на рис. 23.32.
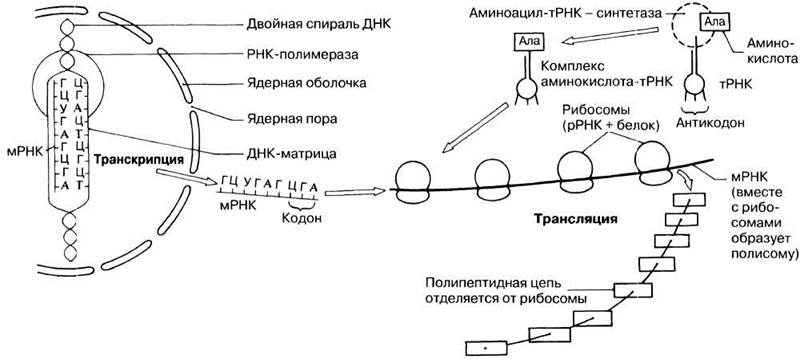
Рис. 23.32. Упрощенная обобщающая схема главных структур и процессов, участвующих в белковом синтезе.
23.8.8. Некодирующая ДНК
ДНК человека содержит примерно 3000 млн. пар оснований и, согласно оценкам, 100 000 генов, хотя в настоящее время эти оценки могут быть лишь весьма приблизительными. Проблема состоит в том, что большая часть ДНК (примерно 95%), по-видимому, лишена каких-либо явных функций, потому что она не участвует в кодировании. Иными словами, она не кодирует ни синтез белков, ни синтез РНК. Иногда ее называют «бросовой ДНК», хотя предполагать отсутствие функции лишь на том основании, что эти функции неизвестны, неразумно. Возможно, что часть этой ДНК представляет собой бывшие гены, которые в настоящее время не служат никаким полезным целям. Другая часть может выполнять структурные функции, участвуя, например, в упаковке хромосом. Примерно 30-40% этой ДНК состоят из многократно повторяющихся коротких последовательностей оснований. К этой части ДНК относится сателлитная ДНК, роль которой в «генетической дактилоскопии» описана в разд. 25.7.12. Некоторые участки ДНК, называемые нитронами, рассматриваются ниже.
Интроны и экзоны
В 1977 г. биологи были удивлены, обнаружив в ДНК эукариотических организмов гены, которые длиннее соответствующей мРНК. Казалось бы мРНК должна иметь точно такую же длину, как ген, так как она представляет собой его копию. Выяснилось, что сразу же после завершения синтеза мРНК некоторые участки ее молекулы еще до ее использования в трансляции вырезаются. Участки гена, кодирующие эти неиспользуемые участки РНК, назвали нитронами. Остальные участки гена, кодирующие соответствующий белок, назвали экзонами (рис. 23.33). Размеры и расположение интронов очень разнообразны и характерны для данного гена. У прокариот интронов нет.
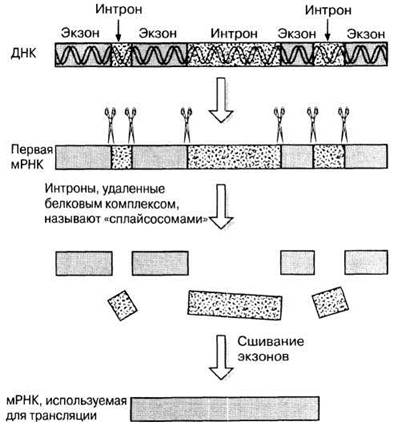
Рис. 23.33. Экзоны, интроны и вырезание интронов.
Одна из возможных функций интронов была обнаружена, когда оказалось, что из одной и той же мРНК в разных клетках могут быть удалены разные интроны. Таким образом, ген может иметь альтернативные интроны и кодировать различные, хотя и сходные белки. Это увеличивает его потенциальное использование. Примером служит ген кальцитонина. Этот ген может продуцировать две разные формы мРНК в зависимости от того, какие интроны были удалены. Одна из них образуется в щитовидной железе и кодирует синтез белка кальцитонина, состоящего из 32 аминокислот. Кальцитонин — это гормон, понижающий уровень кальция в крови. Другая форма мРНК синтезируется в гипоталамусе и кодирует белок, состоящий из 37 аминокислот; этот белок, названный пептидом, связанным с геном кальцитонина, сходен с кальцитонином и обладает сильным сосудорасширяющим действием. Он выделяется также из нервных окончаний в некоторых частях периферической нервной системы.