БИОЛОГИЯ Том 3 - руководство по общей биологии - 2004
24. ИЗМЕНЧИВОСТЬ И ГЕНЕТИКА
24.3. Сцепление
Все ситуации и примеры, обсуждавшиеся в этой главе до сих пор, относились к наследованию генов, находящихся в разных хромосомах. Согласно данным цитологических исследований, у человека все соматические клетки содержат по 46 хромосом. Поскольку человек обладает тысячами различных признаков, таких, например, как группа крови, цвет глаз, способность секретировать инсулин и т. п., в каждой хромосоме должно находиться большое число генов.
Гены, расположенные в одной и той же хромосоме, называют сцепленными. Все гены одной хромосомы образуют группу сцепления; они обычно попадают в одну гамету и наследуются вместе. Таким образом, гены, принадлежащие к одной группе сцепления, обычно не подчиняются менделевскому закону независимого распределения. Поэтому при дигибридном скрещивании эти гены не дают ожидаемого соотношения 9:3:3:1. В таких случаях получаются самые разнообразные соотношения, которые теперь, когда нам известны закономерности, открытые Менделем, можно довольно легко объяснить. (Здесь следует еще раз подчеркнуть, что Менделю посчастливилось выбрать для изучения наследования пары признаков гены, локализованные в разных хромосомах.) У дрозофилы гены, контролирующие окраску тела и длину крыла, имеют следующие аллеломорфы (фенотипические признаки, определяемые разными аллелями): серое тело — черное тело, длинные крылья — зачаточные (короткие) крылья. Серое тело и длинные крылья доминируют. При скрещивании гомозиготной мухи с серым телом и длинными крыльями и гомозиготной мухи с черным телом и зачаточными крыльями ожидаемое соотношение фенотипов в F2 должно составить 9:3:3:1. Это указывало бы на обычное менделевское наследование при дигибридном скрещивании, обусловленное случайным распределением генов окраски тела и длины крыла, находящихся в разных негомологичных хромосомах. Однако вместо этого в F2 были получены в основном родительские фенотипы в отношении примерно 3:1. Это можно объяснить, предположив, что гены окраски тела и длины крыла локализованы в одной и той же хромосоме, т. е. сцеплены (рис. 24.8).
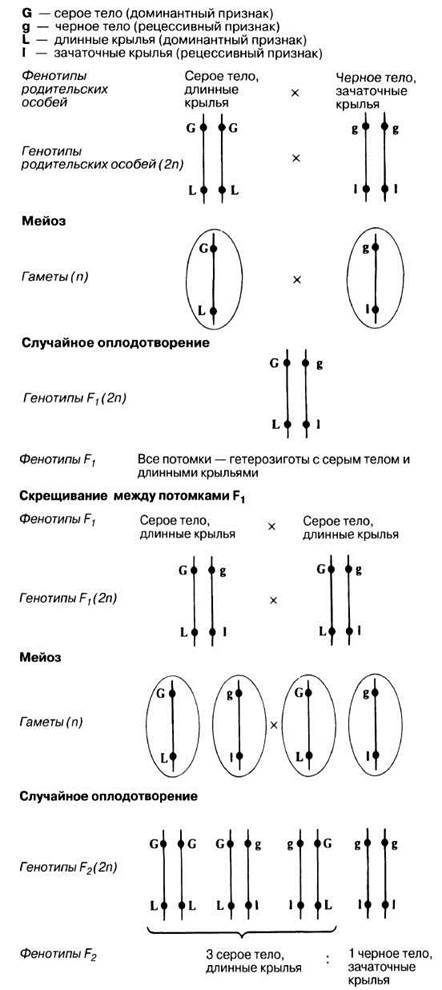
Рис. 24.8. Генетическое объяснение отношения 3:1 между фенотипами F2 как результата сцепления.
Практически, однако, соотношение 3 :1 никогда не получается, а в F2 неизбежно появляются все четыре фенотипа. Это объясняется тем, что полное сцепление встречается крайне редко. В большинстве экспериментов по скрещиванию при наличии сцепления помимо мух с родительскими фенотипами обнаруживаются особи с новыми сочетаниями признаков. Эти новые фенотипы называют рекомбинантными. Все это позволяет дать следующее определение сцепления: два ши более генов называют сцепленными, если потомки с новыми генными комбинациями (рекомбинанты) встречаются реже, чем родительские фенотипы.
