Альгологія - Костіков І.Ю. - 2009-2013
Розділ 16. Бурі водорості – Phaeophyta
Відділ бурі водорості нараховує біля 260 родів, представлених приблизно 2000 видів. Всі представники цього відділу є багатоклітинними, перважно - макроскопічними водоростями, і поширені майже виключно в морях. Найдавніші достовірні викопні рештки бурих водоростей мають вік біля 400 млн.років, а їх вірогідні рештки - біля 570 млн.років. За молекулярними ознаками відділ близько споріднений з Xanthophyta.
Систематичні ознаки відділу
Пігменти та запасні поживні речовини
Бурі водорості мають хлорофіли а та с, β- та ε-каротини, та дві основні групи ксантофілів: по-перше, ксантофіли жовтої пігментної групи - фукоксантин, діатоксантин, діадіноксантин, та по-друге, ксантофіли лютеїнового ряду - віолоксантин, зеаксантин, антероксантин, неоксантин. Фукоксантин та віолоксантин накопичуються у великій кількості і маскують хлорофіли, надаючи хлоропластам яскраво-жовтого або бурого забарвлення.
Основними продуктами асиміляції є полісахарид ламінарин та олія, рідше - багатоатомні спирти (наприклад, манніт). Всі продукти асміляції відкладаються поза межами хлоропластів.
Цитологічні ознаки
Клітинні покриви представлені оболонками. Оболонка поділяється на два шари - зовнішній, до складу якого входять пектнові речовини та розчинні солі альгінової кислоти (переважно альгінат натрію), та внутрішній шар, що має фібрилярну структуру. Основою внутрішнього шару є мікрофібрили целюлози, що утворюють сітчастий каркас оболонки та нерозчинний альгінат кальцію.
Альгінова кислота - це гетерополісахарид, що складається з залишків D- маннуронової та L-гулуронової кислот. Солі альгінової кислоти - альгінати - утворюють гелі. В’язкість гелю визначається складом та пропорціями катіонів, з якими зв’язується альгінова кислота.
Таким чином, клітинні оболонки завдяки великій кількості пектинових речовин та альгінатів здатні сильно ослизнюватись і перебувають у гелеподібному колоїдному стані.
У деяких бурих водоростей крім альгінатів до складу оболонки входить сульфатований гетерополісахарид фукоїдан, що складається з залишків олігосахаридів фукози, галактози, маннози, ксилози та глюкуронової кислоти.
Ядерний апарат. Ядро має типову евкаріотну будову. Як і у переважної більшості водоростей-страменопілів, зовнішня мембрана ядерної оболонки переходить у хлоропластну ендоплазматичну сітку (рис.16.1). У ядрі спостерігається велике, добре помітне ядерце. Біля ядерної оболонки розташовуються кілька комплексів Гольджі та дві центриолі. Останні при каріокінезі стають центрами організації мікротрубочок веретена поділу. На початку поділу ядра вони реплікуються та мігрують до протилежних полюсів ядра. Цікавою особливістю веретена є утворення навколо центриолей астральних променів з мікротрубочок, подібно до того, як це спостерігається у багатоклітинних тварин.
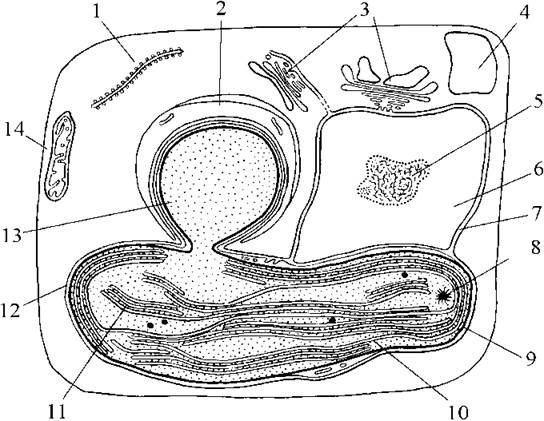
Рис. 16.1. Схема будови клітини бурої водорості (на прикладі ектокарпальних): 1 - ендоплзматична сітка; 2 - платівка ламінарину навколо піреноїду; 3 - комплекс Гольджі; 4 - вакуоля; 5 - ядерце; 6 - ядро; 7 - ядерна оболонка; 8 - хлоропластна ДНК; 9 - двомембранна оболонка хлоропласту; 10 - оперезуючий тилакоїд; 11 - тритилакоїдна ламела; 12 - хлоропластна ендоплазматична сітка; 13 - виступаючий піреноїд; 14 - мітохондріальний профіль (за Bouck, 1965).
У профазі ядерце зникає, ядерна оболонка біля центриолей руйнується, утворюючи дві великі пори. Таким чином, мітоз у Phaeophyta напівзакритий та центричний. Метафаза та анафаза проходять за типовою для евкаріот схемою. У телофазі дочірні ядра розходяться, веретено швидко зникає, після чого ядра знов дещо зближуються. Водночас з цим плазмалема утворює борозну поділу, розділяє ядра та протопласт, і починає секрецію речовин нових клітинних перегородок, причому у поперечній перегородці лишаються пори.
При утворенні зооспор на заключних стадіях цитокінезу у деяких видів (наприклад, у Chorda tomentosa) центриолі мігрують до поверхні дочірніх клітин і перетворюються на базальні тіла джгутиків.
Фотосинтетичний апарат. Хлоропласти бурих водоростей дрібні, численні. Кожний хлоропласт вкритий чотирма мембранами, з яких дві зовнішні утворюють хлоропластну ендоплазматичну сітку, що переходить у ядерну оболонку, а дві внутрішні є власне оболонкою хлоропласту. Між зовнішніми та внутрішніми мембранами є перипластидний простір.
Тилакоїди розміщуються переважно по три, на периферії знаходиться оперезуюча ламела. У багатьох бурих водоростей у хлоропласті є піреноїд. Подібно до золотистих, евстигматофітових та жовтозелених водоростей, він не пронизаний тилакодами. У бурих водоростей з порядку Ectocarpales піреноїд виступаючого типу. Звичайно навколо нього, проте за межами хлоропласту, відкладається ламінарин. Платівки ламінарину оточені власною цитоплазматичною мембраною. У більшості інших бурих водоростей піреноїд рудиментарний, і виявляється тільки методами електронної мікроскопії, причому інколи лише у репродуктивних клітин (зооспор, гамет) або зигот.
Хлоропластна ДНК організована у кільцеподібно замкнений нуклеоїд, що складається із 110-170 тис. пар нуклеотидів і розташовується під оперезуючою ламелою.
У монадних клітин в стромі хлоропласту розташовується також стигма. Вона являє собою шар пігментних глобул, і її положення зкоординовано із положенням парабазального тіла, розташованого при основі гладенького джгутика. Подібно до золотистих та жовтозелених водоростей стигма виконує функцію ширми, що прикриває фоторецептор, розташований у парабазальному тілі.
Мітохондріальний апарат представлений розгалуженим мітохондріоном, внутрішня мембрана якого утворює трубчасті кристи. Мітохондріальна ДНК має кільцеподібну організацію і складається приблизно з 50 тис. пар нуклеотидів.
