Альгологія - Костіков І.Ю. - 2009-2013
Розділ 24. Зелені водорості – Chlorophyta
24.2. Хлорофітна лінія
До хлорофітної лінії входять представники класів зелених водоростей, для монадних стадій яких характерний хрестоподібний цитоскелет та мітоз закритого або напіввідкритого типа. Цитокінез відбувається за допомогою фікопласта або за участю кільцевої борозни (виняток - порядок Trentepohliales). Цю лінію представляють чотири класи - Chlorophyceae, Trebouxiophyceae, Ulvophyceae, Siphonophyceae.
24.2.1. Клас Chlorophyceae
Нараховує близько 4 тис. видів водоростей, що мають одноклітинну, багатоклітинну чи неклітинну будову. Хлорофіцієві мешкають переважно у прісних континентальних водоймах, рідше зустрічаються у наземних біотопах. Представники класу різноманітні за будовою джгутикового апарату, мітозом, морфологією, розмноженням, життєвими циклами. Chlorophyceae представляє лінію зелених водоростей, у межах якої цитокінез відбувається за участю фікопласта, а джгутикова коренева система є хрестоподібною з супротивно розташованими або зміщеними за годинниковою стрілкою базальними тілами.
Ознаки та характеристика класу
Біохімічні ознаки
Склад пігментів та продуктів асиміляції є типовим для зелених водоростей в цілому (хлорофіли a та b, всі типи каротинів, ксантофіли лютеїнового ряду, крохмаль як основний продукт асиміляції).
Цитологічні особливості
Клітинні покриви. Клітини деяких одноклітинних хлорофіцієвих водоростей (наприклад, Dunaliella) вкриті лише плазмалемою. Проте переважна більшість видів має справжні оболонки характерної тришарової будови. У монадних форм оболонки звичайно глікопротеїнові, у кокоїдних, багатоклітинних та неклітинних - целюлозно-пектинові. Інколи між проміжним та внутрішнім шарами оболонки міститься додатковий, спорополеніновий шар, що надає покривам міцності та хімічної стійкості.
Субмікроскопічні лусочки як на поверхні клітин, так і на джгутиках у хлорофіцієвих водоростей відсутні.
Ядерний апарат та особливості мітозу і цитокінезу. У монадних форм відсутні центриолі, а їх функцію виконують базальні тіла джгутиків. Це відбувається завдяки структурному зв’язку між ядерною мембраною та базальними тілами, що здійснюється двома тонкими гладенькими ризопластами, побудованими з центрину (рис. 24.15). У нерухомих форм є справжні центриолі (за винятком едогоніальних водоростей).
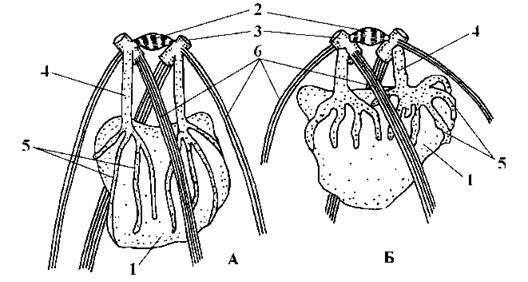
Рис. 24.15. Зв’язок джгутикового апарату з ядром при релаксуючому (А) та скороченому (Б) стані ризопластів у Chlamydomonas: 1 - ядро, 2 - поперечносмугасте з’єднувальне волокно, 3 - базальні тіла, 4 - ризопласти, 5 - розгалуження ризопластів, 6 - мікротубулярні корені (схематизовано за Mattox, Stewart, 1984 та Salisbury, 1989).
Спільна риса всіх хлорофіцієвих - це наявність закритого мітозу та колапсуючого веретна. У межах класу відомо чотири варіанти такого мітозу: а) хламідомонадовий, б) циліндрокапсовий, в) уронемовий, г) едогонієвий (рис. 24.16).
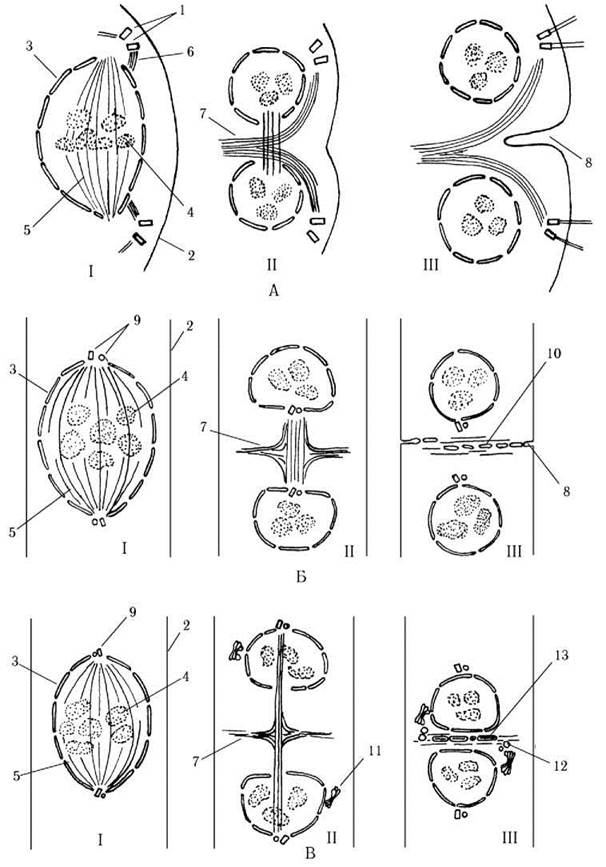
Рис. 24.16. Типи мітозу у хлорофіцієвих водоростей. А - Ch-тип (Chlamydomonas), Б - Cy-тип (Cylindrocapsa), В - Ur-тип (Uronema). І - метафаза, ІІ - анафаза, ІІІ - телофаза. 1 - базальні тіла джгутиків, 2 - плазмалема, 3 - ядерна оболонка, 4 - хромосоми, 5 - веретено, 6 - мікротубулярний джгутиковий корінь, 7 - фікопласт, 8 - кільцева борозна, 9 - центріолі, 10 - пухирці ендоплазматичної сітки, 11 - комплекс Гольджі, 12 - пухирці комплексу Гольджі, 13 - клітинна платівка (за Floyd, Stewart, Mattox, 1972; Triemer, Brown, 1974; Sluiman, 1985).
Хламідомонадовий мітоз (Ch-мітоз) властивий монадним та гемімонадним формам: веретено формується з участю базальних тіл, в анафазі дочірні ядра розходяться, фікопласт утворюється з мікротрубочок джгутикових мікротубулярних коренів, у пізній анафазі та ранній телофазі ядра дещо зближуються за рахунок скорочення залишків інтерзонального веретена, а цитокінез здійснюється кільцевою борозною шляхом схізотомії або схізогонії.
Циліндрокапсовий мітоз (Cy-мітоз) властивий кокоїдним та сифонокладальним формам, а також тим нитчастим представникам, у яких поперечні перегородки не мають первинних пор: веретено формується з участю центриолей, в анафазі дочірні ядра розходяться, центриолі розташовуються на проксимальних полюсах дочірніх ядер і з залишків веретена утворюється фікопласт. У телофазі помітного зближення ядер не спостерігається, а цитокінез здійснюється кільцевою борозною, що росте за рахунок злиття пухирців ендоплазматичної сітки з інвагінацією плазмалеми. Якщо наприкінці телофази пухирці комплексу Гольджі починають активно транспортувати до поверхні плазмалеми матеріал майбутньої клітинної стінки, то після закінчення поділу кожна дочірня клітина вже має свою власну оболонку, тобто має місце схізогонія. У протилежному випадку дочірні клітини лишаються голими. Це спостерігається при утворенні гамет та позбавлених оболонки зооспор.
Уронемовий мітоз (Ur-мітоз) представлений у багатоклітинних водоростей, що мають первинні пори в поперечних перегородках (за винятком едогоніальних водоростей): веретено формується з участю центріолей, в анафазі дочірні ядра розходяться, центріолі розташовуються на дистальних полюсах дочірніх ядер і з залишків веретена утворюється фікопласт. У телофазі ядра помітно зближуються за рахунок скорочення залишків інтерзонального веретена. У фікопласті скупчуються пухирці комплексу Гольджі з матеріалом майбутньої перегородки. Далі вони частково зливаються, утворюючи клітинну платівку, в якій лишаються пори з плазмадесмами, тобто відбувається цитотомія.
Едогоніевий тип (Oe-мітоз) представлений лише в порядку Oedogoniales: центріолі тут відсутні і центрами організації веретена виступають, вірогідно, елементи ендоплазматичного ретикулюму. В анафазі дочірні ядра розходяться, із залишків веретена утворюється фікопласт. У телофазі ядра помітно зближуються за рахунок скорочення залишків інтерзонального веретена. У фікопласті скупчуються пухирці, що відшнуровуються від ендоплазматичної сітки і містять матеріал майбутньої перегородки. Далі пухирці частково зливаються, і, подібно до уронемового типу, утворюється клітинна платівка з первинними порам. Таким чином, за цитокінезом цей тип подібний як до циліндрокапсового (злиття пухирців, що походять від ендоплазматичної сітки, а не від комплексу Гольджі), так і до уронемового (утворення клітинної платівки, зближення ядер у телофазі). Специфічною особливістю цитокінезу є процес росту однієї з дочірніх клітин, пов’язаний з розтягуванням кільцевого потовщення оболонки та утворенням особливих клітинних ковпачків (див. нижче).
Джгутиковий апарат. Монадні клітини хлорофіцієвих водоростей мають 2-4 джгутики або є стефаноконтними. В останньому випадку на передньому кінці клітини знаходиться багато джгутиків, розташованих по колу віночком. Всі джгутики ізоконтні або майже ізоконтні, ізоморфні та ізодинамічні.
Перехідна зона містить лише зірчасту структура. Спіралеподібні стрічки, платівки та циліндри у перехідній зоні відсутні. Базальні тіла відносно короткі. Вони або лежать одне напроти одного (конфігурація 12-6), або зміщені за годинниковою стрілкою (конфігурація 1-7).
Зв’язок між базальними тілами здійснюється поперечно-смугастим волокном. До основи базальних тіл приєднуються також дві короткі поперечносмугасті волокнисті структури, які нагадують ризопласти празинофіцієвих водоростей, проте не з'єднуються ані з ядерною мембраною, ані з плазмалемою. Справжні ризопласти є гладенькими, і приєднуються до верхньої частини базальних тіл. Коренева система хрестоподібна і відповідає формулі 4-2-4-2 (рис. 24.17).
