БОТАНІКА 3 ОСНОВАМИ ГІДРОБОТАНІКИ - 2010
1. ОСНОВИ ВЧЕННЯ ПРО КЛІТИНУ (ЦИТОЛОГІЯ)
1.1. Форми життя на Землі
Клітина — це основна структурно-функціональна одиниця всіх живих організмів, елементарна жива система.
У біосфері існує близько 5 млрд. різних організмів. Більшість із них мають клітинну будову. Лише віруси являють собою неклітинні форми життя. Віруси складаються лише із молекули нуклеїнової кислоти (ДНК або РНК) й білкової оболонки. Вони є найпростішою формою життя - доклітинною.
У більшості живих організмів тіло складається з однієї (одноклітинні) або багатьох (багатоклітинні) клітин. На підставі особливостей будови клітин клітинні організми поділено на дві групи: прокаріоти та еукаріоти. Виявлено і третю форму - мезокаріоти - організми з проміжним типом організації генетичного апарату.
Прокаріот означає доядерний. Такі живі організми не мають чітко сформованого ядра, і генетичний матеріал у них міститься у вигляді молекули ДНК безпосередньо в протоплазмі клітини, не захищений ядерною оболонкою. До прокаріотів відносяться бактерії і синьо-зелені водорості.
Еукаріот у перекладі з грецької означає: той, що має справжнє ядро. Клітини еукаріотів містять справжнє ядро, відокремлене від цитоплазми подвійною мембраною. До еукаріотів належать усі тварини й вищі рослини, одно- і багатоклітинні водорості, гриби.
Клітина є одиницею будови і життєдіяльності всіх живих організмів. У ній зосереджені всі прояви життя. Вона засвоює речовини й енергію з довкілля, дихає, відповідає на подразнення, росте, розвивається, розмножується шляхом поділу.
Одноклітинні організми виконують усі властиві їм функції. Характерною рисою одноклітинних структур є одноклітинна будова і чітко відособлене ядро та інші органоїди. До одноклітинних організмів відносяться діатомові, зелені, евгленові та інші водорості.
У колоніальних структур організм складається із численних клітин, об'єднаних загальною слизовою масою. При цьому клітини зберігають самостійність і не з'єднані між собою обміном речовин. Але серед них виділяються окремі групи клітин (вольвокс), які об’єднані цитоплазматично і функціонально. Такі організми займають проміжне положення між типовими колоніальними та багатоклітинними структурами, властивими вищим рослинам.
У багатоклітинних організмів групи клітин пристосувалися до виконання будь-якої однієї або декількох функцій, але виконують їх повніше та досконаліше. Клітини багатоклітинного організму пов'язані між собою в єдину систему. Багатоклітинні структури властиві організмам, що досягли високого рівня розвитку. Серед них є такі, що не мають розчленування тіла на справжні тканини і вегетативні органи. Це харові, бурі та червоні водорості. А вищі спорові та квіткові рослини вже диференційовані на справжні тканини, вегетативні та генеративні органи.
1.2. Клітинна теорія будови організмів
Структурною та функціональною одиницею рослинного організму, як і інших живих істот, є клітина. Вперше цей термін запропонував у 1665 р. англійський вчений Роберт Гук.
Німецький ботанік М. Шлейден (1838) довів, що клітина та її органоїди є складовими рослинного організму. Наступного року його співвітчизник зоолог Т. Шванн (1839) вперше в біології чітко і повно сформулював клітинну теорію. Ось її основні постулати:
- клітина — основна структурна і функціональна одиниця живих організмів;
- клітини тварин і рослин мають спільні принципи будови;
- клітина виникає від клітини;
- живий організм — це сукупність клітин, що зв'язані між собою різноманітними формами регуляції.
Клітинна теорія будови організмів мала великий вплив на розвиток не тільки ботаніки, а й всього природознавства. Вона спрямовувала зусилля вчених біологів на детальніше вивчення вмісту клітини, довела єдність походження органічного світу. Таким чином, клітинна теорія стала основою еволюційної теорії рослинного світу.
1.3. Основні відмінності рослинної клітини від тваринної
Рослинні й тваринні клітини подібні між собою. Однак, незважаючи на спільність основних структур, вони мають ряд істотних відмінностей, пов'язаних з особливостями будови, життєдіяльності та способу життя рослин і тварин.
Основні відмінності рослинної клітини від тваринної полягають
у наступному:
- у рослинних клітинах присутні особливі органоїди — пластиди, яких немає у тварин;
- рослинні клітини завжди оточені міцною твердою оболонкою, що захищає їх від несприятливих умов навколишнього середовища, надає їм форми і міцності;
- рослинні клітини характеризуються розвиненою системою вакуолей з клітинним соком, які забезпечують осмотичні властивості клітин — тургор і надходження до них води;
- рослинні клітини пов'язані між собою цитоплазматичними тяжами — плазмодесмами, що проходять крізь тверду клітинну оболонку і з'єднують клітини організму в єдину систему;
- у багатьох рослинних клітинах у різних формах відкладаються запасні поживні речовини, цьому сприяє перевага в автотрофних рослин процесів синтезу над розпадом.
У рослинному організмі, як правило, присутні й мертві клітини. Особливо багато їх (до 80 %) у стеблах багаторічних рослин. У цьому випадку клітиною називають оболонку, яка позбавлена живого вмісту, але виконує в організмі певні функції (опорну, провідну).
1.4. Форма і розміри рослинних клітин
Рослинні клітини мають різноманітну форму (округлу, кубічну, призматичну, овальну, веретеноподібну, зірчасту тощо). Це залежить від середовища, яке їх оточує, та функції клітини.
Залежно від форми рослинні клітини поділяють на два типи: па-ренхімні та прозенхімні.
Паренхімні клітини мають більш-менш рівні величини всіх трьох вимірів (довжина, ширина і висота). Найбільший діаметр не більше ніж у 2—3 рази перевищує найменший. Паренхімними є клітини твірної тканини і деяких постійних — шкірки, запасаючих тканин, серцевини стебла, основної паренхіми тощо. Тканини, що складаються з паренхімних клітин, як правило, живі.
Прозенхімні клітини витягнуті у довжину і часто мають загострені кінці. Довжина прозенхімних клітин перевищує їх ширину в 5—20 і більше разів. Прозенхімні клітини частіше мертві, позбавлені живого вмісту. З них складаються провідні та механічні тканини.
Форма і будова клітин залежать від місця їх розміщення та функції. Клітини конуса наростання ізодіаметричні, але, спеціалізуючись на виконанні фізіологічних функцій, вони змінюють свою форму і будову. Клітини зовнішнього шару, що дають початок епідермісу, призматичні або плоскі. Клітини центральної частини конуса наростання багатокутні, видовжені, дають початок провідним, основним та іншим тканинам. Клітини епідермісу плоскі із звивистими оболонками; клітини, які проводять речовини, трубчасті з тонкостінною целюлозною оболонкою; клітини, що виконують механічну функцію, мають потовщені клітинні оболонки.
Розміри рослинних клітин дуже малі: їх можна побачити тільки під мікроскопом. Середній діаметр їх коливається від 10 до 100 мкм. Клітини твірної тканини звичайно дрібніші — 5-8 мкм, а клітини запасаючих тканин серцевини стебла й м'якоті соковитих плодів (яблуко, кавун, цитрусові), навпаки, великі — 0,2-1 мм, їх можна бачити неозброєним оком. Значну довжину мають прозенхімні клітини — волокна. Так, волокна льону в довжину досягають 4 см, бавовнику — 5 см, кропиви звичайної — 8 см, рамі — 22 см. Однак у поперечнику вони мають звичайні клітинні розміри — 50-100 мкм, тому їх не можна побачити неозброєним оком. Найбільшу довжину, до декількох метрів, мають молочники деревних рослин.