Джгутикові стадії представлені зооспорами та гаметами, що мають два джгутики нерівної довжини. На відміну від інших водоростей-страменопілв, джгутики розташовуються не апікально або субапікально, а на черевному боці клітини (рис. 16.2). Довший джгутик пірчастий і вкритий тричленними матигонемами. Мастигонеми утворюються у цистернах ендоплазматичної сітки та у пухирцях, що відшнуровуються від зовнішньої мембрани ядерної оболонки. Вони складаються із розширеної підошви, мікротубулярної проміжної частини та фібрилярних термінальних волосків. Коротший джгутик гладенький. У диктиотальних водорості гладенький джгутик редукований і від нього зберігається лише базальне тіло. У сперматозоїдів бурих водоростей на передньому кінці клітини є складчаста структура - хобіток, за допомогою якого відбувається дихання та пошук яйцеклітини.
Перехідна зона джгутиків у бурих водоростей позбавлена спіралеподібної структури, яка є характерною для переважної більшості водоростей-страменопілів. Базальні тіла джгутиків з'єднуються між собою поперечно-смугастим сполучним волокном.
Коренева система джгутиків представлена чотирма коренями. Перший корінь складається з семи мікротрубочок, і направлений до переднього кінця клітини. Другий корінь проходить від заднього кінця клітини до переднього, з'єднуючись у центральній частині з базальними тілами двома сполучними волокнами. Цей корінь називають двонаправленим. До його складу входять п’ять мікротрубочок та мікрофібрилярне волокно, що асоційоване з направленою до заднього кінця клітини мікротубулярною частиною. Третій та четвертий корені утворені однією-двома мікротрубочками і проходять поблизу ядерної оболонки; один корінь поблизу ядра розвертається вперед, другий - назад.
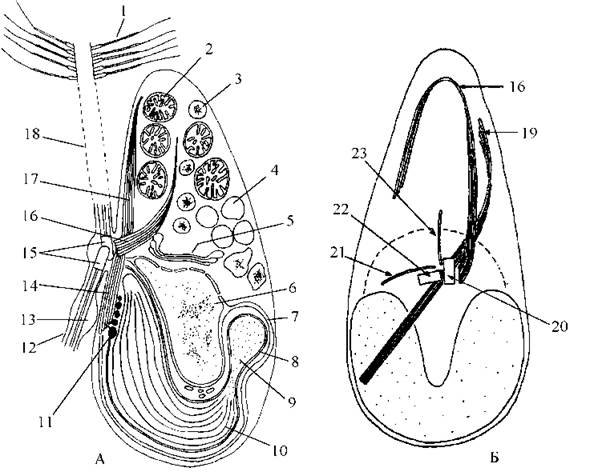
Рис. 16.2. Будова зооспори бурих водоростей: А - загальний план будови (латеральний вид); Б - коренева система джгутиків (вентральний вид). 1 - тричленна мастигонема; 2 - мітохондріальний профіль; 3 - вакуоля зі хризоламінарином; 4 - вакуоля з клітинним соком; 5 - комплекс Гольджі; 6 - ядро; 7 - хлоропластна ендоплазматична сітка; 8 - двомембранна хлоропластна оболонка; 9 - піреноїд; 10 - тритилакоїдна ламела; 11 - глобула стигми; 12 - короткий джгутик; 13 - парабазальне тіло; 14 - нижня частина двонаправленого мікротубулярного кореня; 15 - базальні тіла; 16 - передній мікротубулярний корінь; 17 - верхня частина двонаправленого кореня; 18 - довгий джгутик; 19 - двонаправленй корінь; 20, 21 - базальні тіла довгого та короткого джгутиків, відповідно; 22, 23 - третій та четвертий мікротубулярні корені (за Manton, 1957; O'Kelly, Floyd, 1984).
Типи морфологічних структур та будова талому
В межах відділу представлено нитчастий, гетеротрихальний та справжній тканинний типи структур тіла. Всі бурі водорості, за винятком деяких видів роду Sargassum, ведуть прикріплений спосіб життя. Органами прикріплення є ризоїди або дископодібні підошви.
Таломи нитчастих представників перевжно утворені системою однорядних розгалужених ниток (наприклад, рід Ectocarpus). Гетеротрихальні водорості мають переважно вигляд досить товстих шнурів. Такі шнури мають або одноосьову, або багатоосьову будову (рис. 16.3).
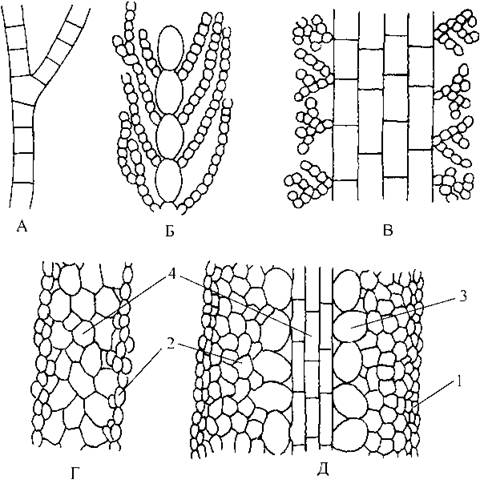
Рис. 16.3. Схема будови таломів бурих водоростей. А - одноосьовий однорядний талом; Б - одноосьовий багаторядний; В - багатоосьовий; Г - "простий" тканинний; Д - "складний" тканинний. 1 - меристодерма, 2 - кора, 3 - проміжна тканина, 4 — серцевина.
Різноманітна форма таломів характерна для водоростей з тканинною будовою. У найпростішому випадку такий талом складається лише з двох типів тканин - кори та серцевини. Кора утворена дрібними забарвленими клітинами, що виконують функцію фотосинтезу та з яких утворюються органи розмноження. Серцевина складається з великих, переважно безбарвних клітин, що виконують провідну та запасаючу функції.
У складніших випадках талом утворений більшою кількістю тканин: меристодермою, корою, проміжною тканиною та серцевиною. Меристодерма - це поверхнева тканина, що виконує захисну та репродуктивну функції. Клітини меристодерми звичайно дрібні, і здатні до активного поділу. Кора утворена більшими за розміром клітинами з добре розвиненими хлоропластами. Основною функцією кори є здійснення процесу фотосинтезу. У клітинах проміжної тканини накопичуються продукти асиміляції. Самі клітини великі, звичайно з редукованими хлоропластами. Серцевина утворена ситовидними трубками та трубчастими нитками.
Ситовидні трубки витягнуті у довжину, їх поперечні стінки мають пори з досить товстими плазмадесмами. У цитоплазмі міститься розвинений комплекс Гольджі, мітохондрії, ендоплазматична сітка. Цікаво, що в одних представників ситовидні трубки мають велике лопатеве ядро з добре помітним ядерцем (Laminaria), у інших ядро відсутнє (Macrocystis). Ситовидні трубки виконують провідну функцію: спеціальними дослідженнями встановлено, що по них транспортується маніт та амінокислоти, причому швидкість руху становить 0.10.6 м за годину.
Нерідко у серцевині та проміжній тканині знаходяться клітини-фізоїди, заповнені пухирцями з танінами (флюроглюцином та поліфенолами) та слизові канали з секреторними клітинами.
Ріст таломів бурих водоростей відбувається кількома шляхами: за рахунок поділу клітин середньої частини талому (інтеркалярний ріст), групи верхівкових клітин (апікальної меристеми), лише однієї великої верхівкової клітини або трихоталічно - шляхом поділу групи клітин, що розташовуються субапікально - між верхівковими волосками та основною частиною "простого" тканинного талому. Тип наростання талому є однією з систематичних ознак на рівні порядків.