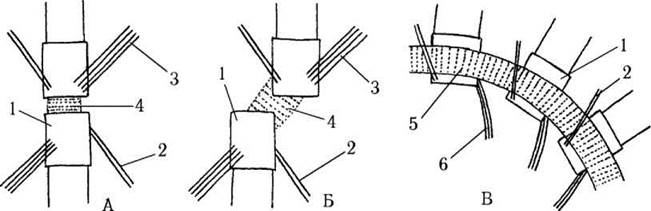
Рис. 24.17. Типи кореневих систем джгутиків хлорофіцієвих водоростей: А - з орієнтацією базальних тіл 12-6; Б - з орієнтацією базальних тіл 1-7; В - стефаноконтний тип. 1 - базальне тіло, 2 - корінь з двох мікротрубочок, 3 - корінь з чотирьох мікротрубочок, 4 - поперечно-смугасте сполучне волокно, 5 - фіброзне кільце, 6 - корінь з трьох мікротрубочок (схематизовано за Kouwets, 1994, Markowitz, 1978).
Особливою кореневою системою, що утворилася з хрестоподібної, є радіальна система стефаноконтних зооспор, сперматозоїдів та андроспор едогоніальних водоростей: на передньому кінці клітини розташовується поперечно-смугасте фіброзне кільце, до якого прикріплюються базальні тіла джгутиків. Від кожного базального тіла починаються два мікротубулярні корені: один з них спрямований вперед і складається з двох мікротрубочок, другий - назад, і містить три мікротрубочки. До кожного мікротубулярного кореня прилягає поперечно-смугастий мікрофібрилярний корінь.
Типи морфологічної структури тіла
У межах класу представлено водорості з усіма відомими типами морфологічної структури, за винятком ризоподіального та тканинного. Найчисленними є таксони з монадним, кокоїдним та нитчастим типами структури.
Монадні форми переважно одноклітинні, рідше - ценобіальні. Серед гемімонадних представників переважають форми, що утворюють масивні слизові колонії, деякі мають редуковані нерухомі джгутики (псевдоцилії). Кокоїдні представники дуже різноманітні і представлені як одноклітнинними, так і колоніальними, у т.ч. ценобіальними формами.
Водорості з ничастим типом структури звичайно виглядають як прості нитки, інколи з загостреною верхівковою та розширеною базальною клітинами. Гетеротрихальні форми переважно мають вигляд розгалужених кущиків, диференційованих на слану та висхідну системи ниток. Деякі об'єднуються загальним слизом у колонії.
Сифональні форми звичайно дрібніші, ніж у інших класах і мають мішкоподібну форму; сифонокладальні види виключно ниткоподібні.
Розмноження та життєві цикли
У межах класу широко представлені всі типи нестатевого та статевого розмноження. Переважають еугамні види з гаплофазним життєвим циклом з зиготичною редукцією, без зміни поколінь. У агамних видів спостерігаються як прості, так і досить складні цикломорфози.
Вегетативне розмноження поділом клітин надвоє шляхом схізогонії відоме у монадних форм, позбавлених клітинної оболонки. Поширеним є розмноження фрагментацією ниток та колоній. Багатьом видам властиве утворення акінет та інших стадій спокою, які одночасно з переживанням несприятливих умов виконують репродуктивну функцію.
Основним способом розмноження є споруляція з утворенням зооспор та апланоспор, рідше - гемізооспор. Зооспори можуть бути або голими (протосифон-тип, п-тип), або вкритими оболонками (хламідомонас-тип, х-тип). Звичайно всі види, що мають зооспори, здатні також розмножуватися за допомогою апланоспор, проте у багатьох автоспорових видів зооспори ніколи не утворюються.
Статеве розмноження у голих форм представлене гологамією та усіма відомими варіантами мерогамії: ізо-, гетеро- та оогамією.
Хоча життєві цикли переважно гаплофазні без зміни поколінь, проте як виняток зустрічаються диплофазні цикли (Chlorococcum diplobionticum), гаплофазні з гетероморфною зміною поколінь (Sphaeroplea) або з яскраво вираженим статевим диморфізмом (нанадрієві види Oedogonium).
Система класу
Молекулярно-філогенетичні реконструкції, що узгоджуються з фенотипічними ознаками, вказають на те, що в межах класу чітко окреслюються три великі групи:
1) одноклітинні, багатоклітинні та неклітинні водорості, у яких базальні тіла джгутиків змішені за годинниковою стрілкою і мають орієнтацію 1-7;
2) одноклітинні, виключно кокоїдні водорості з тенденцією переходу до ценобіального плану будови, у яких базальні тіла не зміщені і мають орієнтацію 12-6;
3) нитчасті та гетеротрихальні водорості, у яких базальні тіла з’єднані фіброзним кільцем, через що монадні стадії стефаноконтні.
Перша група є вихідною, і включає шість порядків - Volvocales, Tetrasporales, Chlorococcales, Cylindrocapsales, Chaetophorales, Sphaeropleales. Всі ці порядки, за винятком Tetrasporales та Chlorococcales, монофілітичні, їх генотипічна спорідненість добре узгоджується з комплексом специфічних морфологічних та цитологічних ознак. Chlorococcales об'єднує водорості двох незалежних еволюційних ліній, що беруть початок від різних вольвокальних. Tetrasporales включає багато незалежних ліній, що є тупими кутами еволюції різних видів, родів та родин вольвокальних водоростей.
Друга група - порядок Scenedesmales - монофілітична, і за класичною системою становить частину Chlorococcales. Проте молекулярні дані свідчать про філогенетичну відокремленість сценедесмальних, а на електронно-мікроскопічному рівні простежується кілька специфічних цитологічних ознак цього порядку.
Третя група також монофілітична і відповідає класичному порядку Oedogoniales.
Основні фенотипічні ознаки порядків подані у таблиці 24.3.
Таблиця 24.3. Основні систематичні ознаки різних порядків Chlorophyceae
Порядок |
Тип структури |
Орієнтація базальних тіл |
Тип зооспор |
Тип мітозу |
Тип цитокінезу |
Volvocales* |
монадний |
1-7 |
х, п |
Ch |
кб |
Tetrasporales* |
гемімонадний |
1-7 |
х, п, - |
Ch |
кб |
Chlorococcales* |
кокоїдний, сарциноїдний, сифональний |
1-7 |
х, п, - |
Cy |
кб |
Cylindrocapsales |
нитчастий |
1-7 |
п, - |
Cy |
кб |
Chaetophorales |
нитчастий та гетеротрихальний |
1-7 |
п, - |
Ur |
пл |
Oedogoniales |
нитчастий та гетеротрихальний |
радіальна |
ст |
Oe |
пл |
Scenedesmales |
кокоїдний |
12-6 |
п, - |
Cy |
кб |
Microsporales |
нитчастий |
12-6 |
п |
? |
? |
Sphaeropleales |
сифонокладальний |
12-6 |
п |
Cy |
кб |
Умовні позначення: тип зооспор: "х" - зооспори з оболонкою (хламідомонас-тип), "п" - зооспори голі (протосифон-тип), "-'' - зооспори відсутні; тип мітозу: "Ch" - хламідомонадовий, "Су" - циліндрокапсовий, "Ur" - уронемовий, "Oe" -едогонієвий; "кб" - за способом кільцевої борозни, "пл" - за способом клітинної платівки. Знаком (*) позначено збірні, парафілетичні порядки.
Volvocales. об’єднує біля 1000 видів монадних водоростей, які вірогідно походять від празинофіцієвих з порядку Chlorodendrales. Клітини мають два-чотири ізоконтні та ізоморфні джгутики, базальні тіла яких зміщені за годинниковою стрілкою. Клітинні оболонки є або відсутні. Вольвокальні водорості бувають одноклітинні (наприклад, Chlamydomonas) або клітини об'єднуються у ценобії різної будови (Pandorina, Volvox). Під час поділу клітин фікопласт утворюється з тих джгутикових коренів, до складу яких входять чотири мікротрубочки.
У межах порядку представлено всі типи мерогамії (ізогамія, гетерогамія, оогамія), у позбавлених оболонок представників статевий процес гологамний. Життєві цикли гаплофазні, з зиготичною редукцією числа хромосом. Зміна поколінь відсутня.
Серед голих одноклітинних водоростей найвідомішим є рід Dunaliella (рис. 24.18), що зустрічається переважно у гіпергалінних водоймах. Dunaliella має два джгутики однакової довжини, хлоропласт з піреноїдом, розмножується поділом надвоє. Статевий поцесгологамія, зигота проростає після періоду спокою. Деякі види (наприклад, Dunaliella salina) здатні до гіперсинтезу β-каротину і спричиняють червоне "цвітіння" ропи солоних озер. Dunaliella salina введена у промислову культуру як сировина для отримання каротину та гліцерину.
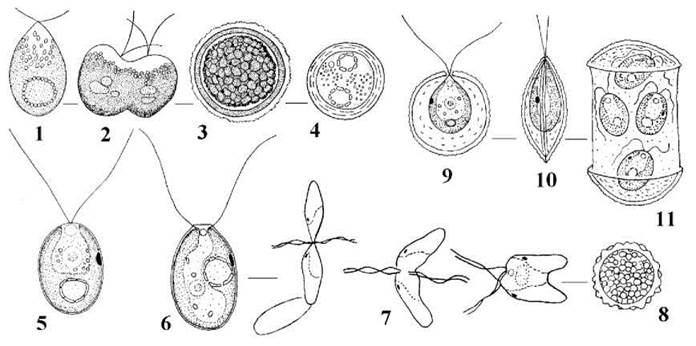
Рис. 24.18. Деякі одноклітинні представники Volvocales: 1-4 - Dunaliella salina (1 - вегетативна клітина, 2 - поділ клітини, 3 - циста, 4 - зигота), 5 - Chamydomonas reinhardtii, 6-8 - Chlamydomonas moewusii (6 - вегетативна клітина, 7 - послідовні стадії копуляції ізогамет, 8 - зигота), 9-11 - Phacotus lenticularis (9 - вегетативна клітина у фронтальній площині, 10 - збоку, 11 - утворення зооспор) (1-4 - Масюк, 1973; 5 - за Ettl, 1983; 6, 7, 9, 10 - ориг.; 8 - за Gerloff, 1940; 11 - за Stein, 1878).
Центральний рід порядку - Chlamydomonas - нараховує біля 500 видів. Клітини вкриті целюлозно-пектиновою оболонкою, мають два джгутики, хлоропласт з піреноїдом, одне ядро (при великих збільшеннях мікроскопу помітно навіть без спеціального забарвлення); звичайно є стигма та дві пульсуючі вакуолі. Розмноження відбувається за допомогою зооспор або статевим шляхом (статевий процес - ізо-, гетеро- та оогамія).
