БИОЛОГИЯ РАЗМНОЖЕНИЯ РАСТЕНИЙ - Н. Л. Колясникова - 2017
РАЗДЕЛ 2. ПОЛОВАЯ РЕПРОДУКЦИЯ РАСТЕНИЙ
Цветок - это репродуктивный орган покрытосеменных растений, совокупность стерильных и фертильных структур. К стерильным относят околоцветник, состоящий из чашечки и венчика. К фертильным - тычинки и плодолистики. Границы между стерильными и фертильными частями условны.
Рассмотрим фертильные части. Тычинка - это преобразованный микроспорофилл. Совокупность тычинок - андроцей. В цветке может быть одна или несколько тычинок. Число их может быть неопределенным или фиксированным. Они могут располагаться на цветоложе по спирали, по кругу или пучками.
Тычинки закладываются в виде бугорков. Сначала развивается ее фертильная часть - пыльник, затем стерильная - тычиночная нить. У некоторых видов (сем. Магнолиевые) часть тычинок полностью стерильны. Они называются стаминодиями. В развитии пыльника различают три периода: предмейотический, мейотический, постмейотический. В первом периоде формируется стенка пыльника, образуется микроспорангий. Во второй период идет дифференциация стенки пыльника и мейоз в микроспороцитах. В третий период происходит созревание пыльцевых зерен.
На ранних этапах пыльник состоит из эпидермиса и меристематических клеток. Далее под эпидермисом выделяются тяжи археспориальных клеток. В результате периклинальных делений этих клеток образуется два слоя: париетальный и спорогенный. Из париетального слоя формируется стенка пыльника, из спорогенного - микроспороциты. В зависимости от направления дифференциации различают 4 типа формирования стенки пыльника [2, 3, 4].
1. Основной тип. Париетальный слой формирует вторичный париетальный слой. Дальнейшие периклинальные деления клеток приводят к образованию эндотеция, нескольких средних слоев и тапетума. Этот тип характерен для древних растений, в частности сем. Винтеровых.
2. Тип двудольных. Париетальный слой клеток путем деления формирует вторичный париетальный слой и тапетум. Далее вторичный париетальный слой дает эндотеций и средний слой.
3. Тип однодольных. Из первичного париетального слоя путем периклинальных делений образуется эндотеций и вторичный париетальный слой. Далее вторичный париетальный слой клеток формирует средний слой и тапетум.
4. Редуцированный тип. Из париетального слоя образуется эндотеций и тапетум.
2.1 Пыльник
На ранних этапах развития в стенке пыльника выделяют четыре слоя клеток: эпидермис, эндотеций, средний слой и тапетум.
Клетки эпидермиса правильной формы, делятся только антиклинального. На поздних этапах покрываются кутикулой, увеличиваются, иногда приобретают зубчатую форму или сплющиваются [9]. В клетках может откладываться крахмал или танин (рис. 6, 7).