Розмноження та життєві цикли
Розмножуються бурі водорості нестатевим та статевим шляхами. Нестатеве розмноження може відбуватися фрагментацією таломів, вивідковими бруньками, зооспорами та тетраспорами.
Фрагментація таломів відбувається звичайно під час сильних штормів. Проте у переважній більшості випадків фрагменти не здатні прикріплюватись до субстрату, а у вільноплаваючому стані не розвиваються у нормальну нову особину. Такі фрагменти, звичайно, під час штормів викидаються на берег та гинуть. Проте клітини фрагментів таломів у частини видів здатні протягом короткого часу перетворюватися на зооспорангії. Таким чином, опосередковано фрагментація приймає участь у здійсненні розселювальної функції.
У видів роду Sphacellaria вегетативне розмноження здійснюється за допомогою спеціалізованих вивідкових бруньок, що здатні прикріплюватися до субстрату та розвиватися у нормальну репродуктивно дозрілу особину (рис. 16.4).
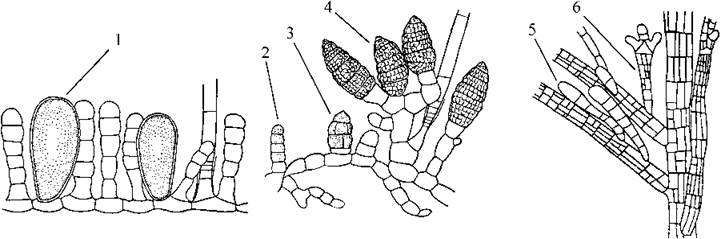
Рис. 16.4. Органи нестатевого розмноження бурих водоростей. 1 - одногніздний спорангій; 2, 3 - послідовні стадії розвитку багатогніздного спорангію; 4 - зрілий багатогніздний спорангій; 5, 6 - зачаткова (5) та зріла (6) вивідкова бруньки у Sphacellaria (за Зинова, 1967; Петров, 1977).
Нестатеве розмноження за допомогою зооспор властиве переважній більшості бурих водоростей, за винятком диктиотальних та фукальних. Зооспорангії бурих водоростей можуть бути одногніздні або багатогніздні. Одногніздний спорангій являє собою збільшену за розмірами клітину, вміст якої після серії поділів ядра розпадається на велику кількість зооспор. Перший поділ ядра звичайно редукційний, і тому зооспори, що утворюються в одногніздних спорангіях мають гаплоїдний набір хромосом. Багатогніздні спорангії утворюються з великої клітини, в якій перед формуванням спор протоласт розділяється тонкими клітинними перегородками на багато одноядерних камер (гнізд). У кожному гнізду утворюється одна зооспора (див. рис.16.4).
У диктиотальних водоростей зооспори відсутні, замість них утворюються нерухливі тетраспори, а у нечисленної групи тилоптеридієвих - моноспори.
Статевий процес відбувається у вигляді ізо- або оогамії, дуже зрідка зустрічається гетерогамія. Гаметангії бурих водоростей багатогніздні (за винятком представників Fucales). У нитчастих форм гаметангії утворюються на особливих бічних гілках, у гетеротрихальних - з клітин асиміляторних ниток, у тканинних - з кори або меристодерми.
У видів з оогамним типом статевого процесу перед копуляцією яйцеклітину оточує велика кількість сперматозоїдів. Переднім джгутиком вони прикріплюються до мембрани яйцеклітини. Далі мембрана джгутику одного з них зливається з плазмалемою яйцеклітини, у протопласт проникає аксонема, а за нею і ядро сперматозоїду. На цій стадії інші сперматозоїди відкріпляються та залишають яйцеклітину. Після злиття ядер утворюється зигота, яка без періоду спокою проростає у нову особину.
У бурих водоростей життєвий цикл гаплодиплофазний з чергуванням поколінь (ізоморфним або гетероморфним) або диплофазний без зміни поколінь.
При гаплодиплофазному циклі на диплоїдному спорофіті в одногніздних спорангіях відбувається мейоз, і утворюються гаплоїдні зооспори. Вони проростають у гаплоїдні гаметофіти. Після копуляції зигота без періоду спокою розвивається у новий спорофіт. Якщо чергування поколінь гетероморфне, то гаметофіти, як правило, мікроскопічні, а спорофіти макроскопічні.
Зміна поколінь може бути усталеною або неусталеною. При усталеній зміні (наприклад, у Laminaria) спорофіт завжди дає гаплоїдні спори, що проростають тільки у гаметофіт. Останній в свою чергу продукує гамети, які не здатні до партеногенетичного розвитку.
При неусталеній зміні поколінь спорфіт може утворювати як одно-, так і багатогніздні спорангії, відповідно з гаплоїдними або диплоїдним зооспорами. Гаплоїдні зооспори проростають у гаплоїдний гаметофіт, диплоїдні - диплоїдний спорофіт. На гаметофіті утворюються гамети, які або копулюють (в цьому випадку зигота проростає у спорофіт), або проростають партеногенетично у новий гаметофіт. Таким чином спорофіт та гаметофіт не закономірно змінюють один одного, і чергування поколінь є неусталеним.
Представники з диплофазним життєвим циклом не мають зооспор, мейоз відбувається при утворенні гамет, а статевий процес переважно оогамний. Оогонії та антеридії утворюються на спеціальних плодущих гілочках (рецептакулах) в напівзамкнутих порожнинах - скафідіях. Далі сперматозоїди, що утворились в антеридіях чоловічих скафідіїв, виходять назовні. В жіночих скафідіях оболонки оогоніїв розриваються, яйцеклітини потрапляють у порожнину скафідія, і виділяють атрактанти, які приваблюють сперматозоїди. Після запліднення зигота без періоду спокою проростає, не виходячи за межи скафідію.
Система відділу
Сьогодні існує дві групи систем Phaeophyta - класичні системи, що ґрунтуються на ознаках, пов’язаних з особливостями розмноження, та синтетична система. Остання розроблена за молекулярними даними на основі аналізу нуклеотидних послідовностей цілого ряду генів (зокрема, тих що кодують малу та велику субодиниці рибосомальної РНК, RuBisCo, фактори подовження тубуліну, актин), та узгоджена з комплексом фенотипічних ознак. Важливим є й те, що синтетична система відповідає вимогам Міжнародного Кодексу ботанічної номенклатури.
Класичні системи
За класичними системами відділ найчастіше поділяють на два класи - Phaeophyceae (=Phaeozoosporosphyceae) та Cyclosporophyceae. В основі поділу покладено тип життєвого циклу та здатність до розмноження за допомогою спор.
Phaeophyceae (=Phaeozoosporosphyceae) об’єднує водорості, що мають гаплодиплофазний життєвий цикл та здатні до нестатевого розмноження за допомогою зооспор або тетраспор. Згідно до типу чергування поколінь (ізо- або гетероморфне, усталене або ні), типу статевого процесу, плану будови талому та характеру його наростання різні автори виділяють у межах класу від 11 до 15 порядків, серед яких провідними вважаються Ectocarpales, Dictyosiphonales, Cutleriales, Sphacellariales, Dictyotales та Laminariales. Характерними представниками класу є Ectocarpus,Dictyosiphon, Cutleria, Sphacellaria, Dictyota, Laminaria, Macrocystis.