При несприятливих умовах клітина втрачає джгутики і переходить до нерухомого способу життя. У деяких видів перехід до нерухомого стану супроводжується ослизненням оболонки і утворенням пальмел. Тривалість перебування у цьому стані може значно перевищувати тривалість перебування у монадному стані. У нерухомому стані Chlamydomonas розмножується зоо- та гемізооспорами. Поступова редукції монадної стадії при збереженні і переважному розвитку нерухомої стадії обумовила виникнення видів з гемімонадною будовою - водоростей порядку Tetrasporales.
Досить численними є одноклітинні водорості, у яких оболонка просякується солями заліза або марганцю, стає крихкою ї за зовнішнім виглядом нагадує панцир (наприклад, у прісноводної водорості Phacotus).
У деяких родів протопласт місцями значно відстає від оболонки. Цікаво, що у кількох видів цієї групи (Chlorogonium euchlorum, Hyalogonium massukiae) спостерігається статевий процес, подібний до кон’югації. У групі окремі види здатні до гіперсинтезу каротину (Haematococcus pluvialis), і можуть викликати червоне "цвітіння". Зокрема, широко відомі "криваві дощі" та "криваві сніги", обумовлені масовим розвитком гематококу. Дослідження 18S РНК показали, що ця група є спорідненою з багатоклітинним зеленими водоростями, зокрема з родом Uronema.
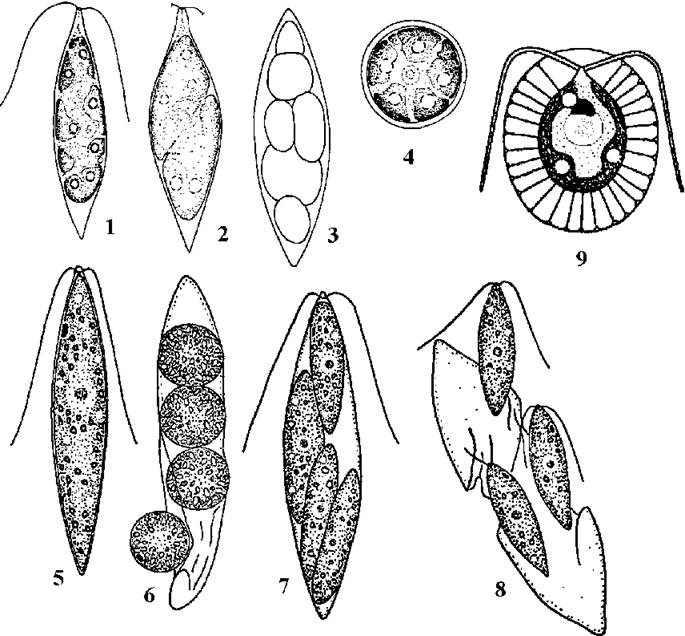
Рис. 24.19. Деякі одноклітинні представники Volvocales: 1-4 - Chlorogonium (1 - вегетативна клітина, 2 - початок утворення зооспор, 3 - гаметангій з протопластами гамет різного розміру, 4 - акінета), 5-8 - Hyalogonium (5 - вегетативна клітина, 6 - гемізооспори, 7 - зооспорангій, 8 - вихід зооспор), 9 - Haematococcus (1-4 - Костіков, Солоненко, 1991; 5-8 - Костіков, 1987; 9 - за Blochmann, 1886).
У ценобіальних форми клітини з'єднуються або власними оболонками, без участі слизу (Spondylomorum, Pyrobotrys), або утримуються разом спільною слизовою обгорткою - інволюкром (Pandorina, Gonium, Volvox).
Gonium має ценобії у вигляді платівки, що, залежно від виду, складається з 4-х або 16 клітин. При нестатевому розмноженні кожна клітина два - вісім разів ділиться, і дочірні клітини ще всередині материнської оболонки складаються у новий ценобій. Статевий процес ізогамний: з кожної вегетативної клітини утворюється 16 однакових гамет, що попарно копулюють. Зигота одягаються товстою гладкою оболонкою і переходить у стан спокою. По його завершенні відбувається мейоз, утворюються чотири гаплоїдні клітини, які ще у зиготі складаються у дочірній ценобій.
У роду Pandorina ценобії овальні, містять 16 або 32 клітини, що щільно прилягають одна до одної і через це набувають гранчастої форми. Нестатеве розмноження відбувається так само, як і у Gonium. Статевий процес - гетерогамія. В клітинах чоловічих ценобіїв утворюються дводжгутикові сперматозоїди, в жіночих - малорухливі яйцеклітини. Після запліднення зигота одягається товстою гладкою оболонкою, накопичує велику кількість гематохрому і переходить у стан спокою. По закінченні періоду спокою у зиготі відбувається мейоз, з чотирьох гаплоїдних ядер три дегенерують, а одне проростає великою зооспорою, у якій через деякий час відбувається серія мітозів і дочірні клітини складаються у новий ценобій.
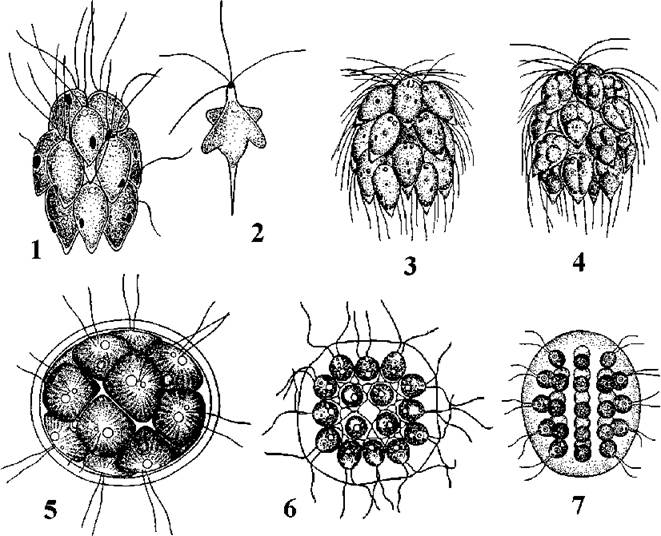
Рис. 24.20. Деякі ценобіальні представники Volvocales: 1, 2 - Pyrobotrys (1 - ценобій, 2 - планзигота); 3, 4 - Spondylomorum (3 - ценобій, 4 - утворення дочірніх ценобіїв), 5 - Pandorina; 6 - Gonium; 7 - Eudorina(1, 2 - ориг., 3, 4 - за Stein, 1878; 5 - за Conrad, 1913; 6 - за Hartmann, 1924; 7 - за Ehrenberg, 1831).
Рід Volvox має ценобії у вигляді порожньої кулі, що складається з 500 - 50 000 дрібних вегетативних дводжгутикових клітин, з’єднаних між собою тонкими плазмадесмами. Крім вегетативних, у ценобії знаходяться також інші типи клітин: спеціалізовані клітини нестатевого розмноження - партеногонідії, та клітини, що утворюють статеві органи - оогонії та антеридії.
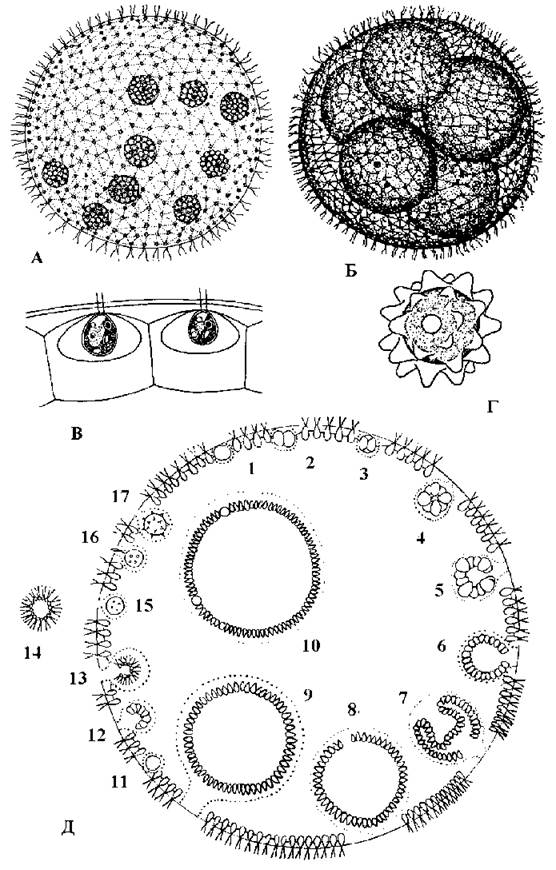
Рис. 24.21. Будова та схема розвитку Volvox: А - ценобій з партеногонідіями, Б - ценобій з дочірніми ценобіями, В - вегетативні клітини, Г - зигота, Д - схема розвитку дочірнього ценобію (1 - клітина партеногонідію, 2-5 - стадії поділу партеногонідію, 6 - ембріональний ценобій з первинною порожниною, 7 - вивертання ценобію, 8-10 - розвиток дочірнього ценобію з вторинною порожниною), утворення сперматозоїдів (11-14), яйцеклітина (15), статевий процес (16) та зигота (17) (А, Б, В - Коршиков, 1938; Г, Д - за Smith, 1955).
Партеногонідії здатні до вегетативного поділу, внаслідок якого з них утворюються дочірні ценобії. Кожний оогоній дає початок одній яйцеклітині, а кожний антеридій - багатьом сперматозоїдам. Після статевого процесу зигота одягається товстою шипастою оболонкою і переходить у стан спокою. По його завершенні ядро зиготи редукційно ділиться. Три дочірні ядра дегенерують, життєздатним лишається тільки одне ядро. Як наслідок зигота проростає однією зооспорою, яка після серії поділів розвивається у новий ценобій.
Tetrasporales. Порядок об’єднує водорості з гемімонадною будовою тіла. Клітини звичайно асиметричні та гетерополярні, на передньому кінці зберігають скоротливі вакуолі, інколи - стигму та нерухомі видозмінені джгутики - псевдоцилії, і часто утворюють слизові колонії різної будови. Розмножуються тетраспоральні водорості фрагментацією колоній, х- та п-зооспорами, зрідка - статевим шляхом. Джутиковий апарат, мітоз, спосіб утворення фікопласта такі ж, як і у вольвокальних водоростей. Статевий процес переважно ізо- або гетерогамний, як виняток - оогамний. Життевий цикл у еугамних видів гаплофазний, з зиготичною редукцією, без зміни поколінь.