Рис. 6. Пыльник козлятника восточного: 1 - поперечный срез пыльника; 2 - стенка молодого пыльника на стадии П I мейоза в микроспороцитах; 3 - стенка зрелого пыльника
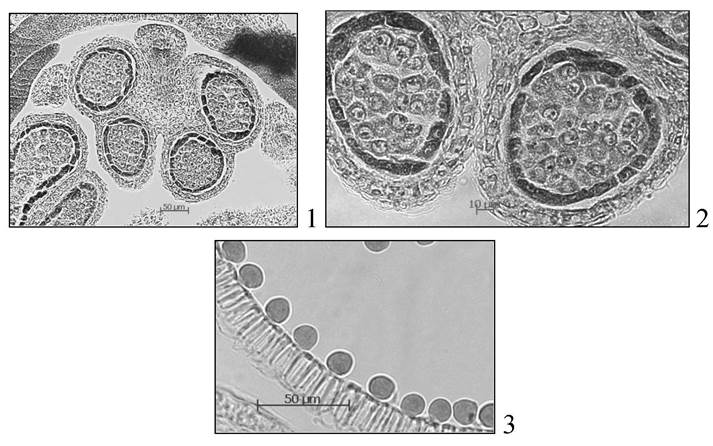
Рис. 7. Развитие стенки пыльника у синяка приятного: 1 - формирование тапетума; 2 - двуядерный тапетум; 3 - дегенерация тапетума, начало развития эндотеция; 4 - стенка зрелого пыльника: т - тапетум; эн - эндотеций; эп - эпидермис
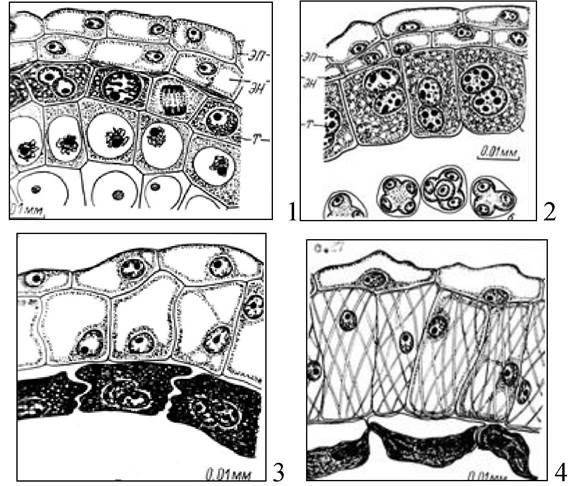
В клетках эндотеция стенки утолщаются, появляются фиброзные тяжи. Эти утолщения способствуют вскрыванию пыльника, когда в гнездах пыльника созрела пыльца.
Количество слоев среднего слоя может быть от 1 до 6. Увеличение происходит за счет дополнительных периклинальных делений. Средние слои постепенно дегенерируют и к моменту созревания пыльцы полностью исчезают.
Тапетум различают наружный (обращен к стенке пыльника) и внутренний (обращен к связнику). Внутренний тапетум формируется за счет деления основной ткани связника. Различают два основных типа тапетума: секреторный и периплазмодиальный. Секреторный сохраняет клеточную структуру до стадии тетрад микроспор. Клетки тапетума могут быть многоядерными, иногда при слиянии ядер полиплоидными. В дальнейшем секреторный тапетум превращается в амебоидный (после стадии тетрад). Часть оболочек клеток растворяются, и цитоплазма впячивается в гнездо пыльника. Периплазмодиальный тапетум образуется до и во время мейоза в микроспороцитах. Цитоплазма заполняет гнездо пыльника, находящиеся в ней ядра делятся синхронно. К моменту созревания пыльцевых зерен тапетум лизируется.
Спорогенные клетки в гнездах пыльника становятся микроспороцитами. Процесс микроспорогенеза у цветковых растений протекает в пыльниках в то время, когда бутон еще не раскрылся. Микроспороциты окружаются специальной каллозной оболочкой. В основе микроспорогенеза лежит мейоз, в результате которого из одной материнской клетки микроспор образуется тетрада микроспор. Различают два типа образования тетрад - одновременное - симультанное и последовательное - сукцессивное. Редко встречается промежуточный тип. Различают тетраэдральные, Т-образные, линейные типы тетрад. В постмейотический период происходит разделение тетрад. Пыльцевое зерно и его ядро увеличиваются в размерах. Появляется вакуоль, ядро смещается к стенке.
Происходит митоз. Образуется линзовидная генеративная клетка. Она перемещается внутрь вегетативной клетки. Вакуоль исчезает, цитоплазма заполняется запасными питательными веществами. Зрелые пыльцевые зерна могут быть двуклеточными и трехклеточными (рис. 8).