1.5. Будова рослинної клітини
Доросла рослинна клітина складається з трьох частин: щільної еластичної оболонки, що оточує клітину зовні; протопласта — живого вмісту клітини, притиснутого у вигляді тонкого шару до клітинної оболонки; вакуолі — порожнини, що займає центральну частину клітини і заповнена водянистим клітинним соком.
Протопласт, або живий вміст клітини, — її основна, найважливіша частина, що складається із сукупності клітинних органоїдів (ядро, пластиди, мітохондрії, лізосоми, рибосоми та ін.).
Клітинна оболонка і вакуолі є продуктами життєдіяльності протопласта, його похідними і з'являються на певному етапі розвитку клітини.
Протопласт поділяють на два компоненти: ядро і цитоплазму, яка, в свою чергу, складається з корпускулярних і мембранних органоїдів, занурених в основну речовину — матрикс, або цитозоль (рис. 1.1).
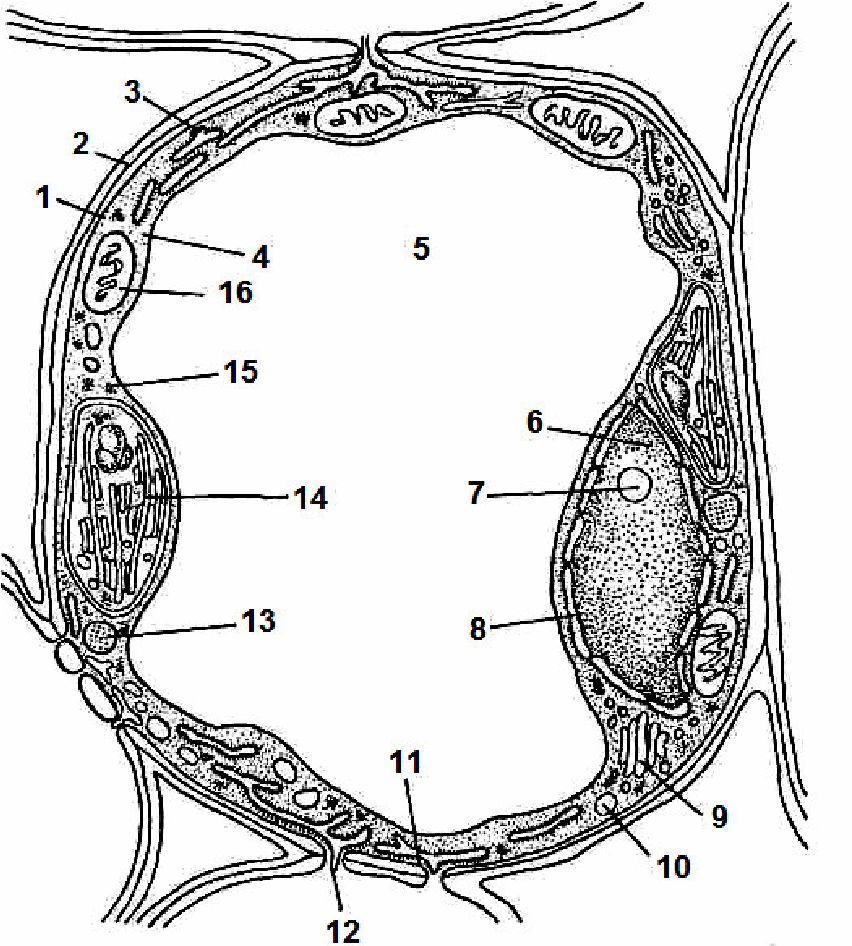
Рис. 1.1. Схема будови клітини мезофілу листка
1— клітинна оболонка; 2 — серединна пластинка; 3 — плазмалема; 4 — цитоплазма; 5 — вакуоля; 6 — ядро; 7— ядерце; 8 — ядерна оболонка; 9 — апарат Гольджі; 10 — ліпідна краплина; 11 — ендоплазматичний ретикулум; 12 — плазмо десма; 13 — мікротільця; 14 — хлоропласт; 15 — рибосома; 16 — мітохондрія
Протопласт і цитоплазма. Протопласт — живий вміст клітини, він безбарвний і прозорий. В'язкість протопласта в клітинах різних рослин неоднакова. У середньому вона перевищує в'язкість води у 12-20 разів, у водних рослин — лише в 5-6.
Частина протопласта рослинної клітини, за винятком ядра, має назву цитоплазма. В цитоплазмі можна виділити три шари: плазма-лему, мезоплазму і тонопласт.
Зовнішній шар, що прилягає до клітинної оболонки, — плазмалема — дуже тонкий, містить багато ліпідів, міцели в ньому розташовані впорядковано і точно орієнтовані. Органоїдів у цьому шарі немає. Середній шар — мезоплазма — товстіший, міцели тут розташовані безладно. Він містить усі клітинні органоїди. Третій, внутрішній, шар — тонопласт — оточує вакуолю. Він подібний до пла-
змалеми.
Мембрани. Протопласт зовні та зсередини обмежений відповідними мембранами: плазмалема відокремлює його від клітинної оболонки, а тонопласт — від вакуолі. Розрізняють також мембрани ядра, мітохондрій, пластид, субодиниць апарату Гольджі, а також внутрішні мембрани цитоплазми — ендоплазматичного ретикулуму, мітохондрій і хлоропластів. Мембрани — високоорганізовані структури клітин, склад яких залежить від типу і функції мембрани, але завжди в них є ліпіди та білки.
У 1972 р. англійські вчені С. Сінгер і Г. Ніколсон запропонували рідинно-мозаїчну модель мембрани, за якою молекули білків у ліпідах утворюють щось подібне до мозаїки (рис. 1.2).