Cyclosporophyceae включає водорості з диплофазним життєвим циклом, що не здатні утворювати спори. Клас поділяють на три порядки - Ascoseirales, Durvilleales, Fucales. Характерними представниками є Ascoseira, Durvillea, Fucus, Ascophyllum, Cystoseira, Sargassum.
Синтетична система
Система була створена внаслідок сумісних досліджень систематиків та молекулярних біологів і набула цілісного вигляду лише в останні роки ХХ ст. Згідно з цією системою, відділ Phaeophytaвключає лише один клас - Phaeophyceae та вісім порядків, з яких нижче розглядаються сім1.
Молекулярно-філогенетичні клади, що відповідають рівню порядків, корелюють з такими фенотипічними ознаками, як наявність та будова піреноїду, особливості розмноження, наявність зооспор, тип статевого процесу, життєвий цикл. Основні фенотипічні ознаки порядків подано у таблиці 16.1.
Ectocarpales - об’єднує водорості, які, на відміну від інших бурих водоростей, мають добре розвинений піреноїд виступаючого типу. Таломи ектокарпальних можуть мати нитчасту, гетеротрихальну або "просту" тканинну будову. Життєвий цикл диплогаплофазний, з неусталеним чергуванням поколінь, причому у випадку гетероморфної зміни спорофіти макро-, а гаметофіти мікроскопічні. За молекулярними даними, Ectocarpales є найпримітивнішим порядком бурих водоростей.
Таблиця 16.1. Принципи поділу бурих водоростей на порядки за фенотипічними ознаками
Ознака → Порядок ↓ |
Піреноїд |
Хлоропласт |
Життєвий цикл |
Статевий процес |
Нестатеве розмноження |
Таломи |
Ріст талому |
Ectocarpales |
В |
Пар. |
n-2n |
І, Г |
зооспори |
Н, Г, ПТ |
ІН, АМ |
Scytothamnales |
В |
Цент. |
n-2n |
? |
зооспори |
Г |
ІН, АМ |
Cutleriales |
Р |
Пар. |
n-2n |
Г |
зооспори |
ПТ |
ТТ |
Dictyothales |
Р |
Пар. |
n-2n |
О |
тетраспори |
ПТ |
АК |
Laminariales |
Р |
Пар. |
n-2n |
О |
зооспори |
СТ |
ІН |
Sphacellariales |
Р |
Пар. |
n-2n |
І, Г, О |
зооспори |
Г |
АК |
Fucales |
Р, Н |
Пар. |
2n |
Г, О |
відсутне |
СТ |
ІН, АК, АМ |
Скорочення: піреноїд: В - виступаючий розвинений, Р - рудиментарний, Н - відсутній; хлоропласт: Пар. - парієтальні численні дископодібні, Цент. - центральний поодинокий лопатевий; життєвий цикл: n-2n - гаплодиплофазний, 2n - диплофазний; статевий процес: І - ізогамія, Г - гетерогамія, О - оогамія, ? - не досліджений; таломи: Н - нитчасті, Г - гетеротрихальні, ПТ - "прості" тканинні, СТ - "складні" тканинні; ріст талому: І - інтеркалярний, АМ - апікальна меристема, ТТ - трихоталічний, АК - апікальною клітиною
Прикладом водорості з найпростішим планом будови є Ectocarpus (рис. 16.5). Його спорофіти і гаметофіти морфологічно не відрізняються між собою і мають вигляд кущиків, утворених розгалуженими однорядними нитками, що прикріплюються до підводних субстратів ризоїдами. Висхідні нитки часто закінчуються багатоклітинними безбарвними волосками. Будь-яка вегетативна клітина висхідної нитки здатня до поділу, і тому наростання висхідної частини талому відбувається інтеркалярно. Сланкі нитки, навпаки, наростають внаслідок поділу верхівкових клітин.
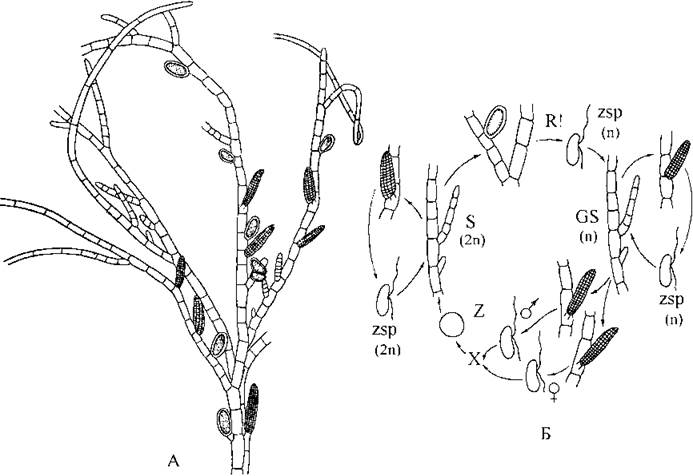
Рис. 16.5. Ectocarpus: А - спорофіт із одно- та багатогніздними зооспорангіями; Б - життєвий цикл (А - Зенова, 1967; Б - схематизовано за Mоller, 1972).
Вегетативні клітини містять пристінні стрічкоподібні хлоропласти з типовими для порядку виступаючими піреноїдами (див. рис. 16.1).
Спорофіти диплоїдні, і утворюють зооспорангії двох типів - одно- та багатогніздні, які розвиваються на верхівках коротких бічних гілочок. Зооспори, які утворилися у багатогніздних спорангіях, завжди диплоїдні і дають початок новим спорофітам. Зооспори, що зріють в одногніздних спорангіях, можуть бути як диплоїдними, так і гаплоїдними. Гаплоїдні спори надалі розвиваються у гаплоїдні гаметофіти.
Гаметофіти двостатеві та однодомні, на них розвиваються багатогніздні гаметангії, в яких утворюються ізогамети обох статевих знаків. Гамети морфологічно майже не відрізняються між собою, проте гамети, що за поведінкою вважаються жіночими ("+"-гамети) після виходу з гаметангію виділяють феромон ектокарпен. Чоловічі ("-"-гамети) рухаються за зростаючим градієнтом концентрації ектокарпену, причому хеморецепторні функції виконує субмікроскопічний складчастий апікальний хобіток. Після копуляції утворюється диплоїдна зигота, яка без періоду спокою починає проростати у новий спорофіт.
Види роду Ectocarpus зустрічаються у літоральній зоні морів всіх широт, в тому числі - у Чорному та Азовському.
Представником з гетероморфною зміною поколінь є Dictyosiphon. Спорофіти цієї водорості мають кущиків, утворених довгими (до 20 см завд.) розгалуженими трубками, стінки яких складаються з двох-трьох поверхневих шарів дрібних корових клітин та внутрішнього шару великих клітин серцевини. Ріст талому верхівковий, здійснюється внаслідок поділу дрібних клітин апікальної меристеми (рис. 16.6: А). З клітин серцевини утворюються одногніздні зооспорангії. Зміна поколінь неусталена, гетероморфна. Гаметофіт водорості мікроскопічний, причому гамети можуть партеногентично проростати у нові гаплоїдні гаметофіти. Види роду зустрічаються у морьких бентосних угрупованнях холодного та помірного поясів, звичайно оселяючись на камінні та черепашках молюсків.