Тетраспоральні водорості є збірним таксоном, який об"єднує кілька різних ліній; кожна з цих ліній походить від різних груп вольвокальних водоростей. Тетраспоральні водорості добре ілюструють різноманітність шляхів, за якими відбувалися переходи до типово рослинного нерухомого способу життя. На прикладі цього порядку можна побачити принаймні чотири такі шляхи (рис. 24.22):
1. Джгутики монадних форм починають виконувати функції органів прикріплення до субстрату і тому втрачають рухливість, перетворюючись на псевдоцилії (наприклад, у Chaetochloris та Dicranochaete);
2. Монадні клітини втрачають джгутики і без виділення слизу переходять у нерухомий стан (наприклад, у нейстонного Nautococcus);
3. Монадні клітини втрачають джгутики, на одному кінці клітини виділяють невелику кількість слизу, за допомогою якого прикріплюються до субстрату (наприклад, Chlorophysema);
4. Монадні клітини переходять у пальмелевидний стан і утворюють слизові колонії. У деяких родів редуковані джгутики зберігаються у вигляді псевдоцилій (наприклад, у Tetraspora), або повністю втрачаються (наприклад, у Palmellopsis).
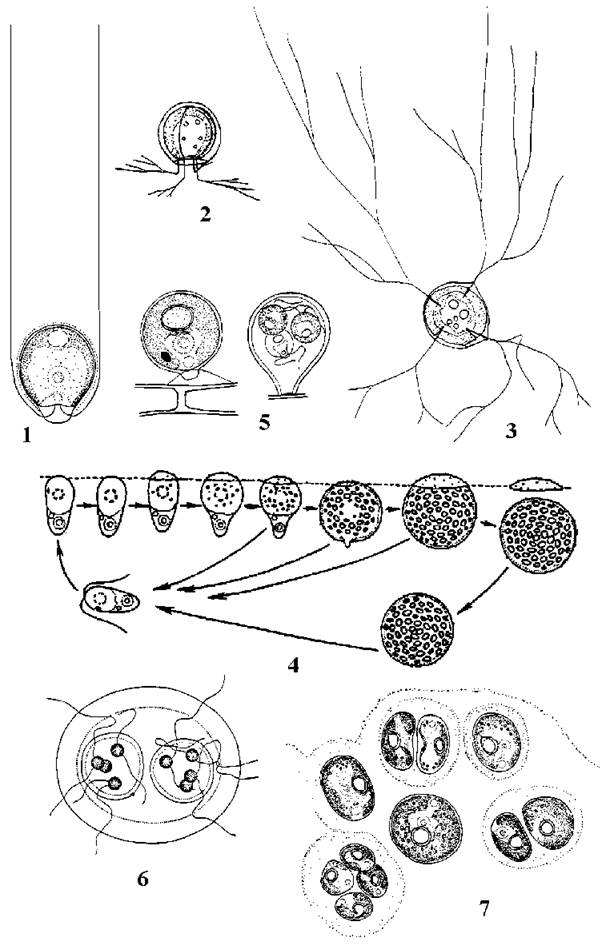
Рис. 24.22. Деякі представники Tetrasporales: 1 - Chaetochloris; 2 - молода (2) та доросла (3) вегетативні клітини Dicranochaete; 4 - цикл розвитку Nautococcus; 5 - вегетативна клітина та зооспорангій Chlorophysema; 6 - колонія Tetraspora; 7 - фрагмент колонії Palmellopsis (1, 5, 6 - за Коршиков, 1953; 2-4, 7 - ориг.).
Chlorococcales. Порядок об’єднує кокоїдні, сарциноїдні та сифональні водорості. В межах порядку більшість представників існує у вигляді поодиноких клітин, хоча відомі також колоніальні форми. Цікаво, що ценобіальні кокоїдні водорості, що за класичною системою відносились до Chlorococcales, за молекулярно-біологічними та цитологічними ознаками виявились самостійною монофілітичною лінією Chlorophyceae, і склали окремий порядок Scenedesmales. Хлорококальні водорості є переважно одноядерними, проте у частини родів простежується тенденція переходу до багатоядерності, яка привела до виникнення сифонального типу структури тіла.
Монадні стадії представлені зооспорами або гаметами, що належать до х- або п-типу, мають два або чотири джгутики, базальні тіла орієнтуються за типом 1 -7. Під час мітозу та цитокінезу фікопласт утворюється із залишків веретена, а кільцева борозна росте внаслідок злиття пухирців ендоплазматичної сітки (Cy-мітоз).
Вегетативне розмноження у колоніальних форм здійснюється фрагментацією колоній. Нестатеве розмноження відбувається зооспорами або апланоспорами. Статевий процес досить рідкісний, переважно ізогамний, зрідка гетеро- або оогамний. Життєвий цикл гаплофазний, хоча відомі також представники з диплофазним циклом. Чергування поколінь не виявлене.
Chlorococcales, так само, як і Tetrasporales, є збірною групою. У межах цього порядку за молекулярними ознаками виявлено дві лінії: перша представляє водорості кокоїдного та сарциноїдного типів структур, що походять від вольвокальних водоростей з зооспорами х-типу та зберігають атрибути гемімонадної організації у вигляді скоротливих вакуолей; у другій лінії вихідним типом монадних клітин є п-тип, скоротливі вакуолі у вегетативних клітинах відсутні, а крім кокоїдних та сарциноїдних представників відомі ще й сифональні форми (рис. 24.23).
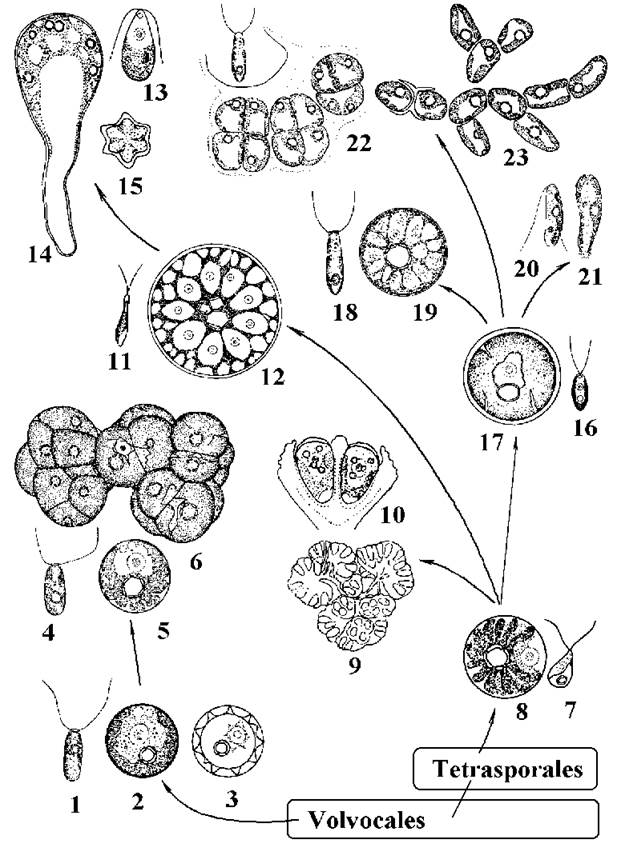
Рис. 24.23. Характерні представники Chlorococcales та напрямки еволюції порядку за молекулярними ознаками: 1-3 - Chlorococcum hypnosporum (1 - зооспора, 2 - вегетативна клітина, 3 - гіпноспора); 4-6 - Tetracystis aeria (4 - зооспора, 5 - окрема вегетативна клітина, 6 - комплекси тетрад); 7, 8 - Ettlia minuta (7 - зооспора, 8 - вегетативна клітина); 9-10 - Botryococcus braunii (9 - колонія, 10 - вегетативні клітини); 11-12 - Spongiochloris (11 - зооспора, 12 - вегетативна клітина); 13-15 - Protosiphon botryoides (13 - зооспора, 14 - вегетативна клітина, 15 - зигота); 16, 17 - Chlorococcum oleofaciens (16 - зооспора, 17 - вегетативна клітина); 18, 19 - Deasonia multinucleata (18 - зооспора, 19 - вегетативна клітина); 20, 21 - Chlamydopodium vacuolatum (20 - зооспора, 21 - вегетативна клітина); 22, 23 - Pleurastrum insigne (22 - зооспора та клітини у сарциноїдному стані, 23 - ниткоподібний стан) (за Коршиков, 1955; Зауер, 1977; Андреева, 1998; Ettl, Gartner, 1995; та ориг. даними).
Характерними представниками першої лінії є Chlorococcum hypnosporum, Ch. diplobionticum та Tetracystis aeria. Всі ці представники мешкають у ґрунтах.
Клітини Chlorococcum hypnosporum сферичні, мають одне ядро, великий пристінний хлоропласт з піреноїдом, дві скоротливі вакуолі. Нестатеве розмноження відбувається дводжгутиковими зооспорами х-типу або гемізооспорами. Зооспори за ультроструктурою (зокрема - за будовою оболонки) подібні до вегетативних клітин хламідомонад. При несприятливих умовах вегетативні клітини одягаються товстою шипуватою оболонкою і перетворюються на спочиваючі клітини - гіпноспори. Статевий процес ізогамний, життєвий цикл гаплофазний з зиготичною редукцією. Інший вид цього роду - Chlorococcum diplobionticum - є морфологічним двійником Ch. hypnosporum, проте має диплофазний життєвий цикл з гаметичною редукцією.
Tetracystis aeria має вегетативні клітини та зооспори, подібні до двох попередніх видів. Проте клітини цієї водорості здатні до вегетативного поділу, внаслідок чого утворюються тетради та комплекси тетрад. Таким чином, в межах першої лінії спостерігається тенденція переходу від кокоїдних до багатоклітинних сарциноїдних форм при збереженні ознак монадної організації, зокрема скоротливих вакуолей. Лінія тісно пов"язана з вольвокальними водоростями, зокрема, з деякими видами роду Chlamydomonas (Ch. reinhardtii, Ch. moewusii).
У межах другої лінії простежується одразу кілька тенденцій: втрата зооспор та перехід до колоніальної будови (лінія Ettlia-Botryococcus), перехід від зооспор п-типу до х-типу (Ettlia-Chlorococcum), переходи від одноклітинної до багатоклітинної (Chlorococcum-Pleurastrum) або сифональної будови (Spongiochloris-Protosiphon; Chlorococcum-Deasonia).
Ettlia minuta має одноядерні сферичні клітини з пристінним зірчасто розсіченим хлоропластом та майже центральним піреноїдом. Розмноження відбувається переважно за допомогою голих дводжгутикових зооспор, рідше - апланоспор. Мешкає у ґрунтах тропічних регіонів.
Botryococcus braunii являє собою колоніальну водорість. Колонії мають вигляд грона, і утворені хрящуватим слизом, на периферії якого розташовуються яйцеподібні клітини. Останні містять пристінний глибоко розсічений хлоропласт з піреноїдом, що позбавлений крохмальної обгортки. У цитоплазмі накопичується велика кількість безбарвної або забарвленої у жовтогарячий колір олії, а назовні клітини виділяють вуглеводневі сполуки. Розмноження відбувається автоспорами.