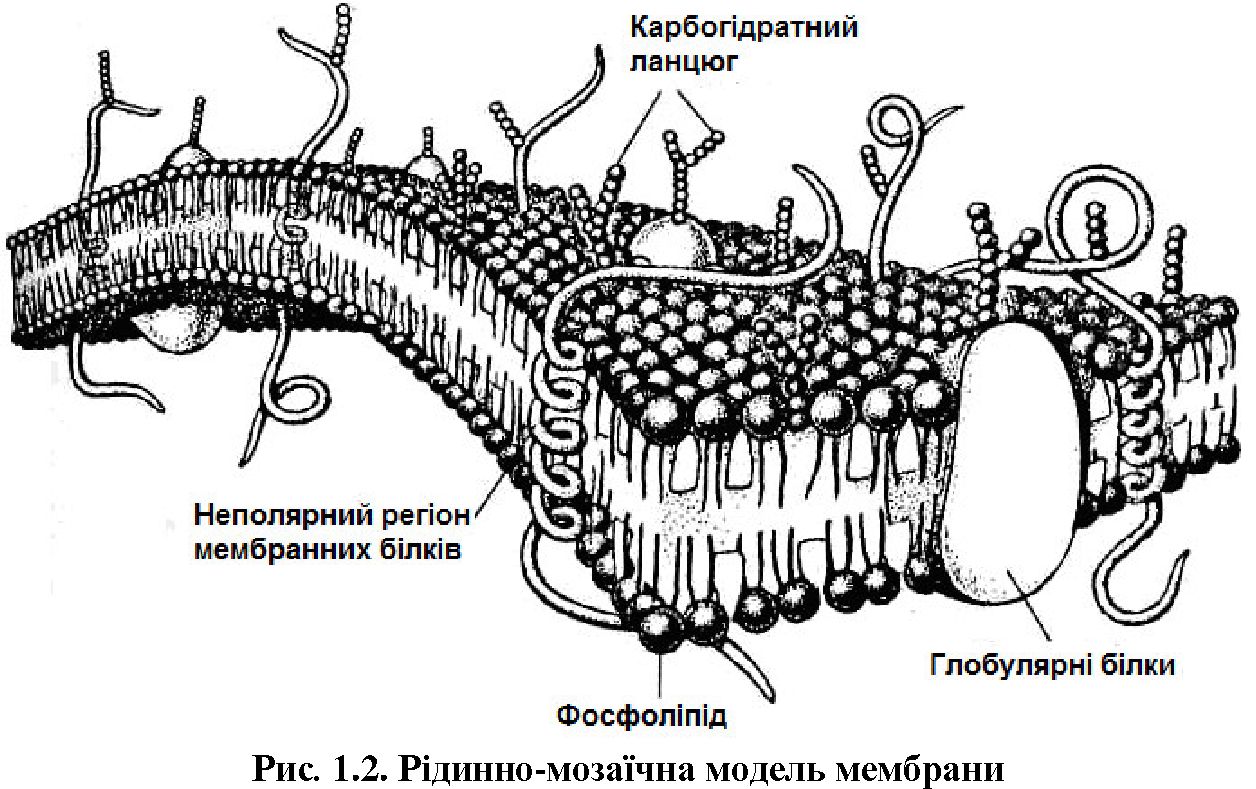
Ліпіди в мембранах представлені фосфоліпідами, гліколіпідами та стеролами. Ймовірно, ліпідний шар пронизують білкові молекули, які можуть бути зв'язані своїми гідрофобними бічними ланцюгами з внутрішніми гідрофобними частинами молекул ліпідів. Гранули внутрішніх ділянок мембран найчастіше є білками, які гідрофобно закріплені в ліпідному матриксі мембрани.
У клітинних мембранах трапляються тисячі різних білків. Деякі з них лише частково занурені в мембрану, інші пронизують її повністю. Гідрофобні ділянки білків взаємодіють з ліпідами, тоді як гідрофільні контактують з водним вмістом клітини. Мембранні ліпіди створюють середовище, потрібне для функціонування цих білків.
Лабільна структура мембран дає їм змогу, в свою чергу, виконувати найрізноманітніші функції: бар'єрну, транспортну, осмотичну, електричну, структурну, енергетичну, біосинтетичну, секреторну, рецепторно-регуляторну тощо. Кожен органоїд теж має власні функції, що здійснюються в унікальному внутрішньому середовищі. Створюється це середовище завдяки вибірковій проникності та іншим специфічним властивостям мембран, що оточують органоїд та відокремлюють її від решти компартментів протопласта.
Матрикс цитоплазми, або цитозоль. Матрикс — це основна речовина цитоплазми, його ще називають цитозоль, гіалоплазма (від грец. «гіалос» — скло). Він являє собою колоїдну систему з великою кількістю води (до 90 %). У матриксі міститься велика кількість білків-ферментів. У нього занурені всі клітинні органоїди, включаючи ядра.
Функції матриксу — об'єднання і взаємозв'язок усіх органоїдів, транспорт речовин між ними, здійснення численних ферментативних процесів (гліколіз, синтез ліпідів, початкові етапи синтезу білка тощо). Матрикси всіх клітин взаємопов'язані через плазмодесми.
Клітинні органоїди. Органоїди — це структуровані елементи цитоплазми, що виконують у клітинах певні функції, пов'язані з їх життєдіяльністю.
Корпускулярні органоїди мають цілком визначену форму (округлу, овальну, дископодібну, паличкоподібну, ниткоподібну тощо). До корпускулярних органоїдів належать ядро, пластиди, мітохонд-рії, лізосоми, рибосоми, мікротільця.
Мембранні органоїди складаються із сукупності мембран і не мають точно обмеженої форми (ЕПС, плазмалема, тонопласт). Апарат Гольджі — органоїд, що поєднує в собі ознаки як корпускулярної, так і мембранної структури.
Серед перерахованих клітинних органоїдів більшість є спільними для рослин і тварин, але є і спеціальні, притаманні тільки рослинним організмам. До останніх належать пластиди і деякі мікротільця (пероксисоми, гліоксисоми). Продукти життєдіяльності протопласта — теж суто рослинні компоненти клітини (клітинна оболонка, вакуолі, відкладення запасних поживних речовин).
Мембранні органоїди. До таких органоїдів належать: плазмалема, тонопласт, ендоплазматична сітка.
Мембрани рослинних клітин, як і всі природні ліпопротеїнові мембрани, складаються з подвійного шару ліпідів, у який занурені білкові глобули.
Плазмалема — це плазматична мембрана, що оточує зовні протопласт і прилягає до клітинної оболонки. Плазмалема може утворювати впинання (інвагінації). Основні функції плазмалеми:
1) обмін речовин між клітиною і довкіллям;
2) синтез целюлози;
3) участь в осмотичних властивостях клітини;
4) сприйняття подразнення;
5) зв'язок між клітинами: плазматичні мембрани всіх живих клітин рослини пов'язані між собою за допомогою плазмодесм.
Тонопласт — мембрана, що оточує вакуолю і відокремлює її від цитоплазми. Тонопласт за будовою багато в чому подібний до плазмалеми: має приблизно ту ж товщину, механізми транспорту речовин — пасивні й активні, а також може зливатися з пухирцями, які оточені мембранами і вміщують речовини, що надходять у вакуолю (за типом екзоцитозу).
Ендоплазматична сітка, або ендоплазматичний ретикулум (reticulum — сітка) — це складна тривимірна мембранна система, яка являє собою систему канальців, пухирців і цистерн, обмежених ліпопротеїновими мембранами.
Розрізняють гранулярну і гладку ендоплазматичну сітку (ЕПС).
На поверхні гранулярної сітки міститься велика кількість гранул-рибосом. Гранулярна ЕПС притаманна клітинам, у яких відбувається інтенсивний синтез білка. її функції пов'язані із транспортуванням білків, синтезованих рибосомами. У процесі перенесення білок зазнає істотних змін, наприклад, фосфорилюється, ацетилюється, перетворюється на глікопротеїни тощо.
Клітини, де відбувається інтенсивний синтез ліпідів, мають іншу досить розгалужену гладеньку трубчасту ЕПС. Вона не несе рибосом на поверхні своїх мембран. Частіше складається з трубчастих видовжених канальців. На гладких мембранах в основному синтезуються ліпіди і вуглеводи. Обидва типи ЕПС можуть бути одночасно навіть в одній клітині, причому між ними існує взаємозв'язок.
Розглянемо функції ЕПС у рослинних клітинах.
Компартменталізація. ЕПС поділяє клітину на ділянки — ком-партменти. Компартменти — це реакційний простір, оточений мембранами. Це дуже важливо для протікання різних реакцій і процесів
метаболізму в окремих ділянках клітини. ЕПС утворює всередині клітини велику поверхню для протікання різних реакцій і процесів.