Scytothamnales об'єднує водорості, які відрізняються від усіх інших наявністю центрального лопатевого зірчастого хлоропласту з центральним піреноїдом. Цей порядок встановлений лише наприкінці 90-х років ХХ ст., причому спочатку на основі молекулярних даних. Всі сцитотамнальні водорості мешкають у морях південної півкулі. Характерним представником є Stereocladon (рис. 16.6: Б).
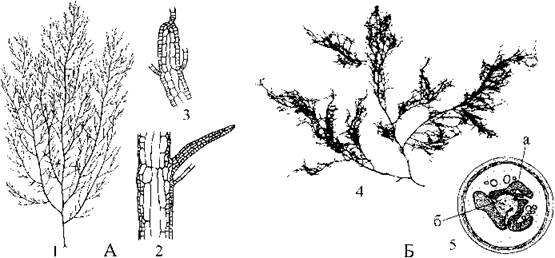
Рис. 16.6. Ектокарпальні та сцитотамнальні водорості. А - Dictyosiphon: 1 - зовнішній вигляд спорофіту; 2 - анатомічна будова талому; 3 - верхівка гілки з дрібними клітинами апікальної меристеми. Б - Stereocladon: 4 - зовнішній вигляд талому; 5 - поперечний зріз через клітину (а - центральний лопатевий хлоропласт, б - піреноїд) (А - за Sauvageau, 1927; Newton, 1931; Б - за Peters, Clayton, 1998).
Cutleriales включає морські водорості, у яких, на відміну від інших бурих водоростей, гаметофіти макроскопічні, а спорофіти переважно мікроскопічні. Таломи на верхівці закінчуються пучками волосків. При основі цих волосків розташовуються шар меристемних клітин. Останні здатні до поділу, і відчленовують від себе сегменти, орієнтовані або назовні, або всередину талому. Зовнішні сегменти обумовлюють ріст волосків у довжину, внутрішні диференціюються у тканини основної частини талому. Такий тип наростання талому називають трихоталічним. Представникам порядку властива "проста" тканинна будова, причому спорофіт і гаметофіт за формою завжди пластинчасті. Статевий процес у кутлеріальних водоростей гетерогамний.
У типового представника - Cutleria - гаметофіт має вигляд великої віялоподібної прямостоячої платівки, яка при основі переходить у коротеньке стебельце з підошвою (рис. 16.7). Спорофіт утворює дрібну мікроскопічну платівку, розпростерту по субстрату, яка тривалий час вважалася самостійним родом Aglaozonia. Стадія Aglaozonia розмножується за допомогою гаплоїдних зооспор, що виникають у розташованих сорусами одногніздних спорангіях. Зооспори проростають у нові гаметофіти.
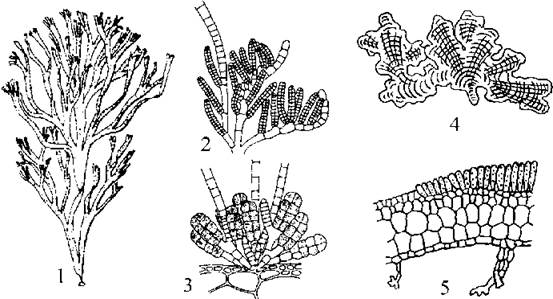
Рис. 16.7. Cutleria: 1 - макроскопічний гаметофіт; 2, 3 - фрагмент гілки з чоловічіми (2) та жіночими (3) гаметангіями; 4 - мікроскопічний спорфіт (стадія Aglaozonia); 5 - поперечний зріз через спорофіт, на поверхні якого розташовується сорус одногніздних спорангіїв (за Newton, 1931; Петров, 1977).
Dictyotales, подібно до попереднього порядку, включає водорості, у яких таломи мають "просту" тканинну будову і виглядають як цілісні чи дихотомічно або віялоподібно розгалужені платівки. Зміна поколінь ізоморфна. Ріст таломів відбувається за рахунок поділу лише однієї верхівкової клітини. Статевий процес оогамний. Звичайно статеві органи зібрані групами і вкриті спільною обгорткою, утворюючи соруси. Характерною специфічною ознакою диктиотальних є відсутність зооспор, нестатеве розмноження відбувається нерухомими тетраспорами. Монадні стадії представлені сперматозоїдами, які мають лише один джгутик. Найбільш поширеними родами є Dictyota, Dilophus та Padina (рис. 15.8).
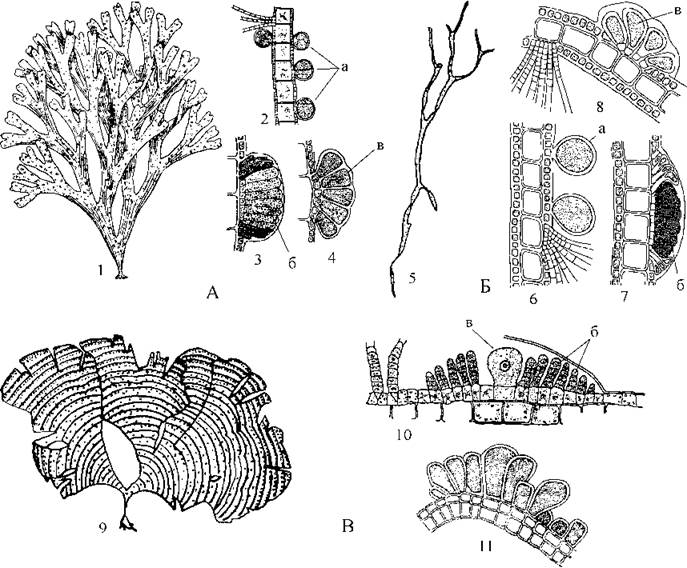
Рис. 16.8. Диктиотальні водорості. А - Dictyota: 1 - зовнішній вигляд талому; 2-4 - поперечні зрізи талому з тетраспорнгіями (2), сорусами антеридіїв (3) та оогоніїв (4). Б - Dilophus: 5 - зовнішній вигляд талому; 6-8 - поперечні зрізи талому з тетраспорнгіями (6), сорусами антеридіїв (7) та оогоніїв (8). В - Padina: 9 - зовнішній вигляд талому; 10 - поперечний зріз талому з оогонієм та антеридіями; 11 - зріз через сорус тетраспорнгіїв (а - тетераспорангії; б - антеридії; в - оогоній) (за Зинова, 1967).
У Dictyota та Dilophus пластинки талому дихотомічно розгалужені, у першого роду вони широкі, тоді як у другого досить вузькі. Таломи Padina мають вигляд цілої або розірваної по краю платівки. На поверхні талому падіни добре помітні концентричні стрічки, утворені скупченнями волосків, а на нижньому боці платівки як правило відкладається шар вапна.
Laminariales об’єднує водорості, спорофіти яких мають вигляд нерозгалужених шнурів або крупних пластин складної тканинної будови. Як правило у серцевині є елементи провідної системи. Ріст таломів інтеркалярний. Зміна поколінь гетероморфна, правильна, статевий процес - оогамія. Гаметофіти мікроскопічні, звичайно у вигляді коротких однорядних розгалужених ниток.
Ламінаріальні водорості є досить важливими у практичному відношенні, оскільки саме до них належить переважна більшість бурих водоростей, які використовуються людиною.