Водорость мешкає у прісноводному планктоні, і у субтропічних та тропічних регіонах часто спричинює "цвітіння" води. B. braunii у деяких країнах введений у промислову культуру як продуцент вуглеводнів та сировини для виробництва пального.
Повний ряд переходів від одноядерної Ettlia minuta до справжніх сифональних форм демонструють ґрунтові водорості Spongiochloris та Protosiphon. Обидва роди мають голі дводжгутикові зооспори (п-тип, протосифоновий тип - за одним з цих родів), клітини з сітчастими хлоропластами та піреноїдами. У Spongiochloris llanoensis клітини сферичні, дрібні (до 25-35 мкм), одноядерні; інший вид - S.typica - має також сферичні, проте більші клітини (до 70 мкм), які швидко стають багатоядерними. У S.irregularis клітини, які менші від 20 мкм, є сферичними та одноядерними; далі вони стають багатоядерними, і поступово втрачають правильну форму, сягаючи 120 мкм.
Protosiphon botryoides на початкових та середніх стадіях онтогенезу проходить всі стадії розвитку Spongiochloris irregularis, а на пізніх - розвиває багатоядерний ризоїдальний талом. Він диференційований на довгий безбарвний ризоїд (до 3 мм завдовжки та до 30-50 мкм завш), та субсферичну верхівкову частину з сітчастим хлоропластом, кількома піреноїдами та кількома десятками ядер; діаметр верхівкової частини може сягати 500 мкм. При несприятливих умовах утворюються товстостінні акінети, схожі на дорослі клітини Spongiochloris. За наявності води відбувається ізогамний статевий процес, утворюється зірчаста зигота, яка проростає після періоду спокою.
Chlorococcum oleofaciens відноситься до тієї групи видів роду Chlorococcum, які позбавлені пульсуючих вакуолей. Вегетативні клітини одноядерні, сферичні, з розсіченим парієтальним хлоропластом. У цитоплазмі при старінні накопичується велика кількість олії. При нестатевому розмноженні утворюються дводжгутикові х-зооспори, які, проте, за ультараструктурою дещо відрізняються від зооспор Ch.hypnosporum та клітин хламідомонад (зокрема, в оболонці відсутній глікопротеіновий шар). Відомі також апланоспори.
За молекулярними ознаками, які корелюють також з морфологічними та деякими цитологічними ознаками, Chlorococcum oleofaciens більш споріднений з Ettlia minuta, ніж з Chlorococcumhypnosporum та Ch.diplobionticum. Це добре ілюструє парафілетичний характер як порядку в цілому, так і окремих родів.
Рід Deasonia нагадує Chlorococcum oleofaciens, проте має хлоропласт губчастого типу. Дорослі клітини стають багатоядерними, і таким чином, демонструють тенденцію перехіду до сифональної будови у представників з зооспорами х-типу.
Pleurasrtum insigne - багатоклітинна ґрунтова водорість, що складається з комплексів сарциноподібних пакетів, що інколи утворюють ниткоподібні структури. Розмноження відбувається фрагментацією агрегатів, а також за допомогою дводжгутикових зооспор х-типу2. При несприятливих умовах утворюються акінетоподібні клітини з шипуватою оболонкою.
Cylindrocapsales. Порядок об’єднує водорості, що мають нитчастий тип морфологічної структури тіла, циліндрокапсовий тип мітозу та цитокінез, що відбувається за участю кільцевої борозни. За цитологічними ознаками циліндрокапсальні водорості дуже близьки до Chlorococcales (див. табл. 24.3), проте чітко відрізняються від них здатністю утворювати нерозгалужені нитки. Типовим представником є рід Cylindrocapsa.
Таломи циліндрокапси представлені нерозгалуженими нитками, що не розпадаються на поодинокі клітини. Кожна клітина одягнена товстою шаруватою оболонкою, містить масивний центральний хлоропласт з піреноїдом, одне ядро. Пори та плазмадесми між сусідніми клітинами відсутні, і через це водорість можна вважати дійсно багатоклітинною лише умовно. Розмноження відбувається за допомогою голих дводжгутикових зооспор (п-тип), що можуть утворюватись у будь-якій клітині нитки.
Статевий процес оогамний. Оогонії утворюються внаслідок розростання вегетативних клітин. Антеридії виникають після серії швидких поділів вегетативної клітини, в кожному антеридії формується по 1-2 сперматозоїди. Після запліднення зигота одягається товстою оболонкою і переходить у стан спокою. По його завершенні відбувається мейоз, утворюються чотири дочірні клітини, кожна з яких здатна прорости у нову нитку.
Cy-тип мітозу виявлений також у нитчастих водоростей з родів Radiofilum, Geminella, Binuclearia (рис. 24.24).
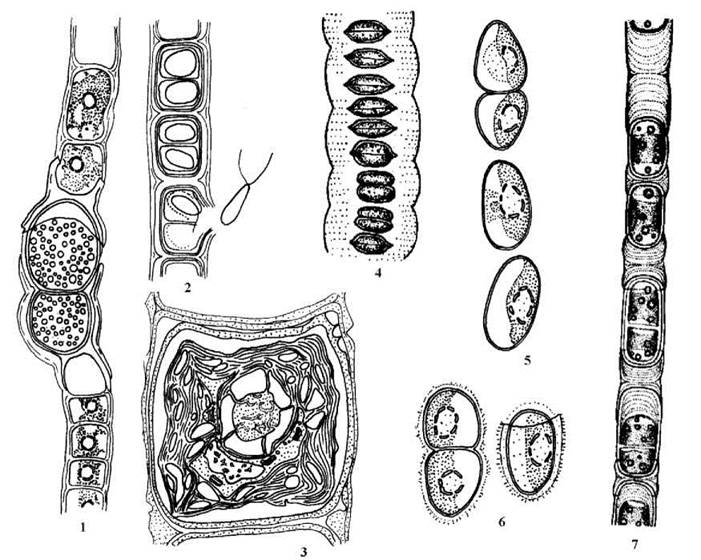
Рис. 24.24. Характерні представники Cylindrocapsales: 1-3 - Cylindrocapsa geminella - нитки з оогоніями (1) та антеридіями (2), ультратонка будова клітини (3); 4 - Radiophylum mesomorphum; 5 - Geminellaterricola без забарвлення (5) та після забарвлення (6) розчином метиленового синього; 7 - Binuclearia tectorum (1, 2, 5, 6 - ориг., 3 - за Kouwets, 1994; 4 - за Skuja, 1955; 7 - за Wichmann, 1937).
Порядок вважається монофілітичним, і виводиться від хлорококальних водоростей з голими зооспорами п-типу.
Chaetophorales. Об’єднує справжні багатоклітинні (нитчасті та гетеротрихальні) водорості, що мають первинні пори, які утворюються внаслідок уронемового типу мітозу. Цитокінез відбувається за участю клітинної платівки, базальні тіла джгутиків зміщені за годинниковою стрілкою. Всі хетофоральні водорості мешкають у перифітоні прісних континентальних водойм.
За результатами цитологічних, молекулярно-біологічних та біохімічних досліджень, Chaetophorales являє собою монофілітичну групу. Дослідження ядерного гену, що кодує рибосомальну РНК, показали, що пращурами порядку є вольвокальні водорості, споріднені з Chlorogonium.
В межах порядку простежується поступове ускладнення плану будови тіла: від нитчастих нерозгалужених водоростей до гетеротрихальних представників, що складно диференційовані у морфологічному та функціональному відношенні. Цей ряд добре простежується на прикладі родів Uronema, Stigeoclonium, Draparnaldia (рис. 24.25).
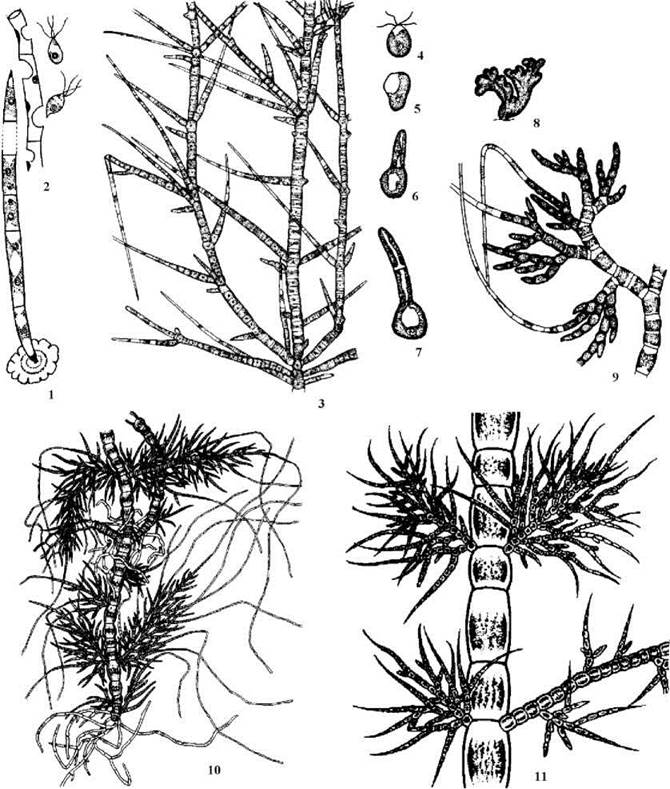
Рис. 24.25. Деякі представники Chaetophorales: 1, 2 - Uronema (1 - вегетативна нитка, 2 - вихід зооспор із спорангія); 3 - 7 - Stigeoclonium (3 - фрагмент висхідної частини талому, 4 - зооспора, 5-7 - стадії проростання зооспори); 8, 9 - Chaetophora (8 - зовнішній вигляд талому, 9 - фрагмент талому з хетами); 10, 11 - Draparnaldia (10 - сланкі, опірні, асиміляторні нитки та хети, 11 - фрагмент висхідної частини з опірними та асимілятроними нитками) (за Мошкова, 1986).
Uronema має талом у вигляді простої нерозгалуженної нитки, що складається з морфологічно та функціонально однакових клітин (за винятком базальної та апікальної). Кожна клітина одноядерна, має пристінний хлоропласт у вигляді незамкненого кільця, в якому розташовується один (рідше - два-чотири) піреноїди. Базальна клітина утворює диск, за допомогою якого водорість прикріплюється до підводних субстратів. Апікальна клітина має звужену та загострену верхівку. Розмноження відбувається за допомогою чотириджгутикових зооспор, або апланоспорами.