Рис. 8. Зрелые пыльцевые зерна: 1 - трехклеточное у подсолнечника; 2 - двуклеточные у люцерны

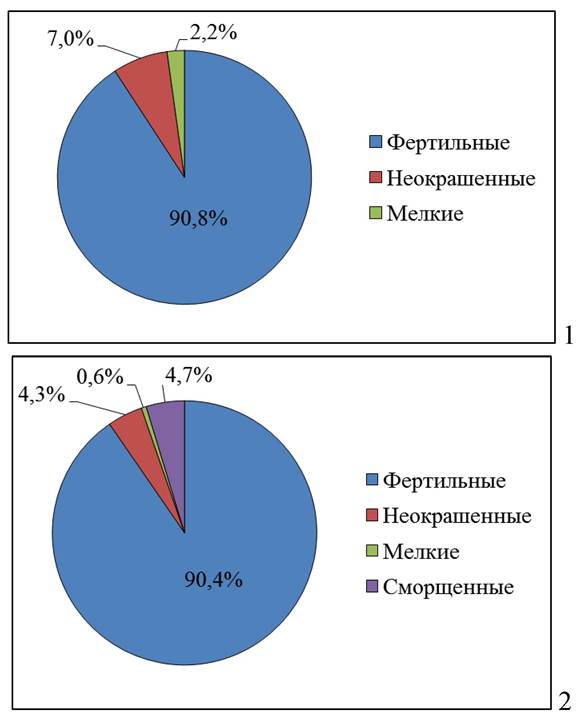
Исследование структурных и функциональных показателей пыльцы растений актуально как с точки зрения теоретической (для понимания механизмов повреждения и процессов адаптации растений в условиях техногенеза), так и практической (для определения потенциальной способности растений из зон загрязнения к формированию полноценного потомства), решения проблем биомониторинга, а также селекции и семеноводства (рис. 9).
Рис. 9. Фертильность пыльцы липы мелколистной в 2016 г.: 1 - берег р. Сылвы; 2 - г. Пермь
Пыльца, способная произвести оплодотворение, называется фертильной. Используют два основных метода: ацетокарминовый и йодный.
Для определения фертильности пыльцевых зерен фиксируют пыльники со зрелой пыльцой в фиксаторе Кларка (3 части этанола: 1 часть ледяной уксусной кислоты). Продолжительность фиксации колеблется от 30 мин до нескольких часов.
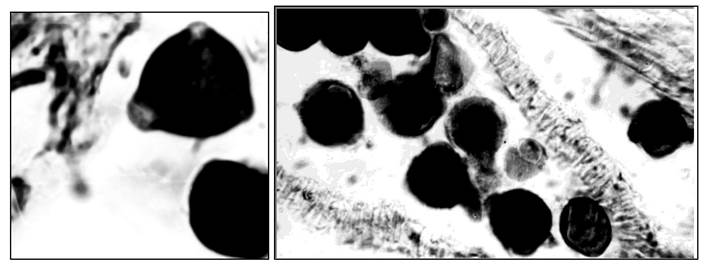
Пыльник переносят на предметное стекло, раздавливают, наносят каплю ацетокармина, препарат накрывают покровным стеклом и осторожно подогревают на спиртовке. У фертильных пыльцевых зерен зернистая цитоплазма и спермии окрашены в густой карминово-красный цвет. Стерильные пыльцевые зерна почти не окрашиваются или окрашиваются неравномерно. Их содержимое часто отходит от оболочки и находится на разных этапах гибели (рис. 10).
Рис. 10. Пыльцевые зерна люцерны посевной: 1 - фертильные; 2 - стерильные
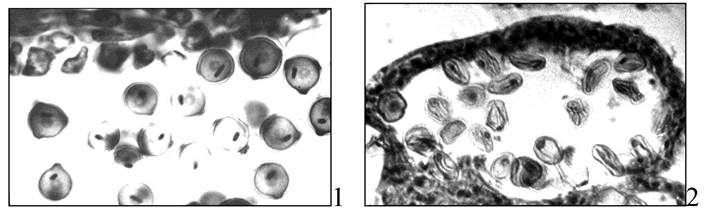
Для некоторых растений, у которых толстая экзина и трудно увидеть спермии при помощи ацетокарминового метода, можно использовать йодный метод. В основе метода лежит определение крахмала при помощи йодной реакции. Фертильные пыльцевые зерна полностью заполнены крахмалом, а стерильные не имеют его совсем или содержат следы (рис. 11).