Найбільш відомий представник порядку - Laminaria, або морська капуста. У північних морях масовими видами є L. saccharina та L. digitata, вздовж атлантичного узбережжя Європи - L. hyperborea, на Далекому Сході - L. japonica (рис. 16.9). У субліторальній зоні морів ці водорості утворюють суцільні зарості, які інколи називають "лісами моря".
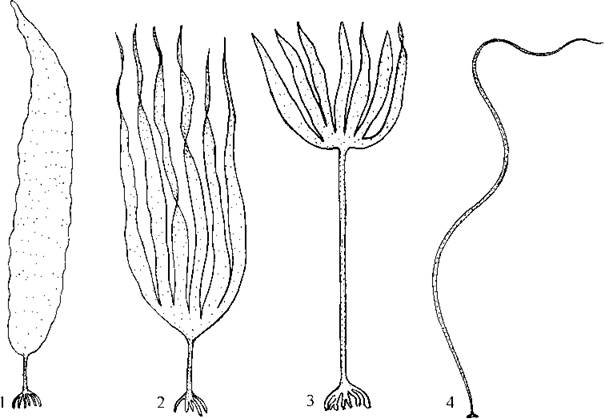
Рис. 16.9. Ламінаріальні водорості: 1 - Laminaria saccharina; 2 - L. digitata; 3 - L. hyperborea; 4 - Chorda filum (ориг.)/
Дорослі спорофіти ламінарії сягають 3-4 м завдовжки і складаються з пластини, "стебла" та розгалуженої системи ризоїдів, за допомогою яких водорость прикріплюється до підводних кам’янистих субстратів. На межі переходу пластини у "стебло" розташовуються меристемні клітини, за рахунок поділу яких відбувається інтеркалярний ріст талому.
Меристема ламінарій активна у холодні пори року, і ріст спорофіту відбувається з січня по квітень-травень у помірних широтах, з лютого по червень-серпень у високих. Поділ клітин меристеми індукується короткою тривалістю світлової фази доби. Це явище отримало назву фотопериодичного відгуку. У напрямку до верхівки талому інтеркалярна меристема відчленовує клітини, що надалі диференціюються у тканини пластини (рис. 16.10), у протилежному напрямку - клітини тканин "стебла". Наприкінці вегетаційного сезону пластина звичайно відмирає, і у наступному році відростає знов, тоді як "стебло" та ризоїди є багаторічними.
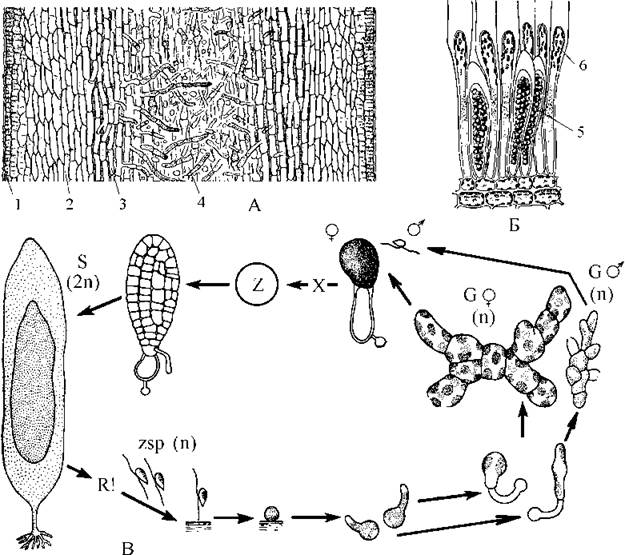
Рис. 16.10. Laminaria: А - поперечний зріз пластини (1 - меристодерма, 2 - кора, 3 - проміжна тканина, 4 - серцевина); Б - сорус зооспорангіїв на поверхні меристодерми (5 - зооспорангій, 6 - захисні видовжені клітини - парафізи); В - схема життєвого циклу (за Петров, 1977).
Зооспорангії ламінарії одногніздні, і утворюються щільними групами (сорусами) з клітин кори стебла восени. Соруси забарвлені у темніший колір, порівняно із стерильною частиною пластини. У кожному спорангію після редукційного поділу утворюється 32 дводжгутикові зооспори. Зооспори, що оточені спільним слизовим міхуром, утвореним переважно альгіновою кислотою, вибухоподібно виштовхуються з пластини у зовнішне середовище. Слизовий міхур розчиняється і зооспори переходять у стан активного руху. Через кілька хвилин вони осідають на субстрат і втрачають джгутики. Надалі половина з зооспор з одного спорангію проростає у чоловічі, а половина - у жіночі гаметофіти.
Чоловічі гаметофіти складаються з кількох десятків клітин, що утворюють коротку, рясно розгалужену нитку. Термінальні клітини бічних гілочок нитки перетворюються на антеридії, в кожному з яких утворюється по одному сперматозоїду.
Жіночі гаметофіти складаються лише з кількох клітин. Будь-яка клітина гаметофіту може розвинутися в оогоній. В ньому утворюється одна яйцеклітина, яка через розрив оболонки оогонію виходить назовні і виділяє феромон ламоксирен, що приваблює сперматозоїди. Після копуляції утворюється зигота, яка без періоду спокою починає проростати у новий спорофіт.
Вздовж узбережжя Америки домінантами багатьох моських фітоценозів є близький до ламінарії рід Macrocystis. Довжина таломів цієї водорості складає до 60 м. Довге тонке стебло під кутом підіймається зі дна до поверхні і несе подвійний ряд довгих (до 1-2 м) "листових" пластин. При основі кожної пластини знаходиться плавальний міхур.
Значна частина ламінаріальних зустрічаються тільки у холодних морях. Наприклад, рід Chorda, що є одним з домінантів морських фітоценозів на нижній межі літоральної зони зустрічається тільки там, де зимова ізотерма не перевищує +12 °С.
Sphacellariales об’єднує водорості з гетеротрихальними багатоосьовими таломами, що мають членисто-мутовчасту будову і виглядають як невеликі розгалужені кущики. Зміна поколінь у сфацеляріальних ізоморфна, статевий процес переважно ізогамний, рідше гетеро- або оогамний. Таломи подібно до диктиотальних наростають внаслідок поділу апікальної клітини.
Типовий представник Sphacellaria (рис. 16.11; див. також рис. 16.4) часто зустрічається як епіфіт на інших водоростях-макрофітах у холодних та теплих морях. Таломи сфацелярії дрібні (в середньому біля 1 см заввишки), і диференційовані на систему висхідних прямостоячих ниток та сланку підошву, інколи - з щільно переплетеними між собою ризоїдами. На верхівці кожної висхідної нитки знаходиться велика апікальна клітина, внаслідок поділу якої відбувається ріст талому. Підошва здатна утворювати столони, на кінцях яких можуть розвиватися нові підошви, від яких, в свою чергу, відростають нові висхідні нитки.
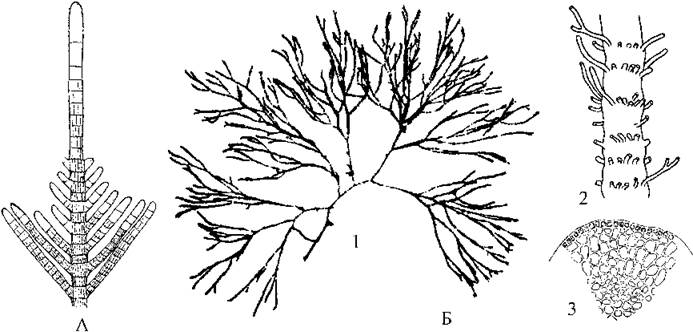
Рис. 16.11. Сфацеляріальні водорості. А - верхівка талому Sphacellaria. Б - Cladostephus: 1 - загальний вигляд талому; 2 - фрагмент талому із залишками мутовчастих гілочок; 3 - поперечний зріз талому (за Петров, 1977; Зинова, 1967).