Stigeoclonium має гетеротрихальні таломи, що складаються з розгалужених висхідних та сланких ниток. Перші здійснюють функції фотосинтезу та розмноження, за допомогою других водорість прикріплюється до субстрату. Висхідні нитки на верхівці звужені, і досить часто закінчуються одно- або багатоклітинними тонкими безбарвними волосками - хетами. Рясність галуження та кількість хет залежить від концентрації азоту у середовищі: якщо у воді міститься велика кількість азоту, галуження зустрічається зрідка, і хети майже відсутні (в цьому випадку водорість нагадує Uronema), і навпаки. Нестатеве розмноження відбувається чотириджгутиковими зооспорами та апланоспорами, статевий процес - ізогамія і здійснюється дводжгутиковими гаметами. Цікаво, що гамети здатні проростати партеногенетично, і таким чином, виконувати також функції зооспор. Життєвий цикл гаплофазний, з зиготичною редукцією і буз зміни поколінь.
Близьким до Stigeoclonium є рід Chaetophora. Таломи хетофори майже повністю ідентичні до попереднього роду, проте занурені у масивний колоніальний слиз.
Таломи Draparnaldia диференційовані на три системи ниток - сланкі, висхідні опірні та асимілятори. Сланки нитки та асимілятори нагадують аналогічні нитки Stigeoclonium, і виконують функції прикліплення до субстрату та фотосинтезу і розмноження, відповідно. Опірні нитки підносять над субстратом всю масу асиміляторів. Клітини опірних ниток мають редукований хлоропласт, товсті клітинні стінки, і не здатні до утворення зооспор, гамет та інтенсивного фотосинтезу.
Oedogoniales. Порядок об’єднує нитчасті та гетеротрихальні водорості, які за цитокінезом з участю клітинної платівки нагадують хетофоральні водорості, проте суттєво відрізняються від них наявністю особливого типу мітозу (едогонієвий мітоз), клітинним поділом з утворенням ковпачків, стефаноконтними монадними клітинами (зооспорами, андроспорами та сперматозоїдами) та високорозвиненим оогамним статевим процесом. До порядку входить біля 500 видів водоростей трьох родів - Oedogonium, Bulbochaete та Oedocladium.
Клітинний поділ у Oedogoniales. На початку поділу біля верхівки клітини з’являється складка з диктіосом, які, зливаючись утворюють увігнуте в цитоплазму кільце. Далі ядро мітотично ділиться. Між дочірніми ядрами за способом фікопласту починає формуватись клітинна перегородка, материнська оболонка у зоні кільця розривається. Кільце починає швидко розпрямлятися, за рахунок чого відбувається ріст верхньої нової клітини. Залишки оболонки материнської клітини лишаються на дочірній у вигляді ковпачка (рис. 24.26). Кількість ковпачків відповідає кількості поділів даної клітини.
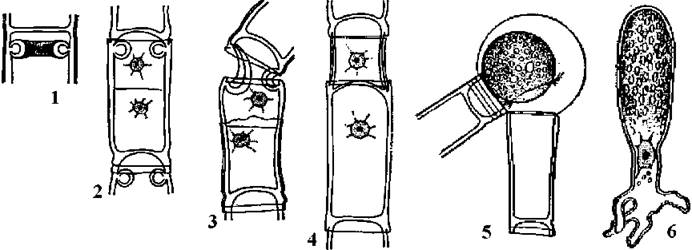
Рис. 24.26. Клітинний поділ (1-4), вихід зооспори (5) та її проростання (6) у едогоніальних водоростей (за Виноградова, 1977).
Монадні клітини едогоніальних водоростей стефаноконтні і можуть бути представлені трьома типами: а) зооспорами, що утворюються по одній в клітині- спорангію; б) андроспорами, з яких виростає особлива мікроскопічна чоловіча рослина - нанандрій; андроспори утворюються по одній, як і зооспори; в) сперматозоїдами, що розвиваються в антеридії по два. Встановлено, що після руйнування оболонок протопласти звичайних вегетативних клітин здатні виробляти джгутики та перетворюватись на зооспори.
Едогоніальні водорості можуть бути однодомними або дводомними. Життєві цикли у едогоніальних водоростей бувають двох типів: гаплофазні без зміни поколінь та гаплофазні з стадією мікроскопічного чоловічого гаметофіту - нанандрію (рис. 24.27). Види, які мають перший тип життєвого циклу називають макрандрієвими, другий - нанандрієвими.
У макрандрієвих видів як антеридії, так і оогонії розвиваються на звичайних вегетативних нитках. В антеридіях формується по два сперматозоїди, в оогонії - одна яйцеклітина. Сперматозоїди виходять з антеридію, підпливають до оогонію, через пору або тріщину в оболонці проникають до яйцеклітини та запліднюють її. Зигота одягається товстою оболонкою і переходить у стан спокою. По закінченні періоду спокою в зиготі відбувається редукційний поділ, з кожного гаплоїдного ядра утворюється зооспора, яка надалі проростає у нову нитку.
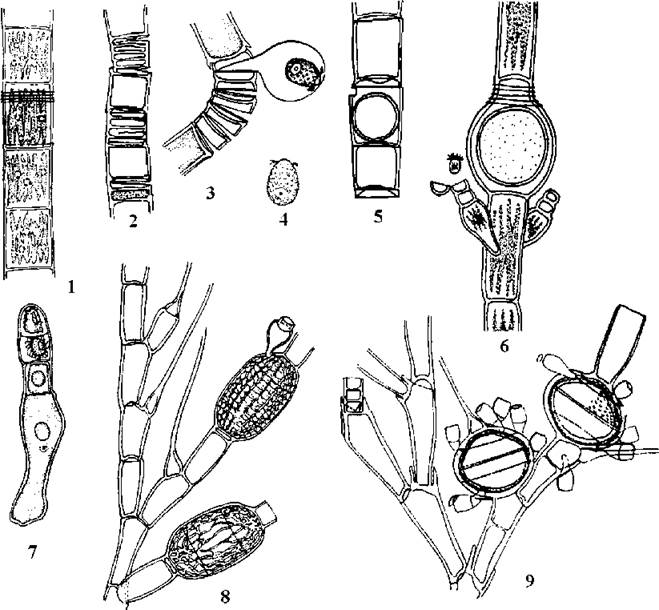
Рис. 24.27. Характерні представники Oedogoniales. 1-7 - Oedogonium: 1 - вегетативна нитка, 2 - нитка з антеридіями (макрандрієвий вид), 3 - вихід сперматозоїду з антеридію, 4 - сперматозоїд, 5 - нитка з оогонієм, 6 - нитка з оогонієм та двома нанандріями (нанадрієвий вид), 7 - нанандрій з верхівковим антеридієм, в якому утворюються сперматозоїди. 8-9 - Bulbochaete: фрагменти таломів з хетами, оогоніями та нанандріями (1, 6 - ориг., 2, 5, 7-9 - за Юнгер, 1993; 3, 4 - за Виноградова, 1977).
У нанандрієвих видів на нитках розвиваються оогонії та особливі спорангії, в яких утворюється чоловіча зооспора - андроспора. Вона підпливає до оогонію, осідає на нього або на сусідню клітину та проростає у 2-4-клітинний нанандрій. Верхівкова клітина нанандрію перетворюється на антеридій, в якому розвиваються два сперматозоїди. Далі цикл розвитку відбувається так само, як і у макрандрієвих видів.
Найчисленнішим родом порядку є Oedogonium (біля 500 видів), для якого характерні нитчасті нерозгалужені таломи. У роду Bulbochaete (більш 100 видів) нитки розгалужені, на кінцевих гілках, як правило, утворюються бульбоподібно потовщені при основі хети. Oedogonium та Bulbochaete широко розповсюджені у перифітоні та бентосі прісних водойм, нерідко відриваючись від субстрату утворюють м’які неслизькі тини на поверхні води. Рід Oedocladium має таломи гетротрихальної будови, диференційовані на підземні сланки та надземні висхідні нитки. Види цього роду мешкають у ґрунтах тропічних регіонів.
Scenedesmales. У класичних системах всі водорості цього порядку відносили до Chlorococcales, і лише молекулярні та електронно-мікроскопічні дослідження 90-х років показали еволюційну самостійність та філогенетичну відокремленість цієї групи. Порядок об’єднує переважно прісноводні планктонні та наземні ґрунтові водорості з кокоїдним типом структури.
Монадні клітини (зооспори та гамети) позбавлені клітинної оболонки (п-тип), мають особливу орієнтацію джгутиків - базальні тіла розташовуються одне навпроти одного і не зміщені ані за, ані проти годинникової стрілки (12-6 орієнтація). При вигляді збоку базальні тіла виглядають майже паралельними одне до одного, що також суттєво відрізняє Scenedesmales від Chlorococcales. Значна кількість видів порядку втратила монадні стадії, і належність таких представників до Scenedesmales встановлена за результатами молекулярно-філогенетичних досліджень. За мітозом та цитокінезом сценедесмальні не відрізняються від хлорококальних водоростей.
За результатами електронно-мікроскопічних досліджень цитоскелету, пращурами сценедесмальних вважають одноклітинні вольвокальні та тетраспоральні водорості зі злегка неоднаковими джгутиками, що нагадують роди Heterochlamydomonas та Heteroteracystis, відповідно.
В межах порядку простежуються дві основні тенденції: переходи до багатоядерного та ценобіального планів будови. Серед сценедесмальних сьогодні виявлено три еволюційні лінії: а) лінію одноклітинних та ценобіальних зооспорових водоростей, у яких є тенденція переходу до багатоядерності; б) лінія одноклітинних та колоніальних автоспорових, у яких простежується тенденція до поділу протопласту у поздовжньому напрямку; в) лінія ценобіальних автоспорових водоростей. Всі ці три лінії в межах порядку за сучасними даними вважаються паралельними еволюційними рядами.
Характерними представниками першої лінії є Neochloris aquatica, Bracteacoccus, Sorastrum, Pediastrum, Hydrodictyon. Всі ці водорості мають голі дводжгутикові зооспори або гамети (п-тип), і показують один з варіантів переходу одноклітинних форм у високоспеціалізовані ценобіальні.
Найпримітивнішим представником є Neochloris aquatica. Ця водорість у дорослому стані одноклітинна. Кожна клітина має парієтальний хлоропласт з піреноїдом та кілька ядер. Розмноження відбувається за допомогою одноядерних дводжгутикових п-зооспор або апланоспор. Зооспори та апланоспори досить часто не одразу звільнюються з оболонки материнської клітини, а починати рости безпосередньо у спорангії. В цьому випадку вони з'єднуються боками, і звільнюються із спорангія у вигляді агрегатів, що нагадують сферичні ценобії. Розпад таких ценобіїв на поодинокі клітини відбувається лише на початку спороутворення, коли кожна клітина стає багатоядерною.