Рис. 11. Фертильные пыльцевые зерна люцерны
2.2 Микроспорогенез
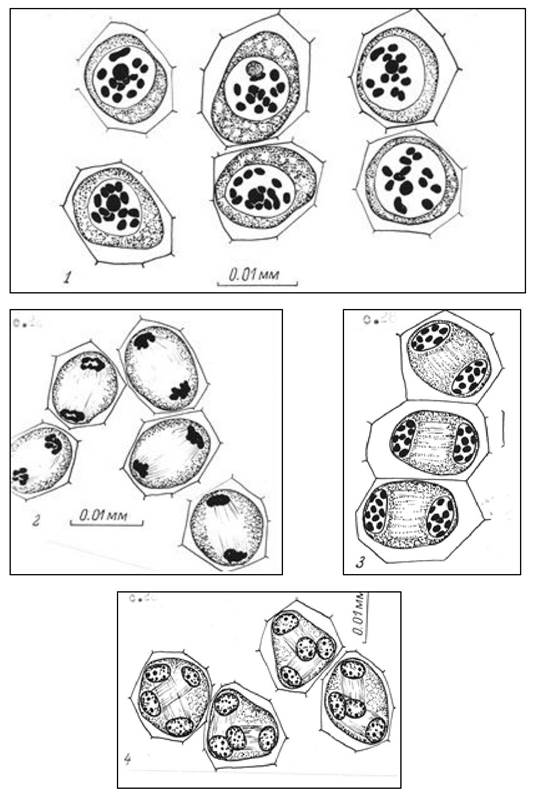
Мейоз - редукционное деление, состоящее из двух делений, следующих друг за другом, приводящее к образованию из одной диплоидной четырех гаплоидных клеток (рис. 12). У высших растений этот особый вид деления протекает до их цветения в молодых пыльниках и семязачатках.
Рис. 12. Мейоз у синяка приятного: 1 - диакинез, 12 бивалентов; 2 - телофаза I; 3 - профаза II; 4 - тетрады ядер
Мейоз протекает в виде двух делений. Наиболее сложной является профаза первого деления мейоза. Начавшийся в интерфазе синтез ДНК продолжается в профазе I.
Профаза включает пять стадий: лептотену, зиготену, пахитену, диплотену и диакинез. На стадии лептотены в ядре появляются тонкие перекрученные нити хромосом. На стадии зиготены происходит конъюгация сначала с концов хромосом, а затем по всей их длине. Конъюгировавшая пара хромосом называется бивалентом. В нем 4 хроматиды, но они еще не различимы микроскопически. На стадии пахитены хроматиды каждой хромосомы уже хорошо видны. Число бивалентов гаплоидно. Конъюгирующие хромосомы могут обмениваться участками хроматид - происходит кроссинговер. В диплотене четко выявляются структуры бивалентов, состоящие из четырех хроматид. Поэтому бивалент называют хромосомной тетрадой. Гомологи отталкиваются друг от друга. В некоторых точках видны места перекреста в виде греческой буквы х - хиазмы. На стадии диакинеза уменьшается число хиазм за счет их передвижения к концам хромосом. Биваленты передвигаются в экваториальную плоскость. Исчезают оболочка ядра и ядрышки.
В метафазе I биваленты выстраиваются в экваториальной плоскости. Нити веретена деления прикреплены к центромерам и тянутся от одного полюса к другому.
В анафазе первого деления мейоза гомологичные хромосомы, состоящие из двух хроматид, отходят к противоположным полюсам клетки.
Телофаза I не отличается от такого митоза. В результате первого деления мейоза образуются два ядра, содержащие гаплоидный набор хромосом, но каждая хромосома имеет две хроматиды.
Второе деление мейоза протекает по типу обычного митоза.
Профаза II не продолжительна, так как хромосомы после телофазы первого деления мейоза остаются спирализованными.
В метафазе II хромосомы располагаются в экваториальной плоскости клетки. Процесс идет, как правило, синхронно в диаде клеток.
В анафазе II к полюсам отходят хроматиды (дочерние хромосомы).
В телофазе II мейоза после цитокинеза одновременно (симультанно) образуются клетки с гаплоидным набором хромосом. Таким образом, в результате двух последовательных делений мейоза из одной диплоидной клетки образуются 4 гаплоидные.
2.3 Семязачаток
Семязачаток состоит из нуцеллуса, интегументов, халазы и семяножки (фуникулюса).
Семязачаток имеет один или два интегумента (покровы). У некоторых растений есть третий - ариллус. Он возникает в результате расщепления наружного интегумента. Также встречается интегумент, сформировавшийся за счет разрастания микропилярной зоны - карункула.