Синтетична функція. На мембранах ЕПС протікають синтетичні процеси: на гранулярних мембранах відбувається синтез білка, на гладких — вуглеводів і ліпідів.
По канальцях ЕПС відбувається внутрішньоклітинний транспорт речовин (білків, ліпідів, вуглеводів тощо).
ЕПС бере участь в утворенні мембран деяких органоїдів (апарат Еольджі, сферосоми тощо).
ЕПС всіх живих клітин рослинного організму пов'язані в єдину систему за допомогою плазмодесм.
Плазмодесми — це цитоплазматичні тяжі, що з'єднують дві сусідні клітини. Цитоплазматичні тяжі проходять через найтонші плаз-модесменні канальці, які пронизують клітинні оболонки двох сусідніх клітин. У центрі канальця знаходиться десмотрубочка, пов'язана з канальцями ЕПС сусідніх клітин і з'єднує їх. Таким чином, плазмодесми поєднують усі живі клітини рослини в єдину живу систему — симпласт. Вона сприяє пересуванню речовин з клітини в клітину. У рослинах можна виділити ще одну систему — апопласт. Це мертва система, яка поєднує клітинні оболонки, міжклітинники і порожнини мертвих клітин. Апопласт знаходиться назовні від пла-змалеми і теж використовується рослиною для транспорту речовин.
Апарат Гольджі. Серед структур цитоплазми мембранної природи велике значення має апарат, або комплекс Еольджі — багатоярусна система плоских порожнистих дископодібних цистерн (диктіосом), які до периферії потовщуються і утворюють трубчасті відростки. Диктіосома (тільце Еольджі) — це купка плоских цистерн із гладких мембран діаметром 1-2 мкм. У диктіосомах найчастіше буває 4-8 цистерн, які не стикаються одна з одною, а розташовуються на відстані приблизно 10 нм.
У рослинних клітинах звичайно присутні в середньому від 10 до 50 диктіосом, безладно розсіяних у цитоплазмі. їх кількість залежить від типу клітини та її активності. У клітинах деяких водоростей міститься лише одна диктіосома. Сукупність усіх диктіосом клітини складає її апарат Еольджі.
Диктіосома звичайно розташовується таким чином, що її зовнішня цистерна паралельна канальцю ЕПС. Між ними знаходиться багато дрібних пухирців, які називаються перехідними. Припускають, що перехідні пухирці відокремлюються від мембран ЕПС і, зливаючись, утворюють цистерну диктіосоми. Цей бік диктіосоми називають таким, що формується. З її протилежного боку остання цистерна розпадається на дрібні пухирці — пухирці Гольджі, які рухаються до плазмалеми або тонопласта і зливаються з ними. Пухирці Гольджі можуть також відокремлюватися від попередньої цистерни або від її периферичних трубчастих утворень. Цей бік диктіосоми називають таким, що дозріває. Отже, диктіосома є динамічною структурою: з одного кінця з перехідних пухирців формується цистерна, яка поступово просувається до іншого, де і розпадається на пухирці Гольджі (рис. 1.3).

Рис. 1.3. Схема будови диктіосоми:
1 — цистерни; 2 — трубочки зі здутими кінцями; 3 — пухирці Гольджі
Назвемо ряд дуже важливих функцій, які виконує апарат Гольджі в рослинній клітині.
У цистернах диктіосом утворюються речовини матриксу клітинної оболонки — полісахариди геміцелюлоза і пектини, а пухирці Гольджі доставляють їх до плазмалеми, вбудовуються в неї і виділяють зазначені речовини назовні. До складу матриксу клітинної стінки входять також білки, що, як і ферменти, надходять із гранулярної ЕПС у перехідні пухирці. У диктіосомах ці білки тільки модифікуються: до них приєднується вуглеводна частина, і вони перетворюються в глікопротеїни, а потім транспортуються в пухирцях Гольджі до плазмалеми.
Через апарат Гольджі з гранулярної ЕПС надходять гідролітичні ферменти, що упаковуються в пухирці Гольджі, транспортуються до тонопласта і виділяються у вакуолю, забезпечуючи її лізосомну функцію.
Мембрани апарату Гольджі за допомогою пухирців Гольджі беруть участь у площинному рості плазмалеми і тонопласта: при виділенні вмісту пухирців у вакуолю і клітинну оболонку їх мембрана вбудовується в тонопласт або плазмалему.
Апарат Гольджі утворює лізосоми. Вони відокремлюються у вигляді пухирців від цистерн із боку, що дозріває, або від периферичних трубчастих утворень. У лізосомах містяться гідролітичні ферменти, що також походять від гранулярних мембран ЕПС.
Через апарат Гольджі здійснюється перетворення внутрішньоклітинних мембран: мембрани ЕПС перетворюються в плазмалему або тонопласт.
У диктіосомах відбуваються не лише синтез і модифікація речовин, але і їх сортування залежно від подальшого призначення. В одні пухирці Гольджі запаковуються речовини матриксу клітинної оболонки, і вони спрямовуються до плазмалеми; в другі — гідролітичні ферменти, і пухирці, які їх містять, дифундують до тонопласта; у третіх — накопичується велика кількість гідролітичних ферментів, і вони стають лізосомами в цитоплазмі.
Концепція ендомембран. Відповідно до концепції ендомемб-ран, яка була сформульована на початку 70-х рр. XX ст., усі внутрішні мембрани клітини складають ендомембранну систему. її компонентами є мембрани ЕПС, ядра, апарату Гольджі і пухирців Гольджі, плазмалема і тонопласт, а також зовнішня мембрана хлоропластів і мітохондрій. Внутрішні мембрани хлоропластів і мітохон-дрій до ендомембранної системи не належать, тому що, згідно із симбіотичною гіпотезою походження цих органоїдів, їх внутрішні мембрани утворилися з плазмалем клітин, які колись потрапили в клітину-господаря й утворили з нею єдиний симбіотичний організм.
Ендомембранна система дуже динамічна, у ній постійно відбувається потік мембран або їх перетворення в напрямку від мембран ЕПС через апарат Гольджі до плазмалеми і тонопласта.
Корпускулярні органоїди
Пластиди — це органоїди, притаманні лише рослинним клітинам. Вони присутні в клітинах усіх органів рослин: у стеблах, коренях, листках, квітках. Є три групи пластид, залежно від їх забарвлення: хлоропласти — зелені; хромопласти — жовтогарячі; лейкопласти — безбарвні.
Як правило, в клітині зустрічаються пластиди тільки одного типу. Усі пластиди мають деякі спільні риси будови. За формою вони найчастіше округлі, овальні, дископодібні. Зовні їх оточує оболонка з двох ліпопротеїнових мембран. Усередині міститься основна речовина — матрикс, яку в пластидах називають стромою. Пластиди розрізняються за компонентами, зануреними в строму. При цьому в матриксі майже всіх пластид є мембранні структури, хоча ступінь їх розвитку дуже різний. Пластиди, як і мітохондрії, мають власну ДНК. До пластид іноді зараховують пропластиди та етіопласти, які є попередниками цих органоїдів.
Хлоропласти мають зелене забарвлення, здійснюють фотосинтез. Вони є майже у всіх клітинах надземних органів рослин, куди потрапляє світло. Як правило, відсутні у клітинах коренів.