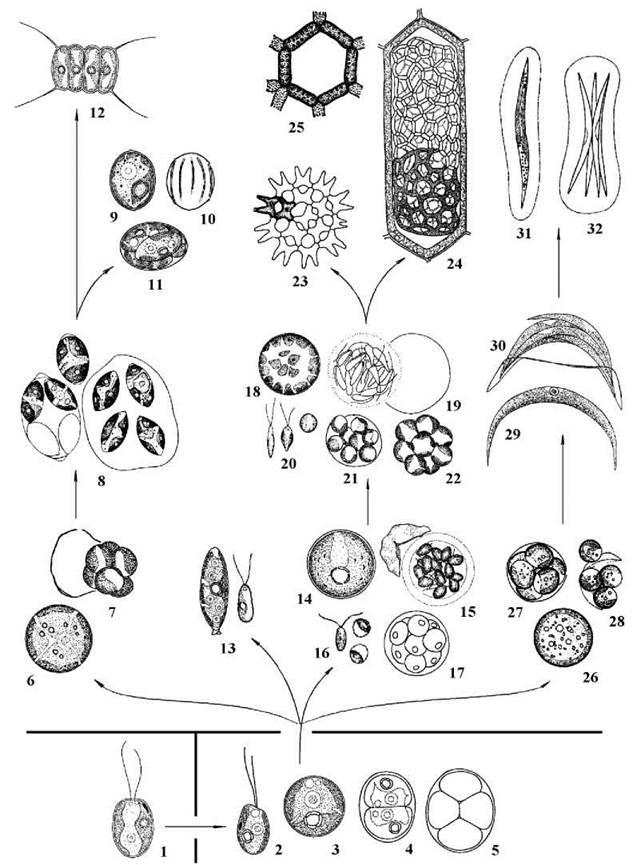
Рис. 24.28. Напрямки еволюції Scenedesmales за молекулярними та цитологічними ознаками. 1 - Heterochlamydomonas (Volvocales), 2-5 - Heterotetracystis(Tetrasporales), 6,7 - Muriella terrestris, 8 - Oocystis, 9-11 - Scotiellopsis, 12 - Scenedesmus, 13 - Characiopodium hindakii, 14-17 - Neochloris aquatica, 18-22 - Bracteacoccus, 23 - Pediastrum, 24, 25 - Hydrodictyon, 26, 28 - Mychonastes homosphaera, 29, 30 - Monoraphidium, 31, 32 - Ankistrodesmus (схема родинних зв’язків - згідно до Friedl, 1997; Lewis, 1997; Sluiman, 1997; та ін. Рисунки - за Oltmanns, 1904; Коршиков, 1953; Starr, 1955; Hindak, 1988; Cox, Deason, 1968, 1969; Fott, Novakova, 1969; Hindak та ін., 1973; Puncocharova, Kalina, 1981; Ettl, Gartner, 1995; та ориг.).
Водорості роду Bracteacoccus у дорослому стані також багатоядерні, здатні утворювати подібні до ценобіїв агрегати, розмножуються зооспорами. На відміну від Neochloris, клітини Bracteacoccusмають багато пристінних дископодібних хлоропластів і позбавлені піреноїдів.
У Pediastrum та Hydrodictyon клітини завжди з'єднуються у характерні для даного роду ценобії. Обидва роди розмножуються зооспорами та мають ізогамний статевий процес.
У видів роду Pediastrum ценобії сплощені, мають вигляд зірки або коліщатка, що складається з 4-128 з'єднаних боками клітин. Кожна доросла клітина містить пристінний хлоропласт з одним-кількома піреноїдами та кілька ядер.
Ценобії Hydrodictyon макроскопічні, і складаються з кількох тисяч великих циліндричних багатоядерних клітин, з'єднаних на кожному полюсі з двома-чотирма сусідніми клітинами. Внаслідок цього утворюються п’яти-шестигранні ячейки, що у сукупності надають ценобію вигляду сітки (звідси походить народна назва Hydrodictyon - водяна сіточка).
При нестатевому розмноженні протопласт кожної клітини водяної сіточки розпадається на кілька тисяч голих дводжгутикових зооспор, що не виходячи за межі материнської клітини з'єднуються між собою і утворюють дочірній ценобій. Після руйнування материнської оболонки такий ценобій починає існувати як самостійний організм.
При статевому розмноженні протопласт клітини розпадається на кілька тисяч голих дводжгутикових ізогамет. Через отвір в оболонці материнської клітини гамети виходять назовні. Далі гамети різних статевих знаків копулюють, і утворюється зигота, що переходить у стан спокою. По його закінченні відбувається мейоз і зигота проростає чотирма великими гаплоїдними зооспорами. Вони здатні відпливати від місця утворення на значну відстань, і таким чином, виконують розсілювальну функцію. По закінченні руху кожна зооспора проростає у багатокутну багатоядерну клітину - т.з. поліедр, що являє собою спорофіт водяної сіточки. Протопласт поліедру ділиться на велику кількість зооспор, що не виходячи за межі поліедру, складаються у новий ценобій. Таким чином, життєвий цикл Hydrodictyon гаплофазний, з зиготичною редукцією та гетероморфною зміною поколінь - ценобіального гаметоспорофіту та одноклітинного спорофіту-поліедру. Аналогічний тип нестатєвого та статевого розмноження має і Pediastrum.
Прикладами водоростей другої лінії сценедесмальних водоростей - лінії одноядерних одноклітинних та колоніальних автоспорових, є роди Mychonastes та Ankistrodesmus.
Клітини Mychonastes поодинокі, дрібні, мають сферичну форму, пристінний хлоропласт без піреноїду, одне ядро. Розмноження відбувається виключно автоспорами, що утворюються по 2-8 і звільнюються при розриві оболонки спорангію. Mychonastes спочатку відносили до роду Chlorella (наприклад Chlorella homosphaera), хоча справжні види Chlorella належать до іншого класу - Trebouxiophyceae. Тільки використання молекулярно- філогенетичних методів дозволило встановити дійсне місце Mychonastes у системі Scenedesmales.
Рід Ankistrodesmus об'єднує колоніальні автоспорові водорості. Колонії утворені двома-чотирма паралельно розташованими клітинами, зануреними у спільний колоніальний слиз. Кожна клітина видовжена та загострена на полюсах, має пристінний хлоропласт, звичайно - з піреноїдом, та одне ядро. При розмноженні протопласт ділиться у поздовжньому напрямку, внаслідок чого утворюється група паралельно розташованих автоспор.
Проміжним родом між двома попередніми є Monoraphidium. Клітини цієї водорості за морфологією та розмноженням подібні до Ankistrodesmus, проте не утворюють колоній, і цим нагадують Mychonastes.
Третю лінію порядку представляють одноядерні автоспорові, переважно - ценобіальні водорості, у яких в оболонці наявний додатковий спорополеніновий шар, а сама оболонка здатна звичайно утворювати різноманітні бородавки, шипи, шипики, щетинки. Характерними представниками є Muriella, Oocystis, Scotiellopsis та Scenedesmus.
Водорості роду Muriella є повним морфологічними "двійниками" роду Bracteacoccus: вони мають сферичні клітини з багатьма пристінними
дископодібними хлоропластами без піреноїдів. Але, на відміну від Bracteacocccus, клітини Muriella одноядерні, не утворюють зооспор, а розмножується лише автоспорами. Досить часто автоспори після звільнення зі спорангію тривалий час лишаються з'єднаними боками, завдяки чому нагадують справжні ценобіальні водорості. Старі клітини накопичують велику кількість вторинних каратиноїдів і забарвлюються у червоний колір.
Для Oocystis характерні еліпсоїдні клітини з невеличкими потовщеннями на полюсах. Водорість вважається колоніальною, оскільки автоспори проростають безпосередньо у спорангії, оболонка материнської клітини при цьому розширюється, проте не руйнується.
Клітини видів роду Scotiellopsis також еліпсоїдні, проте поодинокі. Поверхня оболонки має характерні поздовжні ребра, які добре помітні лише при електронній мікроскопії. Кожна клітина одноядерна, має пристінний перфорований кількома щілинами хлоропласт та один піреноїд. Розмноження відбувається 2-8 автоспорами, що звільнюються при розриві оболонки спорангію.
Для роду Scenedesmus характерні ценобії, утворені звичайно 4 або 8 клітинами, що з'єднані між собою боками. Оболонка клітин має різноманітні "прикраси" у вигляді ребер, шипиків, щетинок, бородавок, довгих рогів. Тип таких "прикрас" є таксономічною ознакою на рівні видів. Розмноження відбувається автоспорами, які ще у материнській клітині з'єднуються у новий ценобій.
Цікаво, що в залежності від хімічного складу води, морфологія "прикрас" може суттєво змінюватись (наприклад, замість рогів утворюватися щетинки або бородавки). Хоча Scenedesmus і вважається типовим прикладом автоспорових хлорофіцієвих водоростей, проте сьогодні у культурі у кількох видів цього роду вдалось отримати також зооспори. Електронно-мікроскопічне дослідження цих зооспор показало, що їх базальні тіла орієнтуються за типом 12-6, тобто так само, як і у Neochloris, Bracteacoccus, Pediastrum, Hydrodictyon.
Сценедесмальні водорості мешкають переважно у планктоні прісних континентальних водойм, і досить часто є домінантами цього типу біоценозів (наприклад, Pediastrum, Monoraphidium, Ankistrodesmus, Scenedesmus). Деякі представники порядку зустрічаються у наземних біотопах (Bracteacoccus, Muriella, Scotiellopsis).
Microsporales. Порядок об’єднує водорості з нитчастим типом структури, що походять від сценедесмальних. Порядок представлений лише одним родом Microspora, що зустрічається у прісних континентальних водоймах.
Систематичними ознаками порядку є наявність голих дводжгутикових зооспор, у яких базальні тіла не зміщені і орієнтовані за схемою 12-6, мікротубулярні корені відповідають формулі 7-1-7-1. З тонкими мікротубулярними коренями пов’язані також додаткові мікрофібрилярні структури (рис. 24.29).
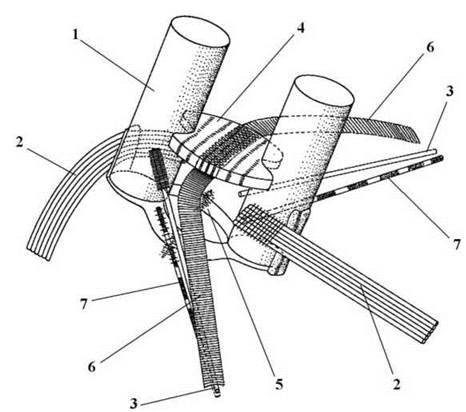
Рис. 24.29. Будова джгутикового апарату у зооспор Microspora: 1 - базальне тіло; 2, 3 - товсті (2) та тонкі (3) мікротубулярні корені; 4, 5 - верхнє (4) та нижнє (5) з’єднувальні волокна; 6, 7 - додаткові мікрофібрилярні корені, асоційовані з тонкими мікротубулярними коренями (за Lokhorst, Star, 1999).