Микропиле может быть образовано либо внутренним, либо внешним интегументом. Иногда оба интегумента участвуют в образовании микропиле.
В зависимости от степени развития нуцеллуса различают крассинуцеллятные и тенуинуцеллятные семязачатки. В первом случае материнские клетки мегаспор отделены от эпидермиса несколькими слоями париетальных клеток. У тенуинуцеллятных материнские клетки непосредственно примыкают к эпидермису. К созреванию зародышевого мешка нуцеллус лизируется.
Обычно одна из клеток нуцеллуса увеличивается в размерах. Цитоплазма становится густой. Это и есть первичная археспориальная клетка. Она может путем деления образовать париетальную и спорогенную клетки или непосредственно превратиться в материнскую клетку мегаспор. Материнская клетка мегаспор делится мейозом, образуя 4 гаплоидные мегаспоры. Расположение тетрады преимущественно линейное, а также Т-образное, крест-накрест, тетраэдрическое. Как правило, из одной из 4-х мегаспор формируется зародышевый мешок.
Типы зародышевых мешков определяются 3 признаками:
1) числом мегаспор, образующих зародышевый мешок;
2) числом митозов;
3) поведением ядер (их распределение).
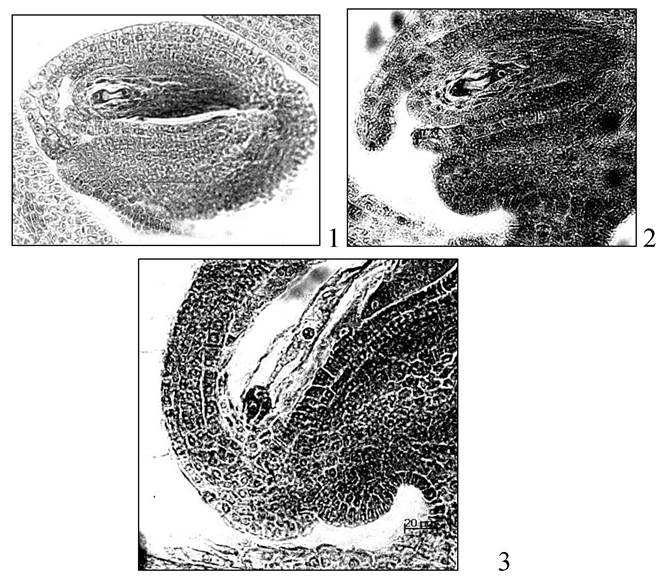
Названия типов зародышевых мешков дают по названию таксонов, где они встречаются. Различают 16 типов. Преобладающий тип - Polygonum (рис. 13).
Рис. 13. Зародышевые мешки козлятника восточного: 1 -двуядерный; 2 - четырехядерный; 3 - зрелый зародышевый мешок
Если в образовании зародышевого мешка участвует одна мегаспора, то зародышевый мешок моноспорический. Если две - биспорический, четыре - тетраспорический.
Примеры разных типов зародышевых мешков:
Моноспорические: Polygonum - тип (развивается из ха- лазальной мегаспоры, трехмитозный) и Oenothera - тип (из микропилярной мегаспоры, двухмитозный).
Биспорический: Allium - тип (из диады мегаспор, хала- зальная делится 3 раза).
Тетраспорические: Fritillaria - тип, Tulipa - тип, Peperonia - тип и др.
Строение зрелого зародышевого мешка.
Яйцевой аппарат состоит из яйцеклетки и двух синергид. Ядро яйцеклетки более крупное. В базальной части яйцеклетки располагается вакуоль. Целлюлозная оболочка только в базальной части. В синергидах расположение ядер обратное. Вакуоли в апикальной, ядро и нитчатый аппарат в базальной части. Нитчатый аппарат способствует проникновению пыльцевой трубки.
Центральная клетка самая большая и сильно вакуолизирована. Она охватывает яйцевой и антиподальный комплексы. В центре клетки два гаплоидных полярных ядра. Центральная клетка в дальнейшем после двойного оплодотворения дает эндосперм.