Хлоропласти присутні в усіх зелених фотосинтезуючих клітинах рослин. їх немає тільки в синьо-зелених водоростях (ціанобактеріях), точніше нема структурно оформлених зелених пластид. їх функції в клітинах синьо-зелених водоростей виконує скупчення мем-
ста, стрічкоподібна, ребриста, циліндрична тощо).
Хлоропласт оточений подвійною ліпопротеїновою мембраною — оболонкою, що регулює обмін речовин між ним і цитоплазмою (рис. 1.5). Усередині хлоропласта міститься основна речовина — матрикс, або строма. У матрикс занурена система мембран, які у хлоропластах називаються ламелами. Вони утворюють плоскі пухирці — тилакоїди, зібрані в стопки — грани. Грани зв'язані між собою в єдину систему трубчастими, витягнутими тилакоїдами строми, або міжгранними тилакоїдами.

розміри, і в клітині їх міститься всього один або декілька. Форма хроматофорів незвичайна і химерна (зірча-
Рис. 1.4. Хлоропласти у клітинах моху
Хлоропласти мають власну білоксинтезуючу систему з ДНК, рРНК і рибосомами. Хлоропластна ДНК передається по материнській лінії (через пропластиди). Від батьківських рослин — через пилкове зерно — хлоропластна ДНК не успадковується.
Рис. 1.5. Будова хлоропласта:
1 — зовнішня оболонка; 2 — внутрішня оболонка; 3 — строма;
4 — краплини ліпідів; 5 — грана;
6 — тилакоїди строми; 7 — крохмальні зерна; 8 — оболонка хлоропласта; 9 — рибосоми
Кільцева ДНК хлоропластів відрізняється від ядерної як за наявною в ній інформацією, так і за фізико-хімічними властивостями. Хлоропласти мають власну ДНК-полімеразу, тому її ДНК репліку-ється в самих хлоропластах.
Хлоропласти вищих рослин містять два види зелених пігментів: хлорофіл а і хлорофіл Ь. Хлорофіл інтенсивно поглинає промені в червоній області спектра (680— 730 нм) і в синьо-фіолетовій (470 нм і нижче), а пропускає жовті та зелені промені.
Крім хлорофілу, в хлоропластах містяться ще жовтогарячі пігменти — каротиноїди. Серед них відомі каротини і ксантофіли. Каротини мають жовтогарячий колір, а ксантофіли - жовтий. Зазвичай, інтенсивніше забарвлення хлорофілу маскує каротиноїди, тому хлоропласти і листя мають зелене забарвлення.
До складу хлоропластів входять багато ферментів, що беруть участь у здійсненні численних реакцій фотосинтезу і пов'язаних із синтезом білків, нуклеїнових кислот тощо.
Основна функція хлоропластів — фотосинтез (процес засвоєння сонячної енергії і перетворення її в енергію хімічних зв'язків).
Процес фотосинтезу поділяють на світлову і темнову стадії. У процесі світлової стадії відбувається запасання енергії світла в АТФ і утворення відновника НАДФ • Н, які витрачаються в темновій стадії на відновлення С02 до рівня вуглеводів. Реакції світлової стадії проходять у ламелах тилакоїдів гран, а темнової — в стромі.
Хлоропласти в клітинах утворюються з пропластид, дрібних тілець (1—1,5 мкм), оточених подвійною мембраною.
Хлоропласти розмножуються поділом. Із закінченням росту листя припиняється поділ хлоропластів.
Хлоропласти звичайно розташовуються так, щоб їх освітлювало розсіяне світло. Вони уникають прямих сонячних променів, які руйнують хлорофіл.
Походження хлоропластів. Оскільки хлоропласти відносно автономні у відношенні до ядра структури, здатні до поділу, росту й диференціювання, виникла гіпотеза стосовно того, що в прадавні часи це були самостійні організми. Ймовірно, хлоропласти сформувалися внаслідок симбіозу якогось автотрофного організму (можливо, ціанобактерії), здатного трансформувати сонячну енергію, з гетеротрофною клітиною. Пластиди і фотосинтезуючі прокаріоти досить подібні між собою (кільцева структура ДНК, відсутність гісто-нів, розміри рибосом тощо). З точки зору еволюції це було взаємовигідним, і тому такий симбіотичний союз зберігся у вигляді сучасної евкаріотичної рослинної клітини.
Хромопластами називають нефотосинтезуючі забарвлені пластиди, які містять, в основному, червоні, помаранчеві й жовті пігменти. Вони, як правило, розвиваються з хлоропластів, мають приблизно такі самі розміри та форму, досить схожі й за структурою.
Усередині хромопласти заповнює строма, в якій іноді зустрічаються рудименти мембранної системи: окремі тилакоїди, скупчення трубочок. Однак замість системи фотосинтетичних мембран в них є структури, багаті на каротиноїди.
Хромопласти зустрічаються в пелюстках квіток деяких рослин (жовтеці, кульбаби та ін.), у плодах (шипшина, перець, горобина, помідори, кавун), рідше — у вегетативних органах (морква).
Фізіологічна функція хромопластів детально не вивчена. Яскраве забарвлення, можливо, принаджує комах для запилення та поширення насіння.
Лейкопласти — дрібні безбарвні пластиди, які переважно містяться в клітинах тканин, на які не потрапляє світло (сім'ядолі, ендосперм насіння, кореневища, бульби, корені), рідше — у тканинах
на світлі (шкірка).
Основна функція лейкопластів — накопичення поживних речовин, які надходять у запасаючі органи.
Внутрішня мембрана лейкопластів формує поодинокі тилакоїди. Лейкопласти мають власну ДНК, рибосоми, а також різноманітні ферменти. Лейкопласти, в яких синтезується і відкладається крохмаль у незелених частинах рослин, називають амілопластами, рослинні олії — олеопластами, білки — протеїнопластами.
Амілопласти — найрозповсюдженіші лейкопласти, в них накопичується запасний крохмаль у вигляді зерен. Амілопласти є в запасних тканинах та органах, а саме: сім'ядолях, ендоспермі, бульбі, а також у кореневому чохлику.
Протеїнопласти містять у стромі запасний білок у вигляді кристалів або аморфної маси. Лейкопласти такого типу є рідкісним явищем (кореневі бульби орхідей).
Олеопласти зустрічаються ще рідше. У їхній стромі збираються жири (олії) у вигляді крапельок або пластоглобул (однодольні).
У кореневищах деяких видів півників у лейкопластах може накопичуватися крохмаль або жир.
Взаємоперетворення пластид. Усі пластиди — хлоропласти, хромопласти, лейкопласти — можуть розвиватися безпосередньо з пропластид, що містяться в клітинах твірної тканини.
Мітохондрії. По всій цитоплазмі клітин розсіяні численні міто-хондрії, довжина яких — від одного до кількох мкм, а ширина — до 0,5 мкм. Мітохондрії — органоїди, присутні як у рослинних, так і у тваринних клітинах. У рослинній клітині може міститися від декількох десятків до декількох тисяч мітохондрій. Кількість їх залежить від віку і метаболічної активності клітин. Деякі водорості (хлорела, хламідомонада) мають у клітині лише одну велику міто-хондрію.
