ГЕТЕРОФІЛІЯ У РОСЛИН - О.М. НЕДУХА - 2011
РОЗДІЛ ПЕРШИЙ. МОРФОЛОГІЧНА ТА СТРУКТУРНО-ФУНКЦІОНАЛЬНА ХАРАКТЕРИСТИКИ ВЕГЕТАТИВНИХ ОРГАНІВ ГЕТЕРОФІЛЬНИХ РОСЛИН
1.7. ЕПІДЕРМІС ЛИСТКІВ ГЕТЕРОФІЛЬНИХ РОСЛИН
1.7.3. Гідатоди, трихоми й гідропоти
Гідатоди. Для плаваючих листків деяких видів гідрофітів і для листків прикореневої розетки Campanula rotundifolia характерна наявність клітинних структур - гідатод, які спеціалізуються на виділенні води та водних розчинів органічних і неорганічних речовин (Takeda et al., 1991; Pederson, Sand-Jensen, 1993; Барабанов, 2006). Гідатоди складаються з епітеми - безхлорофільних епідермальних клітин. Детально описана структура гідатоди в молодих листках Sparganium emersum (L.) Rehman: листкові гідатоди являють собою канал, утворений між двома сусідніми епідермальними клітинами. В порожнину гідатоди повернуті клітини ксилеми провідних пучків і паренхімні клітини; тимчасом як зовні - канал оточений лігніфікованими клітинними оболонками епідермальних клітин (Chen, Chen, 2005). Пектоцелюлозні оболонки двох епідермальних клітин закривають отвір гідатоди. Цей отвір може бути відкритим тривалий час, і тоді водорості та бактерії, попадаючи всередину гідатоди, утворюють «корок» біля відкритого ксилемного елемента, через який надходить вода. Подібний корок може бути утворений також з кутину, суберину чи калози (Pederson et al., 1997). Автори дійшли висновку, що мінімальний кореневий тиск, який спричиняв потік води через гідатоди для Sparganium emersum становив 2,13 мл -лист-1 год. -1, при наявності кореневого тиску близько 3 кПа. Водночас, у гідрофіта Myriophyllum sp. кореневий тиск був на порядок вищий і становив 30 кПа (Pederson, 1994). Встановлено, що гідравлічна провідність (через гідатоди) на апексах молодих листків була вищою, ніж у більш старих листків, що свідчило про онтогенетичну залежність гідравлічної провідності листків досліджуваного гідрофіта. Гутацію в молодих листках Fragaria L. (Takeda et al., 1991) і Lobelia L. (Pederson, Sand-Jensen, 1993) було описано раніше.
Трихоми. На поверхні плаваючих листків Trapa natans (Bercu, 2004) і Victoria amazonica Sowerby (Carpenter, 2006), та повітряних листків Marsilea quadrifolia (Lin et al., 2007; Tai-Chung Wu, 2010) відмічено наявність трихом, які за структурою подібні до трихом листків суходільних рослин (Werker, 2000). Трихома - це одно чи кілька клітинна структура на поверхні епідермісу листків (рис. 1.7.3.1, г), крізь які здійснюються транспорт назовні різних метаболітів і водних розчинів солей. Трихоми захищають надводні листки вищих водних рослин і листки водної папороті від перегріву та надмірної інсоляції. Щільність трихом різна на верхній і нижній поверхнях листків (Tai-Chung Wu, 2010). Прості трихоми, як відомо, можуть утворюватися ще на початку розвитку листкового примордію, коли про- тодермальні клітини стають трихомами, а оточуючі епідермальні клітини примордію продовжують звичайний поділ (Schnittger, Hulskamp, 2002).
Трихоми на певних фазах розвитку листка чи стебла можуть злущуватися або залишатися, доки рослина не постаріє. Мертві трихоми можуть продовжувати функціонувати для поглинання води чи абразивного захисту листкової пластинки (Werker, 2000). Крім того, відомо, що прості трихоми можуть виводити на поверхню епідермальних клітин хімічні елементи, включаючи Ca, Cd, Zn, Mn, Ni, Pb, S, Si та ін. (Uphof, 1962; Salt et al., 1995; Kupper et al., 2000; Choi et al., 2001). Солі, які виходять крізь трихоми, часом формують кристали на їхній поверхні (Choi et al., 2001).