События, приведшие американского исследователя Томаса Моргана к открытию сцепления, можно проиллюстрировать одним из его экспериментов на дрозофиле, в котором он предсказал результаты анализирующего скрещивания между серой длиннокрылой гетерозиготой (из поколения F1 от скрещивания, представленного на рис. 24.8) и рецессивной гомозиготой с черным телом и зачаточными крыльями. Было предложено два возможных результата.
1. Если две пары аллелей, определяющие серую или черную окраску тела и длинные или зачаточные крылья, лежат в разных хромосомах (т. е. не сцеплены), то они должны распределяться независимо при следующем соотношении фенотипов:
1 серое тело, длинные крылья : 1 серое тело, зачаточные крылья;
1 черное тело, длинные крылья : 1 черное тело, зачаточные крылья.
2. Если аллели, определяющие окраску тела и длину крыльев, принадлежат одной и той же паре хромосом (т. е. сцеплены), то соотношение фенотипов будет иным:
1 серое тело, длинные крылья : 1 черное тело, зачаточные крылья.
Объяснение этих предсказаний представлено на рис. 24.9.
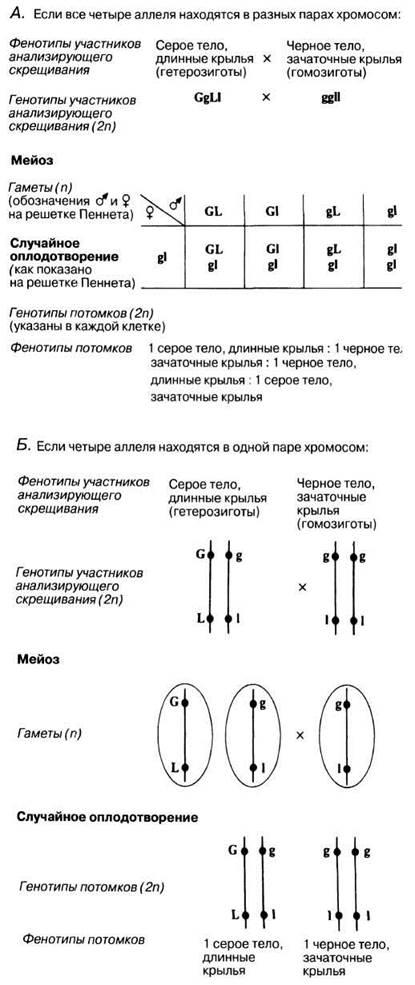
Рис. 24.9. Генетическое объяснение предсказаний Моргана.
Морган провел это анализирующее скрещивание несколько раз и ни разу не получил ни один из предсказанных результатов. Всякий раз он получал следующие результаты:
41,5% — серое тело, длинные крылья
41,5% — черное тело, зачаточные крылья
8,5% — серое тело, зачаточные крылья
8,5% — черное тело, длинные крылья
На основе этих результатов Морган постулировал, что
1) изучаемые гены локализованы в хромосомах;
2) оба гена находятся в одной хромосоме, т. е. сцеплены;
3) аллели каждого гена находятся в гомологичных хромосомах;
4) во время мейоза между гомологичными хромосомами происходит обмен аллелями.
Появление рекомбинантных сочетаний аллелей у 17% потомков было объяснено на основе пункта 4. Это явление получило название кроссинговера (перекреста).
24.7. Гомозиготное растение с пурпурными цветками и коротким стеблем скрестили с гомозиготным растением с красными цветками и длинным стеблем; у гибридов F1 были пурпурные цветки и короткий стебель. При анализирующем скрещивании растения из F1 с двойной гомозиготой по рецессивным генам были получены следующие потомки:
52 с пурпурными цветками и коротким стеблем
47 с пурпурными цветками и длинным стеблем
49 с красными цветками и коротким стеблем
45 с красными цветками и длинным стеблем
Дайте полное объяснение этих результатов.