Мітохондрії оточені подвійною ліпопротеїновою мембраною. Зовнішня мембрана відрізняється від внутрішньої співвідношенням білків і ліпідів, набором ферментів, проникністю. Внутрішня міто-хондріальна мембрана утворює впинання — гребені, або кристи, розмір та кількість яких залежать від функціональної активності мітохондрій (рис. 1.6). Кристи утворюють велику поверхню для реакцій, що проходять на внутрішній мембрані. Простір між кристами заповнено основною речовиною — матриксом, у якому помітні гра
нулярний і фібрилярний компоненти. Гранули представлені рибосомами і фосфатом кальцію, фібрили — це нитки ДНК.
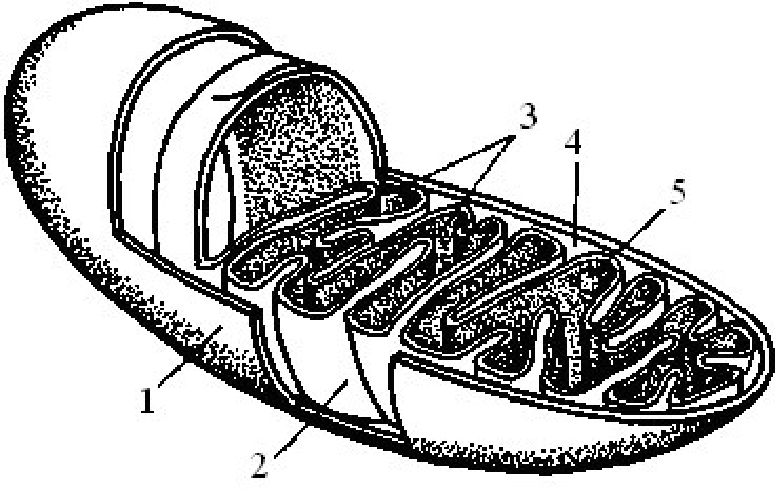
Рис. 1.6. Схема будови мітохондрїї:
1 — зовнішня мембрана; 2 — внутрішня мембрана; 3 — кристи; 4 — міжмембранний простір; 5 — матрикс
Подібно до пластид мітохондрії є напівавтономними органоїдами, оскільки в них відбувається незалежний від ядра білковий синтез. У матриксі мітохондрій локалізована власна білоксинтезуюча система з кільцевих молекул ДНК, РНК і 70S рибосомами. Мітохо-ндріальний геном кодує до 50 мембранних і матриксних білків.
Основна функція мітохондрій — енергетична. У них відбувається процес дихання — окиснення поживних речовин, насамперед вуглеводів, з виділенням енергії, що запасається у формі АТФ. Розклад вуглеводів починається в матриксі цитоплазми (гліколіз), а продовжується в матриксі мітохондрій (цикл Кребса). Синтез АТФ відбувається на внутрішній мітохондріальній мембрані.
Мітохондрії постійно рухаються й збираються там, де потрібна енергія. Тривалість життя мітохондрій — декілька діб. Мітохондрії розмножуються в клітинах шляхом поділу.
Детальне вивчення структури та геному мітохондрій дає можливість припустити, що вони, як і хлоропласти, походять від бактерій, які випадковим симбіозом прижилися в більших гетеротрофних клітинах — попередниках евкаріот.
Лізосоми — дрібні округлі органоїди діаметром 0,5-2,5 мкм. Вважають, що у тваринних клітинах їх більше, ніж у рослинних. Лізосоми оточені однією ліпопротеїновою мембраною, усередині знаходиться матрикс, що містить гідролітичні ферменти, які розщеплюють різні органічні речовини — білки, нуклеїнові кислоти, полісахариди та ін. Лізосоми утворюються з цистерн апарату Гольджі або з гладких мембран ЕПС. Функція лізосом — розщеплення речовин і окремих ділянок цитоплазми, тобто локальний автоліз.
У лізосомах розщеплюються «відпрацьовані» речовини й органоїди, що у даний момент онтогенезу вже виконали свої функції і не потрібні клітині. Продукти їх розпаду використовуються на побудову нових, необхідних їй речовин і структур.
У лізосомах розпадаються речовини, часто шкідливі, які потрапляють у клітину шляхом ендоцитозу. Продукти їх розщеплення використовуються клітиною.
При відмиранні клітин порушуються мембрани лізосом, їхні ферменти виходять у цитоплазму, відбувається автоліз усього живого вмісту, що спричиняє очищення порожнини мертвих клітин (трахеї і трахеїди ксилеми, клітини механічних тканин тощо).
Рибосоми — це дрібні клітинні органоїди, які можна побачити тільки під електронним мікроскопом. Рибосоми мають овальну форму. Кожна рибосома складається з двох субодиниць — великої і малої. Рибосоми розпадаються на дві субодиниці при зниженні в середовищі концентрації іонів Мр 2+.
Рибосоми складаються з приблизно рівної кількості РНК і білка. Між субодиницями при їх об'єднанні в цілу рибосому залишається щілина, у якій проходить матрична РНК, а на великій субодиниці є ще борозенка, в якій розміщається і по якій сповзає білкова молекула, що синтезується.
Рибосоми — центри синтезу білка в клітині. У них відбувається розміщення і з'єднання амінокислот у поліпептидний ланцюг відповідно до генетичної інформації, одержуваної з ядра через матричну РНК. Синтез білка здійснюється на окремих рибосомах і на їх скупченнях, пов'язаних мРНК, — полісомах.
Рибосоми поширені в клітині: вони прикріплені до мембран ЕПС (гранулярні мембрани) і зовнішньої ядерної мембрани, містяться в матриксі цитоплазми, в ядрі, у хлоропластах і мітохондріях.
Кількість рибосом у клітині може сягати десятків мільйонів. Вона залежить від інтенсивності синтезу білка, фізіологічного стану клітини, дії факторів довкілля (світло, температура, вологість, умови мінерального живлення та ін.).
рРНК синтезується в ядерці, а рибосомні білки — у цитоплазмі, звідки частина їх надходить у ядро. В ядерці починається складання рибосом, а закінчується воно вже в цитоплазмі.
Мікротільця, або мікрочастинки, — дрібні клітинні органоїди — округлі й ниткоподібні. До перших належать пероксисоми, гліо-ксисоми, сферосоми, до других — мікротрубочки і мікрофіламенти.
Пероксисоми містяться в клітинах фотосинтезуючих тканин. Тут відбувається фотодихання — процес, функціонально пов'язаний з хлоропластами і мітохондріями. При фотодиханні в пероксисомах утворюється і розщеплюється перекис, звідси їх назва.
Гліоксисоми містяться в насінні олійних рослин і функціонують при їх проростанні. У них відбувається процес перетворення запасних жирів у вуглеводи (глюкозу), які транспортуються в паросток і живлять його до виходу з грунту.
Сферосоми — це органоїди, що запасають жир (олії). Вони присутні в насінні олійних культур і містять ферменти, які синтезують жири і в процесі дозрівання насіння накопичують жир.
Мікротрубочки являють собою тонкі циліндричні структури діаметром 24—25 нм і непостійної довжини (до декількох мкм). Мікротрубочки виконують у клітині ряд важливих функцій:
- утворюють ахроматинове веретено при поділі ядра і фрагмоп-ласт при поділі клітини;
- розташовуються під плазмалемою, визначаючи орієнтацію в клітинній оболонці мікрофібрил целюлози;
- спрямовують рух пухирців Гольджі, що несуть речовини матриксу клітинної оболонки, до тих місць плазмалеми, де активно синтезується целюлоза;
- у рухливих клітин мікротрубочки є компонентами джгутиків і війок.