Апікальна клітина ділиться тільки у поперечному напрямку, відчленовуючи від себе субапікальі клітини. Останні здатні до поділу у поздовжньому напрямку, внаслідок чого виникають мутовки-сифони. Переважна більшість клітин у мутовках до наступних поділів не здатні, проте окремі клітини можуть дати початок клітинам, що функціонально подібні до апікальних - бічним меристемним клітинам. Внаслідок поділу останніх утворюються репродуктивні структури - вивідкові бруньки, короткі бічні гілки з одногніздними спорангіями (на спорофітах) або з багатогніздними жіночими чи чоловічими гаметангіями (на гаметофітах). Статевий процес гетерогамний, зигота проростає без періоду спокою у новий спорофіт. Гамети здатні також партеногенетично проростати у нові гаметофіти, внаслідок чого зміна поколінь вважається неусталеною.
Інший представник - Cladostephus - зустрічається у бентосі на невеликих глибинах (до 2-7 м). Цю водорість часто можна знайти у штормових викидах на узбережжі Чорного моря. Її таломи досить великі (до 20 см заввишки), дихотомічно розгалужені, мають членисто-мутовчасту будову, і на верхівках рясно вкриті численними короткими гілочками, через що здаються булавоподібними (рис. 16.11).
Fucales - це еволюційно наймолодший порядок бурих водоростей. Представники порядку мають складну тканинну будову, переважно оогамний (як вийняток - гетерогамний) статевий процес, диплофазний життєвий цикл без зміни поколінь. У фукальних статеві органи утворюються в особливих порожнинах кори та проміжної тканини, що отримали назву скафідіїв або концептакулів. Нестатевого розмноження спорами у представників порядку немає. Фукальні водорості поширені як у холодних, так і у теплих морях, і звичайно є домінантами фітоценозів літоралі.
У холодних північних морях (наприклад, Білому, Баренцевому, Охотському) дно, що оголюється під час відпливів, часто вкрите суцільним килимом таломів Fucus та Ascophyllum. Таломи Fucusвеликі (30-70 см заввишки), складаються з дископодібної прикріплювальної підошви, циліндричного стебельця, та дихотомічно розгалужених пластин, що розширюються до верхівки. Стебельце у пластинах переходить у центральне ребро. Фотосинтез здійснюють переважно клітини пластин, стебельце та центральне ребро виконують опірну та провідну функції. Поблизу верхівки у більшості видів роду є добре помітні здуття, виповнені повітрям - повітряні міхури. Завдяки їм таломи у воді підтримуються у вертикальному положенні.
Ріст таломів відбувається внаслідок поділу апікальної клітини, що відчленовує від себе клітини субапікальної меристеми. Останні надалі дають початок всім іншим тканинам.
Статеві органи розвиваються на особливих плодущих гілочках - рецептакулах. Ці гілочки, порівняно зі стерильною частиною пластин, потовщені та мають шерхату поверхню. Шерхатість обумовлена наявністю в рецептакулах великих (до 1-3 мм у діаметрі) порожнин - скафідіїв, в яких розвиваються жінчі та чоловічі статеві органи.
Скафідії виповнені слизом, їх внутрішня поверхня вкрита шаром вистілаючих клітин, що дають початок коротким стерильним ниткам, та ініціальним репродуктивним клітинам, з яких утворюються антеридії та оогонії. Скафідії у фукусу двостатеві або роздільностатеві.
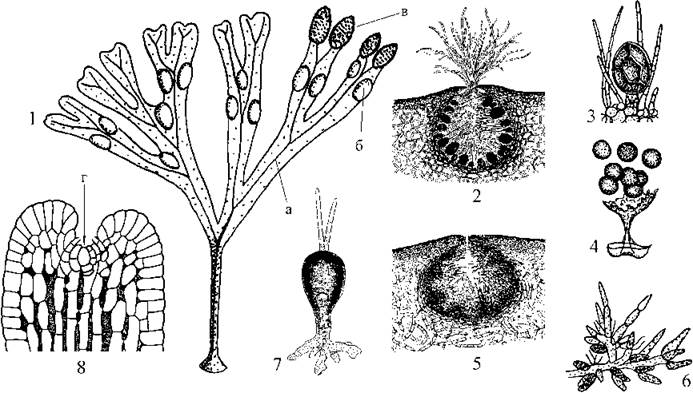
Рис. 16.12. Fucus. 1 - зовнішній вигляд талому (а - центральне ребро, б - повітряний міхур, в - рецептакул зі скафідіями); 2 - жіночій скафідій з оогоніями; 3 - оогоній з вісьма яйцеклітинами; 4 - вихід яйцеклітин з оогонію; 5 - чоловічий скафідій з антеридіальними нитками; 6 - антеридіальна нитка з антеридіями; 7 - проросток; 8 - верхівка талому (г - апікальна клітина) (1 - ориг.; 2-8 - за Петров, 1977).
Коли ініціальна репродуктивна клітина починає розвиватися у статевий орган, її ядро мейотично ділиться. Далі гаплоїдні клітини діляться мітотично і диференціюються або в оогонії, або в антеридії. Оогонії еліпсоїдні, мають коротку одноклітинну ніжку та вісім яйцеклітин. Антеридії утворюються з верхівкових клітин коротких, рясно розгалужених антеридіальних ниток, що являють собою видозмінені парафізи. У кожному антеридії утворюється 64 дводжгутикових сперматозоїда.
Перед заплідненням антеридії та оогонії через отвір скафідію звичайно виштовхуються слизом у зовнішне середовище. Далі оболонки антеридіїв та оогоніїв розпливаються, звільняючи гамети. Яйцеклітини виділяють феромон фукосератин, завдяки якому навколо кожної яйцеклітини швидко концентрується велика кількість сперматозоїдів. Після запліднення зигота без періоду спокою починає проростати у новий талом.
Процес розвитку антеридії та оогоніїв у представників Fucales різні автори інтерпретують по різному. Так, за класичними поглядами (Kylin, 1940; Fritsch, 1945) статеві органи у цих водоростей утворюються внаслідок звичайної гаметичної редукції спеціалізованих клітин талому, і тому життєвий цикл є диплофазним та відбувається без зміни поколінь. На думку інших дослідників (Strasburger, 1906; van den Hoek, Mann, Jahns, 1995), ініціальні клітини, з яких утворюються статеві органи, є гомологами одногніздних спорангіїв, а самі оогонії та антеридіальні нитки - мікроскопічними гаплоїдними гаметофітами. Тому життєвий цикл фукальних гаплодиплофазний, зі споричною редукцією та правильною гетероморфною зміною поколінь. Цікаву гіпотезу висловив Ю.Е. Петров (1986). На його думку, весь шар вистілаючих клітин скафідіїв є видозміненим диплоїдним гаметофітом, і тому життєвий цикл слід розглядати як диплофазний, з гаметичною редукцією та гетероморфним чергуванням поколінь2.