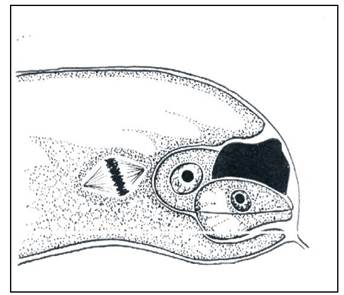
Число антипод зависит от типа зародышевого мешка. В основном - три. Антиподы эфемерны. Есть виды, где число антипод может достигать 300. У Oenothera - типа антиподы отсутствуют. Антиподы выполняют роль гаусториев. У некоторых растений ядра их полиплоидны (рис. 14).
Рис. 14. Зародышевый мешок роговика перед оплодотворением: я - яйцеклетка; с - синергиды; пя - полярные ядра; а - антиподы

2.4 Двойное оплодотворение
Прогамная фаза оплодотворения включает следующие события: прилипание пыльцевых зерен к рыльцу, гидратацию пыльцевых зерен, активацию ферментных систем, узнавание пыльцы, набухание и прорастание пыльцевых зерен, рост пыльцевой трубки, излияние содержимого пыльцевой трубки.
Экзина и интина пыльцевого зерна являются физиологически активными.
Пыльцевые зерна к моменту высыпания из пыльников сильно обезвожены. При попадании на рыльце и прилипании пыльцевого зерна начинается его гидратация - поступление воды из тканей рыльца. Пыльцевое зерно набухает, начинается рост пыльцевой трубки. Все рыльца делятся на два типа: «влажные» с поверхностным секретом и «сухие», без секрета с папиллами (сосочками). Папиллы покрыты пелликулой, которая участвует во взаимодействии рыльца и пыльцевого зерна. Первые - гладкие с поверхности, характерны для растений сем. Бобовые, Пасленовые, Лилейные и др. Вторые - встречаются у растений сем. Астровые, Капустные, Злаковые и др.
Столбики в пестиках также делятся на два типа: открытые (полые) и закрытые. В закрытых столбиках центральную часть занимает проводниковая ткань.
Прорастание пыльцевого зерна начинается с выпячивания интины через проростковую пору. Оболочка пыльцевой трубки состоит из двух слоев: внутреннего (каллозного) и наружного (пектоцеллюлозного). В кончике пыльцевой трубки находится вегетативное и генеративное ядра. По мере роста в пыльцевой трубке образуются каллозные пробки, которые отсекают ее верхушечную часть от остальной (рис. 15).
Рис. 15. Рост пыльцевых трубок в рыльце и столбике разных видов люцерны: 1, 2 - Medicago scutellatа; 3 - Medicago turbinatа

Сначала пыльцевая трубка растет за счет питательных веществ самого пыльцевого зерна, затем питается за счет веществ рыльца, столбика, завязи.
На рыльце гораздо больше растущих пыльцевых зерен, чем число семязачатков. Но скорость роста пыльцевых трубок различна. Некоторые останавливают рост и образуют булавовидные вздутия. В зародышевый мешок входит только одна пыльцевая трубка.
При самонесовместимости пыльца не закрепляется на рыльце, или пыльцевые трубки аномально утолщаются или разветвляются.
Вхождение пыльцевой трубки в зародышевый мешок через микропиле наиболее типичное и называется порогамией (рис. 16). Иногда пыльцевыя трубка входит через халазу или сбоку (халазогамия, мезогамия).
Рис. 16. Порогамия у люцерны посевной

Два спермия из пыльцевой трубки попадают в зародышевый мешок. Один спермий сливается с ядром яйцеклетки, образуя диплоидную зиготу. Второй спермий сливается с полярными ядрами, давая триплоидное ядро эндосперма (рис. 17).
Рис. 17. Метафаза митоза первичного ядра эндосперма и зигота люцерны посевной
2.5 Эндосперм, зародыш
Эндосперм служит источником питания для развивающегося зародыша. По характеру деления первичного ядра эндосперма различают три типа: нуклеарный, целлюлярный и гелобиальный.
Нуклеарный эндосперм. Деление ядер не сопровождается образованием перегородок. Образуется ценоцитная фаза. Иногда могут образовываться выросты - гаустории.