Мікрофіламенти — це нитчасті структури, що складаються із субодиниць білка актину, близького за властивостями до актину м'язів. Мікрофіламенти генерують рух цитоплазми та її органоїдів.
Мікротрубочки і мікрофіламенти, завдяки досить жорсткій структурі, відіграють ще одну важливу роль у клітині — утворюють цитоскелет цитоплазми.
Ядро — найбільший і найважливіший корпускулярний органоїд еукаріотичних клітин. Це сферичне тіло діаметром 5-10 мкм, що містить більшу частину генетичної інформації у вигляді довгих ланцюгів ДНК. Цей органоїд присутній в усіх рослинних клітинах, за винятком члеників ситоподібних трубок флоеми.
У молодій клітині ядро звичайно перебуває в центрі. При рості і спеціалізації клітини, коли її центральну частину займає вакуоля, ядро разом із цитоплазмою відтискується до клітинної стінки.
Форма ядра у паренхімних клітинах частіше округла, дископодібна, а у прозенхімних — подовжена, веретеноподібна, ниткоподібна. У деяких клітинах форма ядра лопатева або навіть розгалужена.
Ддро виконує дві найважливіші функції:
- це місце збереження і відтворення генетичної інформації, що передається від материнської клітини дочірнім у процесі поділу;
- контролює життєдіяльність клітини, її ріст і розвиток за допомогою насамперед мРНК, що синтезується в ядрі і несе інформацію про склад білків клітини в той чи інший момент її життя. Склад білків-ферментів визначає напрямок шляхів метаболізму, отже, і властивості клітин та організму в цілому. Якщо з клітини видалити ядро, вона гине.
У ядрі зосереджена майже вся ДНК клітини (99 %) і набагато менше РНК. 1 %, що залишився, припадає в клітині на частку ДНК хлоропластів і мітохондрій.
Основними структурними компонентами ядра є ядерна оболонка, ядерний матрикс, хроматин і ядерця.
Ядерна оболонка складається з двох елементарних мембран, між якими знаходиться порожнина — перинуклеарний простір. Зовнішня мембрана безперервно зв'язана з мембранами ЕПС, а перинуклеарний простір сполучається з порожнинами її канальців і цистерн.
Основну речовину ядра — ядерний матрикс — називають нукле-оплазмою, або каріоплазмою. Нуклеоплазма є безструктурною масою, у якій розрізняють гранули, подібні до рибосом. Вона пов'язана з матриксом цитоплазми за допомогою ядерних пор. Функції ну-клеоплазми полягають у взаємозв'язку всіх структурних компонентів ядра і здійсненні ряду ферментних реакцій.
Хроматин — це скупчення тонких ниток, занурених у матрикс. Нитчастий компонент називають еухроматином, а гранулярний — гетерохроматином. Хроматин являє собою структурну видозміну хромосом. В інтерфазі і метаболічно активному ядрі хромосоми дуже деспіралізуються і гідратуються, утворюючи в матриксі слабко помітну мережу еухроматину. Гранули гетерохроматину — це ділянки, де спіралізація й упакування хромосом зберігається також у ядрі, яке не ділиться. Хроматин характерний для клітин у період їх активної життєдіяльності, а в період поділу він організовується в щільно запаковані хромосоми. У хроматині зосереджена майже вся ДНК ядра і ферменти її синтезу.
Функції хроматину полягають у такому:
- у синтезі специфічних для даного організму нуклеїнових кислот, що направляють синтез специфічних білків;
- у передачі спадкових властивостей від материнської клітини дочірнім, для чого хроматинові нитки в процесі поділу упаковуються в хромосоми.
Ядерце — кулясте, добре помітне під мікроскопом тільце діаметром 1-3 мкм. Воно формується на ділянці хроматину, яку називають ядерцевим організатором. Функція ядерця — синтез рРНК і формування початкових форм рибосом шляхом об'єднання рРНК з білками, що надходять з цитоплазми.
Запасні поживні речовини. Багато їх накопичується в насінні, плодах, вегетативних органах (коренях, бульбах, кореневищах, цибулинах). Основними запасними поживними речовинами рослин є вуглеводи, білки, жири (олії).
Крохмаль — основний запасний вуглевод рослин. У хлоропластах у процесі фотосинтезу утворюється асиміляційний (первинний) крохмаль. Однак він швидко гідролізується до цукрів, що відтікають у запасаючі органи. Там в амілопластах у вигляді зерен відкладається запасний (вторинний) крохмаль. Крохмальні зерна (амілопласти) різноманітні за формою і розмірами: округлі, яйцеподібні, багатогранні, від 2 до 150 мкм. Найдрібніші крохмальні зерна в насінні рису і гречки, найбільші — у бульбах картоплі.
Крохмаль складає велику частину насіння злаків, бобових, бульб картоплі (30-85 %).
Запасні білки найчастіше відкладаються у вигляді алейронових (білкових) зерен, які зустрічаються переважно в насінні.
Запасні білки рослин відіграють дуже важливу роль у харчуванні людини. У наш час потреби людини в харчовому білку на 70—80 % задовольняються за рахунок рослинних білків. Насіння зернових культур містить до 20 % білка. Найбагатше запасним білком насіння бобових рослин — 30-35 %, соя — до 40 %.
Рослинні жири рідкі, тому їх ще називають оліями. Найбагатші на олії клітини насіння, плодів, серцевини і кори стебел, кореневища. Більшість рослин (до 90 % усіх родин) як запасні поживні речовини накопичують олії. Насіння багатьох рослин містить до 50 % і більше олії (соняшник, гірчиця, соя, софлор, рижій, арахіс та ін.). Олії є вигідною запасною поживною речовиною для рослин, тому що вони багатші на енергію, ніж крохмаль і білки.
Вакуолі та їх функції. Вакуоля являє собою ділянку всередині протопласта, оточену мембраною і заповнену клітинним соком. Мембрану вакуолі, як уже зазначалося, називають тонопластом.
У меристематичних клітинах звичайно вакуолей немає або вони дуже дрібні. У процесі росту і розвитку клітини дрібні вакуолі поступово зливаються в одну велику центральну вакуолю, що характерна для дорослої рослинної клітини. При цьому протопласт відсувається і притискається до клітинної оболонки.
Вакуолі рослинних клітин виконують три основні функції:
1. Запасаюча. Тут відкладаються запасні поживні речовини (вуглеводи, білки, органічні кислоти та ін.) і продукти життєдіяльності клітини, функції яких різноманітні і поки ще не всі з'ясовані.
2. Осмотична. Завдяки досить високій концентрації речовин у клітинному соку утворюється його осмотичний тиск: створюється сисна сила, що забезпечує надходження води в клітину і пересування її з клітини в клітину; виникає тургор, що створює опору клітинам і м'яким органам, надає їм міцності та положення в просторі.
3. Лізосомна. Гідролітичні ферменти, присутні у клітинному соку, розщеплюють полімерні речовини і структури, що вже виконали свою роль у клітині та проникають у вакуолю шляхом інвагінації тонопласта. Прості сполуки (цукри, амінокислоти), які утворилися в результаті розщеплення, надходять назад у цитоплазму і знову використовуються в метаболізмі.
Клітинний сік, його хімічний склад. Клітинний сік — це водний розчин різних органічних і мінеральних речовин. Вода є його основним компонентом (до 98 %) і міститься у вакуолях усіх клітин. Інші речовини у вакуолях можуть перебувати у вигляді справжніх розчинів, колоїдних, емульсій, суспензій і твердих відкладень.