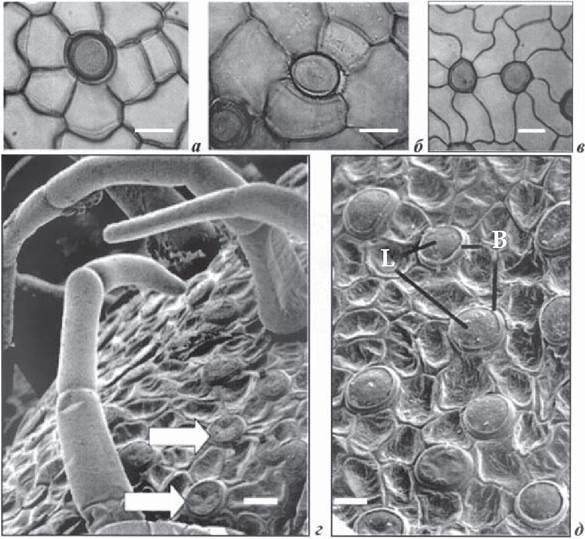
Рис. 1.7.3.1. Фрагменти клітин нижньої поверхні листків гідрофітів з гідропотами (а, б, в- світлова мікроскопія, після видалення кутикули, г, д - скануюча електронна мікроскопія: а - гідропота стефаноцитного типу, Numphar advena (Aiton) W.T. Aiton, б - комплекс гідропоти з неправильною архітектурою, N. polysepala; в - комплекс двох актиноцитних гідропот, N. caerulea Savigny; г - нижня поверхня листка Victoria amazonica з численними гідропотами без волосоподібних верхніх частин (стрілки) і кілька гідропoт з прикріпленими волосоподібними структурами; д - базальна частина (В) і лінзоподібна клітина (L) комплексу гідропот V. amazonica (Carpenter, 2006). Реперна мітка = 20 мкм
Трихоми також беруть участь у регуляції виходу апопластного кальцію на поверхню органу (DeSilva et al., 2001), регулюючи роботу продихів. Крізь трихоми на поверхню листка виходять і токсичні компоненти. Фенольний ексудат трихом забезпечує першу лінію захисту листків від дії мікроорганізмів (Wagner et al., 2004). Дослідники вважають, що трихоми можуть також виводити на поверхню олігосахарини з апопласту з метою захисту листків від інфікування. Недавно описано новий дитерпеноїд (лабдіен-диол), виділений з трихом тютюну при вірусному інфікуванні. Цей дитерпеноїд активував протеїнкіназу та збільшував резистентність до вірусного інфікування. Автори припускають, що лабдіен-диол є сигнальною молекулою під час дії патогену на рослинну клітину (Seo et al., 2003).
Гідропоти. Для плаваючих і підводних листків, стебел і пелюсток квітів гетерофільних водних рослин характерна наявність особливих структур - гідропот, що представляють собою певний тип залоз, і мають дві функції: секреторну та абсорбційну (Luttge, Kraft, 1969; Wilkinson, 1979). Гідропоти беруть участь у транспорті іонів і води як на близькі віддалі (водне оточення ↔ клітини епідерми), середні (водне оточення ^ фотосинтезуючі клітини листка), так і на далекі віддалі (водне оточення ↔ листок ↔ черешок ↔ корінь) (Luttge, Kraft, 1983). Гідропота це структура на поверхні епідерми листків і стебел з незвичайними лабіринтами клітинних оболонок. Гідропоти мають чашеподібну форму, ці структури підняті над епідермою. Досліджуючи методом світлової й скануючої електронної мікроскопії нижню й верхню поверхні плаваючих листків шести видів Nuphar advena, N. lutea, Numphaea polysepala (Engelm.) Greene., Numphaea caerulea, Numphaea flava Aether ex Audubon і Numphaea nouchali Burm f. , а також листків Victoria amazonica (рис. 1.7.3.1) Дж. Карпентер (Carpenter, 2006) встановив, що гідропоти - це одна клітина або ж група клітин сте- фаноцитного чи актиноцитного типу, що мають у своїй основі субепідер- мальну клітину двояковипуклої лінзоподібної форми. За термінологією У. Лютге та ін. (Luttge, Krapf, 1969; Wilkinson, 1979) ця клітина має назву клітина-підошва («foot» клітина) (рис. 1.7.3.1), верхня клітина - це основна клітина гідропотного комплексу чашоподібної форми, має заглибину й товсті клітинні оболонки. Крім того, деякі автори відмічають, що гідропоти можуть виявлятися на обох поверхнях листків у широкого кола водних покритонасінних і водних папоротей (Kaul, 1976; Wilkinson, 1979; Carpenter, 2006). Розмір і щільність гідропот в епідермі залежить від виду й типу епідерми. Р. Каул, вивчаючи гідатоди в гетерофільних і гомофільних видів із різних родів, припустив, що гідропота, як структура епідерми, є типовим прикладом конвергентної морфології водних рослин (Kaul, 1976).