Целлюлярный эндосперм. Деление ядер сопровождается образованием клеточных перегородок. Образуется клеточная фаза. Также могут образовываться гаустории.
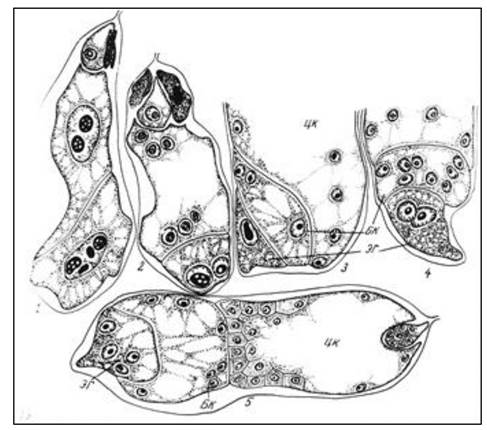
Гелобиальный (промежуточный) эндосперм. Сначала образуется перегородка. Выделяется микропилярная и халазальная части эндосперма. В микропилярной части деление ядер идет по нуклеарному типу с последующим образованием перегородок. В халазальной части - перегородок нет (рис. 18).
Рис. 18. Развитие эндосперма у синяка приятного: 1, 2 - образование перегородки после первого деления первичного ядра эндосперма, боковая и центральная камеры; 3 - образование эндоспермального гаустория; 4 - увеличение числа ядер в боковой камере и эндоспермальном гаустории; 5 - образование клеточного эндосперма в центральной камере: цк - центральная камера; бк - боковая камера; эг - эндоспермальный гаусторий
Независимо от способа образования вполне сформировавшийся эндосперм состоит из довольно крупных клеток, накапливающих запасные вещества в виде крахмала, жиров, алейрона или гемицеллюлозы.
У некоторых растений по мере развития зародышевого мешка нуцеллус не полностью расходуется, а частично сохраняется, превращаясь в запасающую ткань - перисперм, состоящий из диплоидных клеток.
Зигота перед делением проходит фазу созревания или покоя. Первое деление зиготы дает две клетки: апикальную и базальную. Базальная дает суспензор или подвесок. Апикальная - глобулярное тело с радиальной симметрией.
В зависимости от числа делений и направления перегородок различают несколько типов развития зародыша. Рассмотрим некоторые из них.
1. Paeoniad тип. Зигота некоторое время развивается как ценоцит путем свободного деления ядер без образования клеточных перегородок.
2. Piperad -тип. Зигота делится продольной перегородкой.
. Зигота делится поперечной перегородкой.
а) апикальная клетка делится продольно:
- базальная клетка не участвует в построении зародыша - Onagrad -тип.
- апикальная и базальная клетки участвуют в построении зародыша - Asterad - тип.
б) апикальная клетка делится поперечно.
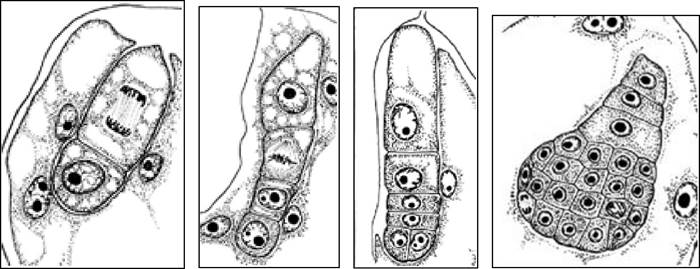
- базальная клетка не участвует в построении зародыша, превращаясь в подвесок - Caryophyllad - тип (рис. 19).
- базальная клетка делится, образуя подвесок - Solanad - тип.
- базальная клетка участвует в построении зародыша - Chenopodiad - тип.
Рис. 19. Развитие зародыша у люцерны (Caryophyllad - тип Medicago - вариация)
В эмбриональный период развития зародыша в апикальной зоне шарообразного тела зародыша появляются точки роста стебля, в базальной - корня и колеоризе. Между ними образуются туника и корпус.
В дальнейшем у растений класса Двудольные формируются два бугорка - семядоли. У растений класса Однодольные образуется один очаг деления клеток, второй блокирован.