Таломи Ascophyllum ременеподібні, до верхівки не розширюються, а рецептакули утворюються не апікально, а на бічних гілочках.
У Чорному морі на глибинах 1-4 м морські "луки" утворюють види роду Cystoseira (переважно C. barbata та C. crinita). Вони складають основу ланки продуцентів у бентосних фітоценозах літоралі. Таломи Cystoseira мають вигляд кущиків з більш-менш циліндричними рясно розгалуженими висхідними гілками та короткими стеблами. Взимку на восени на верхівках гілок утворюються ланцюжки невеличних повітряних міхурів. Невеличкі рецептакули також розташовуються апікально
У літоральній зоні Атлантичного та Тихоокеанського узбережь домінуючою водорістю є Sargassum. Окремі види цього роду, що мешкають на відкритих акваторіях Атлантики, на відміну від інших бурих водоростей, здатні розмножуватися у неприкріпленому стані і утворюють т.з. Саргасове море. Таломи Sargassum кущисті, і диференційовані на підошву, стебло та основні гілки. Рецептакули та повітряні міхури у саргасу розвиваються на коротких гілочках у пазухах основних гілок.
Головним ценозоутворювачем помірно холодних морів Південної півкулі є Durvillea, що масово розвивається вздовж узбережжя Південної Америки, Австралії та антарктичних островів. Таломи водорості дуже великі (до 10 м заввишки), складаються з підошви, короткого стебла та товстих трубчастих гілок. Ріст відбувається за рахунок як апікальної, так і інтеркалярної меристеми. Скафідії утворюються з кількох клітин меристодерми гілок, що заглиблюються у проміжну тканину та оплетаються меристодермальними нитками сусідніх клітин.

Рис. 16.13. Фукальні водорості: 1, 2 - Ascophyllum (1 - талом, 2 - фрагмент талому з рецептакулами та повітряним міхуром) ; 3, 4 - Cystoseira barbata (3 - фрагмент талому; 4 - верхівка талому зі скафідіями); 5 - Cystoseira crinita (верхівка талому зі скафідіями); 6 - Durvillea (за Зинова, 1953, 1967; Петров, 1977; Fritsch, 1935).
Феромони бурих водоростей
Речовини, що забезпечують хемотаксисний рух сперматозоїдів до яйцеклітин, називають феромонами. У 1963 р. Д. Мюллер, досліджуючи розмноження Ectocarpus siliculosus, звернув увагу на те, що суспензія жіночих гамет, на відміну від чоловічих, має приємний солодкуватий запах. Краплинки олії, насичені парою цієї речовини, приваблювали чоловічі гамети. Для отримання феромону у чистому вигляді було створено апарат, що забезпечував циркуляцію чистого повітря над суспензією жіночих гамет з подальшою конденсацією леткої запашної фракції. Протягом двох років було отримано біля 100 мг досліджуємої речовини. Хімічний аналіз показав, що речовина являє собою фенольну сполуку вуглеводневої природи, яку було названо ектокарпеном.
Далі аналогічні дослідження були проведені з культурами кількох видів роду Fucus, одним видом Sargassum та фукальної водорості Pelvetia. Феромоном, що визначав хемотаксис сперматозоїдів цих видів виявився лінійний вуглеводень, що отримав назву фукосерратену. Цікаво, що у присутності фукосерратену сперматозоїди не відрізняли яйцеклітину "свого" виду від "чужого", концентруючись навколо будь-якої яйцеклітини, яка виділяла цей феромон або була ним оброблена. Розпізнавання "своєї" яйцеклітини відбувалося лише на початкових стадіях копуляції - "чужі" сперматозоїди від яйцеклітини відкріплювались, а один із "своїх" успішно зливався. Тому було припущено, що розпізнавальну функцію у бурих водоростей виконує мембрана яйцеклітини.
Сьогодні у бурих водоростей виявлено біля десяти феромонів, всі вони є циклічними або лінійними вуглеводневими сполуками. Деякі з них, зокрема ламоксирен, виділений з ламінарії, знайшли застосування у технологічному процесі промислового вирощування бурих водоростей.
Практичне значення
Бурі водорості широко використовуються у практичній діяльності людини. Сьогодні в цілому окреслилися три основних напрямки їх застосування: отримання альгінової кислоти та альгінатів, вирощування, збір та переробка їстівних представників, застосування біомаси як органічних добрив, багатих на калій та азот. Основними об'єктами практичного використання є види родів Laminaria, Macrocystis, Fucus, Sargassum, Cystoseira.
Альгінати складають основу клітинних покривів Phaeophyta. Вони мають клеючі та желюючі властивості, і тому використовуються у виробництві лаків та фарб, при виготовлення тканин специфічної текстури, як консерванти у харчовій промисловості. Альгінати (зокрема, альгінат натрію) є такою фармакологічною сировиною, з якої виробляють радіопротекторні препарати.
їстівні водорості або збирають екстенсивно, або штучно вирощують на морських фермах в умовах аквакультури. Головними об’єктами аквакультури є Laminaria saccharina, L.japonica, L. digitata, види роду Macrocystis. Продукти, виготовлені з цих водоростей, відомі під загальною назвою "морська капуста". Наприклад, щорічний урожай ламінарії у 90-х роках складав біля 2 млн.т. З морською капустою проводиться інтенсивна селекційна робота, в тому числі із застосуванням методів генної інженерії, численні біотехнологічні досліди.
Як органічні добрива, багаті на біогенні речовини та мікроелементи, широко використовують Fucus, Cystoseira, Sargassum, Ascophyllum. Ці водорості збирають на морських узбережжях після сильних штормів або в літоральній зоні під час відливів.
Місце в системі
Бурі водорості належать до фотоавтотрофних тубулокристат- страменопілів. За молекулярними даними та комплексом фенотипічних ознак відділ близько споріднений з жовтозеленими водоростями.
1 Систему відділу подано за: L.D. Druehl, C.Mayes, I.H.Tan, G.W. Saunders. Molecular and morphological phylogenesis of kelp and associated brown algae. In: Origin of Algae and their Plastids (ed.D.Bhattacharya). Springer-Verl., Wien - New York, 1997: 221-236; A.Peters, M.Clayton. Molecular and morphological investigation of three brown algal genera with stellate plastids: evidence for Scytothamnales ord. nov. (Phaeophyceae). - Phycologia (1998), 37: 106-113); F. Rousseau, B. de Reviers. Circumscription of the order Ectocarpales (Phaeophyceae): bibliographical synthesis and molecular evidence. - Cryptogamie, Algol. (1999), 20 (1): 5-18; F. Rousseau, B. de Reviers. Phylogenetic relationships within the Fucales (Phaeophyceae) based on combined partial SSU+LSU rDNA sequence data. - Eur.J.Phycol. (1999), 34: 53-64. Не розглядається порядок Ascoseirales(див.: Жизнь растений. Т. 3. Водоросли, лишайники. - М., Просвещение, 1977), представники якого нечисленні і мешкають у морях Південної півкулі.
2 Розгляд аргументів "за" та "проти" кожної з цих гіпотез може бути темою окремого семінарського заняття. Детальніше див.: Петров Ю.Е. Эволюция циклов развития у водорослей. (Комаровские чтения, XXXV). - Л., Наука, 1986. - 61 с.