Особливою морфологічною ознакою, що дозволяє досить легко розпізнавати Microspora, є будова оболонки вегетативних клітин. Оболонка складається з двох Н-подібних сегментів, завдяки чому на кінцях ниток завжди наявна характерна "вилка", схожа на аналогічну структуру жовтозелених водоростей з роду Tribonema (рис. 24.30). Кожна клітина містить одне ядро, пристінний сітчастий хлоропласт, який у деяких видів здатний розпадатися на дископодібні сегменти. Піреноїди відсутні. У кількох видів спостерігався ізогамний статевий процес, проте його деталі лишаються не дослідженими.
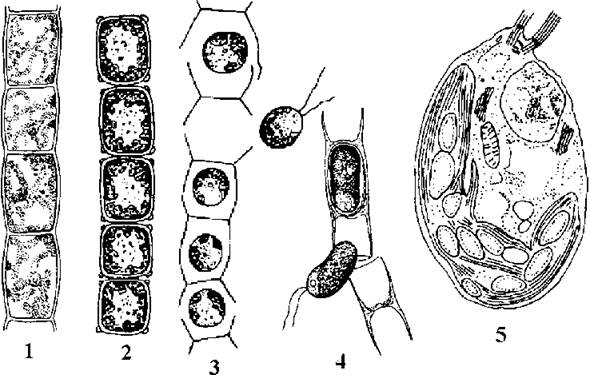
Рис. 24.30. Мікроспоральні водорості: 1-3 Microspora floccosa (1 - вегетативна нитка, 2 - акінети, 3 - вихід зооспор); 4 - клітинні оболонки та зооспори Microspora willeana; 5 - ультратонка будова зооспори Microspora quadrata (1-4 - за Starmach, 1972; 5 - за Lokhorst, Star, 1999).
За цитологічними та молекулярними ознаками Microsporales найбільш близький до зооспороутворюючих сценедесмальних водоростей зі злегка нерівними джгутиками, що нагадують види роду Bracteacoccus.
Sphaeropleales. Порядок включає два роди - Sphaeroplea та Atractomorpha і нараховує біля 10 видів, що зустрічаються у прісних водоймах Африки, Австралії та Південної Америки. Порядок цікавий у теоретичному відношенні, оскільки об’єднує водорості з сифонокладальним типом структури, орієнтацією джгутикових коренів за типом 12-6, "циліндрокапсовим" типом мітозу і голими зооспорами п-типу.
Sphaeroplea має вигляд нерозгалужених ниток, що складаються з морфологічно та функціонально однакових клітин. У кожній клітині міститься сітчастий хлоропласт з піреноїдами, у вільних від хлоропласта ділянках - т.з. лакунах (найчастіше їх чотири у клітині) розташовується два-чотири ядра (рис. 24.31).
Нестатеве розмноження відбувається фрагментацією ниток, зооспорами та особливими клітинами - заростками. Статевий процес оогамний. При споруляції у клітині утворюється кілька великих дводжгутикових макрозооспор. Вони, не виходячи за межі спорангію, проростають у заростки з кількома ядрами. Після цього оболонка спорангію розривається і заростки виходять назовні. З кожного заростку утворюється кілька дводжгутикових мікрозооспор. Останні після короткого періоду руху осідають на субстрат і знов проростають у заросток, що дає початок новій нитці (рис. 24.31).
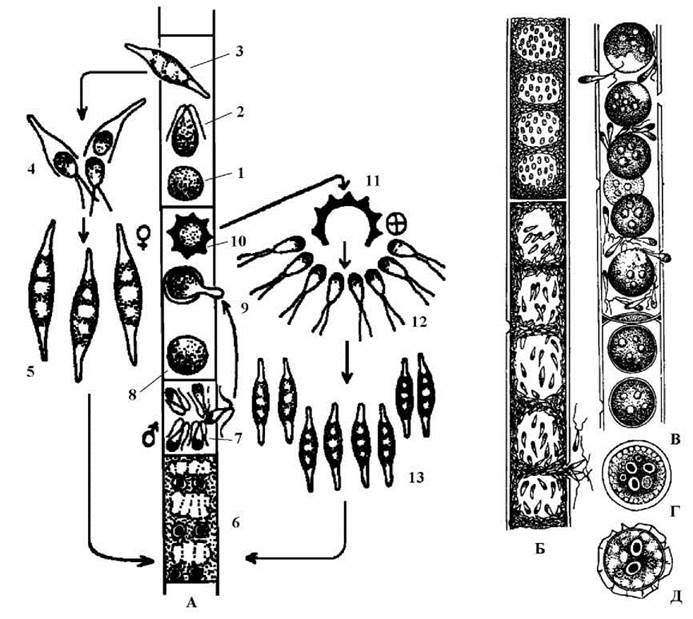
Рис. 24.31. Sphaeroplea. А - цикл розвитку: 1 - протопласт зооспори, 2 - зооспора, 3 - багатоядерний заросток, що утворився із макрозооспори, 4 - проростання заростку мікрозооспорами, 5 - багатоядерний заросток, що утворився із мікрозооспори, 6 - вегетативна багатоядерна клітина, 7 - антеридій із спематозоїдами, 8 - яйцеклітина, 9 - запліднення яйцеклітини сперматозоїдом, 10 - зигота, 11 - проростання зиготи, 12 - зооспори, що утворилися з зиготи, 13 - багатоядерні заростки, що утворилися з зиготичних зооспор; Б - нитка з вегетативною багатоядерною клітиною (зверху) та антеридій з багатьма сперматозоїдами (знизу); В - яйцеклітини та статевий процес; Г - зигота з трьома незаплідненими гаплоїдними ядрами яйцеклітини та одним зиготичним диплоїдним ядром; Д - зріла зигота з шипуватою оболонкою (за Starmach, 1972).
При статевому розмноженні звичайні вегетативні клітини перетворюються на оогонії та антеридії. У Sphaeroplea відомі як однодомні, так і дводомні види. Звичайно в одному оогонії формується 4 яйцеклітини (у відповідності з кількістю лакун), кожна - з чотирма ядрами. В антеридіях розвивається багато дводжгутикових сперматозоїдів. Сперматозоїд запліднює одне з чотирьох ядер яйцеклітини. Три інші ядра дегенерують, після чого утворюється зигота з шипуватою оболонкою. Після періоду спокою в зиготі відбувається редукційний поділ і зигота проростає або зооспорами, які швидко перетворюються на заростки, або безпосередньо заростками. Заростки дають початок новим ниткам.
Поширення та екологія
Хлрофіцієві водорості мешкають переважно в двох типах екотопів: у прісних континентальних водоймах та у ґрунтах, і лише деякі вольвокальні водорості зустрічаються у гіпергалінних озерах.
У водоймах одноклітинні та ценобіальні види представлені переважно планктонними водоростями. Деякі роди зустрічаються майже повсюдно і часто входять до складу домінантів. Це, в першу чергу, Monoraphidium, Ankistrodesmus, Scenedesmus, Pediastrum. В ефемерних водоймах характерними домінантами планктонних угрупувань є Chlamydomonas, Chlorogonium, Pandorina, Eudorina.
У перифітоні Chlorophyceae найчастіше представляють нитчасті та гетеротрихальні водорості з Chaetophorales, Oedogoniales, Microsporales - це роди Uronema, Stigeoclonium, Oedogonium, Microspora, рідше - тетраспоральні та деякі хлорококальні та сценедесмальні - Chaetochloris, Chlamydopodium, Characiopodium. До характерних представників бентосу відносяться Dicranochaete, Bulbochaete, Draparnaldia.
Хлорофіцієві водорості здатні викликати "цвітіння" води та утворювати макроскопічні розростання - жабуріння. Так, "цвітіння" ропи солоних озер звичайно спричинює Dunaliella salina. Вода при цьому набуває червоного забарвлення та фіалкового запаху. Колір обумовлений накопиченням у клітинах великої кількості каротину, а запах - вторинними леткими сполуками, що утворюються при його розкладанні. В ефемерних прісних водоймах червоне "цвітіння" здатний викликати Haematococcus pluvialis. Цікаво, що саме ця водорість найчастіше є забарвлюючим компонентом "кривавих" дощів та "кривавого" снігу. У водоймах, забруднених органічними речовинами, зелені "цвітіння" інколи викликають різноманітні хлорококальні та сценедесмальні водорості.
Жабуріння найчастіше утворюють Oedogonium та Microspora. Обидві водорості проходять початкові стадій розвитку у перифітоні, потім відриваються від субстрату та, продовжуючи розмножуватися, скупчуються у ватоподібних, м’яких та неслизьких на дотик зелених масах на поверхні води.
Рід Nautococcus (Tetrasporales) є нейстонною водорістю. Найчастіше саме вона спричинює утворення зеленої поверхневої блискучої плівки в ефемерних водоймах, особливо - забруднених органічною речовиною.
Численну групу складають ґрунтові хлорфіцієві водорості. У посушливих степових та пустельних ґрунтах масово розвиваються переважно хлорококальні з роду Ettlia та сценедесмальні з роду Scotiellopsis. При старінні вони звичайно накопичують велику кількість каротиноїдів. Так, Ettlia caratinosa є головним збудником червоного "цвітіння" ґрунту посушливих степів та пустель.
У помірно зволожених лісових ґрунтах поширеними хлорофіцієвими водоростями є вольвокальні (Chlamydomonas), хлорококальні (Chlorococcum, Spongiochloris, Tetracystis) та сценедесмальні (Bracteacoccus). Вони зрідка спричиняють зелене "цвітіння" ґрунту. Типовими мешканцями перезволожених ґрунтів є Protosiphon, Pleurastrum, Cylindrocapsa.
Деякі хлорофіцієві, зокрема, Chloromonas nivalis (Volvocales) представляють екологічну групу криофільних водоростей. Цей вид росте на поверхні гірських снігів та льодовиків, і викликає їх зелене та червоне "цвітіння".
У морях та аерофітних біотопах хлорофіцієві водорості майже не зустрічаються.
1 У сучасних визначниках зооспори Pleurastrum insigne наводяться як голі, проте дослідження T.Friedl (1996) показали, що оболонка у зооспор даного виду є, і на електронограмах видно, що вона має електронно-щільний другий шар, чим нагадує оболонки зооспор Chlorococcum oleofaciens.