24.3.1. Кроссинговер и частота рекомбинаций
В 1909 г. бельгийский цитолог Янсене наблюдал образование хиазм во время профазы I мейоза (разд. 23.4). Генетическое происхождение этого процесса разъяснил Морган, высказав мнение, что кроссинговер (обмен аллелями) происходит в результате разрыва и рекомбинации гомологичных хромосом во время образования хиазм. В дальнейшем сопоставление данных микроскопических исследований с данными о соотношениях рекомбинантных фенотипов подтвердило, что обмен генетическим материалом происходит практически между всеми гомозиготными хромосомами. Аллели, входящие в группу сцепления у родительских особей, разделяются и образуют новые сочетания, которые попадают в гаметы, — процесс, называемый генетической рекомбинацией. Потомков, которые получаются из таких гамет с «новыми» сочетаниями аллелей, называют рекомбинантами, или кроссоверами. Таким образом, кроссинговер представляет собой важный источник генетической изменчивости, наблюдаемой в популяциях.
Для иллюстрации принципа кроссинговера можно рассмотреть поведение пары гомологичных хромосом дрозофилы, несущих аллели серой окраски тела и длинных крыльев (оба аллеля доминантные) и черной окраски тела и зачаточных крыльев (оба аллеля рецессивные), во время образования хиазм. Скрещивание между гетерозиготным серым длиннокрылым самцом и гомозиготной черной самкой с зачаточными крыльями дало в F1 гетерозиготных потомков с серым телом и длинными крыльями (рис. 24.10).
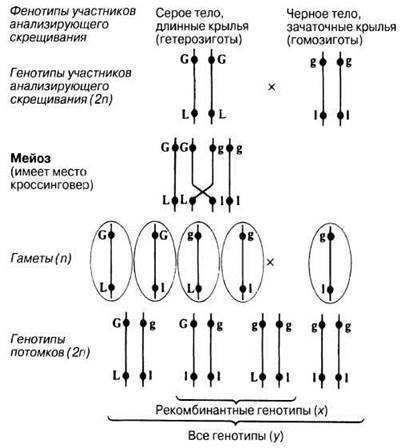
Pиc. 24.10. Генетическое объяснение кроссинговера и появления рекомбинантных генотипов. Подсчитав число особей, у которых выявляется рекомбинация (х) и общее число особей (у), можно вычислить частоту рекомбинаций по формуле:
Частота рекомбинации (%) = х/у х 100
В результате анализирующего скрещивания мух из поколения F1 с гомозиготами по двум рецессивным генам были получены следующие результаты:

Как показывают эти результаты, гены, определяющие окраску тела и длину крыльев, сцеплены. (Напомним, что если бы эти гены находились в разных хромосомах и поэтому распределялись случайным образом, то при скрещивании гетерозиготы из F1 с гомозиготой по двум рецессивным признакам соотношение фенотипов было бы 1:1:1:1.) Из приведенных величин можно вычислить частоту рекомбинации генов, определяющих окраску тела и длину крыльев.
Частота рекомбинаций вычисляется по формуле
![]()
В нашем примере частота рекомбинации равна
![]()
Это значение соответствует числу рекомбинаций, происходящих при образовании гамет. Один из учеников Моргана, А. X. Стертевант, высказал мысль, что частоты рекомбинациисвидетельствуют о линейном расположении генов вдоль хромосомы. Еще более важное предположение Стертеванта состояло в том, что частота рекомбинации отражает относительное расположение генов в хромосоме: чем дальше друг от друга находятся сцепленные гены, тем больше вероятность того, что между ними произойдет кроссинговер, т. е. тем выше частота кроссоверов (рис. 24.11).
![]()
Рис. 24.11. Три генных локуса — А, В и С — расположены в одной хромосоме. Вероятность кроссинговера и разделения для генов В и С выше, чем для генов А и В, так как частота кроссинговера зависит от расстояния между генами.
24.8. На приведенной ниже схеме изображены локусы 12 аллелей, расположенные в паре хромосом. Показаны их относительные расстояния от центромеры.
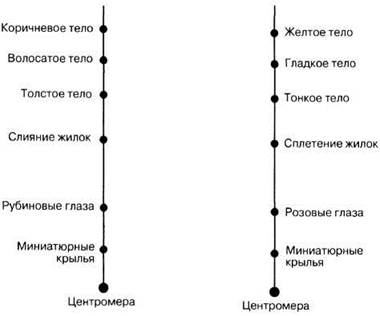
а) Как называются изображенные здесь хромосомы?
б) Между какими двумя локусами чаще всего будет происходить кроссинговер?