У клітинному соку різних рослин містяться вуглеводи, білки, амінокислоти, органічні кислоти, глікозиди, дубильні речовини, алкалоїди, терпени, пігменти, мінеральні солі тощо.
У клітинному соку майже всіх рослин присутні вуглеводи — цукри (моносахариди і дисахариди) і деякі розчинні полісахариди.
У складі клітинного соку ряду рослин містяться запасні білки. Особливо їх багато у дозрілому насінні (пшениця, горох, рицина та ін.). Крім білків, у вакуолях знаходяться різні амінокислоти.
Звичайно органічні кислоти присутні в клітинному соку в значній кількості у вільному стані або у вигляді солей. Саме їх присутність визначає кислу реакцію клітинного соку. Найчастіше зустрічаються щавлева, оцтова, яблучна, винна, лимонна кислоти.
Глікозиди часто мають гіркий смак і специфічний запах. Наприклад, гірчиця містить глікозид синігрин, що додає їй специфічного запаху і смаку.
Дубильні речовини, або таніни накопичуються в порожнині клітин після відмирання їх протопластів. Значну їх кількість (до 20— 30 % і більше) містять кора дерев (дуб, сосна, верба, евкаліпт та ін.), листя (сумах, чай) та плоди (хурма, терен та ін.). Дубильні речовини мають в'яжучі й антисептичні властивості.
Алкалоїди — це гетероциклічні азотовмісні сполуки. Вони мають лужні властивості й утворюють солі з органічними кислотами, які розчиняються в клітинному соку. Алкалоїди спричиняють сильну фізіологічну дію на організм тварин і людини. У малих дозах їх використовують як лікарські препарати, а у великих вони є отрутами (морфін, кодеїн, папаверин з маку снодійного, кокаїн із листків коки, кофеїн із листків чаю, зерен кави, бобів какао, атропін із бела-донни, дурману та ін.)
Терпени — це ненасичені вуглеводні. До цієї групи сполук, що містяться в клітинному соку деяких рослин, належать каучук і тута. У промислових масштабах каучук одержують з гевеї і використовують для виготовлення гуми, а в медицині — пластирів і гірчичників. Гуту добувають з гваюли і використовують для виготовлення ізоляційного матеріалу для покриття підводних кабелів.
Часто клітинний сік має забарвлення, яке надають розчинені в ньому пігменти. Серед пігментів клітинного соку найчастіше зустрічаються антоціани й антохлори.
Антоціани — це найважливіші пігменти рослин, вони забарвлюють квіти, плоди, листки в блакитний, синій, рожевий, червоний, фіолетовий кольори з різними відтінками. Антоціани визначають забарвлення квітів гіацинтів, троянд, незабудок, маку тощо, плодів чорної смородини, вишні, сливи тощо, рідше вегетативних органів (корені столового буряку, листки червонокачанної капусти).
Антохлори — це жовті пігменти, які забарвлюють переважно пелюстки квіток (льонок, первоцвіт) і плоди (лимони, апельсини).
Тверді відкладення в клітинному соку — найчастіше нерозчинні солі кальцію, особливо щавлевокислий кальцій (оксалат кальцію).
1.6. Осмотичні властивості рослинної клітини
Усі клітини рослин відокремлені від довкілля клітинною оболонкою та плазматичною мембраною, що здатні регулювати не тільки кількість і тип речовин, які проходять крізь них, а часто й напрям їхнього руху. Для здійснення всіх процесів життєдіяльності в клітину з довкілля повинні надходити вода та поживні речовини.
Дифузія — це спонтанний процес, що зумовлює переміщення будь-якої речовини з однієї ділянки в іншу, де концентрація даної речовини менша.
Рослинна клітина являє собою осмотичну систему, де один розчин — це зовнішнє середовище (або вакуоля сусідньої клітини), а другий — клітинний сік власної вакуолі. Мембрани живих клітин здатні транспортувати лише певні молекули або йони розчинених речовин, проявляючи вибірковість, яка залежить від природи мембрани. Тому такі мембрани називають вибірково проникними, або напівпроникними, а дифузію води крізь ці мембрани — осмосом. Оскільки в усіх біологічних системах розчинником є вода, тому осмос для них — це дифузія води крізь вибірково проникну мембран. Напівпроникними мембранами є плазмалема і тонопласт.
В осмосі насамперед звертають увагу на осмотичний тиск — силу, яку необхідно прикласти, щоб перешкодити руху води через напівпроникну мембрану в бік розчину з більшою концентрацією. Чим вища концентрація розчину, тим вищий його осмотичний тиск. Чим більша різниця в концентраціях розчинів, розділених напівпроникною перетинкою, тим інтенсивніше буде потрапляти вода в бік більш концентрованого розчину.
Клітинна оболонка захищає протопласт від розриву під дією гідростатичного тиску вакуолі.
Інтенсивність проникнення води в клітину залежить від осмотичного тиску рідини у вакуолі й у зовнішньому середовищі, точніше, від різниці осмотичного тиску всередині і поза клітиною. Силу, з якою вода проникає усередину вакуолі живої клітини, називають сисною силою.
Стосовно концентрації клітинного соку зовнішні розчини можна розділити на три типи: ізотонічні, або ізоосмотичні, осмотичний тиск яких дорівнює осмотичному тиску в клітині; гіпотонічні, осмотичний тиск яких нижчий, ніж у клітині; гіпертонічні, осмотичний тиск яких вищий, ніж у клітині.
Якщо рослинну клітину помістити в ізотонічний розчин, то руху води ні в клітину, ні з клітини не відбуватиметься. Якщо ж клітина опиниться в гіпотонічному розчині, то вода прямуватиме в її вакуолю. У міру поглинання клітиною води об'єм її вакуолі збільшується, вона тисне на протопласт і через нього на клітинну оболонку, що, розтягуючись і напружуючись, у свою чергу, починає тиснути на протопласт. Тиск розтягнутої клітинної оболонки на протопласт називають тургорним тиском, а напружений стан клітини, який при цьому виникає, — тургором.
Чим концентрованіший розчин, тим вищий осмотичний тиск. Деякі речовини, зокрема неорганічні солі, у воді здатні до електролітичної дисоціації, внаслідок чого концентрація частинок зростає. Відповідно зростає осмотичний потенціал такого розчину.
Знання осмотичного потенціалу необхідне у разі проведення різноманітних екологічних досліджень, в результаті чого роблять висновок про здатність рослин поглинати воду із ґрунту та утримувати її незалежно від впливу умов навколишнього середовища.
Осмотичний потенціал коливається в межах 0,1 ...20 МПа. У водних рослин він найменший і досягає 0,1 МПа, а у багатьох галофітів — до 20 МПа. Осмотичний потенціал у мезофітів коливається від 0,5 до 3 МПа.
Під впливом різноманітних факторів величина осмотичного потенціалу може змінюватися навіть у сусідніх клітинах.
При втраті рослиною води в умовах нестачі вологи в грунті і сильному випаровуванні тургор клітин знижується, їх стінки зморщуються, настає зів'янення: листя обвисає, трав'янисті стебла никнуть.
Дослідами доведено, що концентрація клітинного соку, а отже, і осмотичний тиск у клітинах верхніх листків рослин вищий, ніж у нижніх, а в клітинах кореня більше, ніж у ґрунті.
Осмотичним властивостям належить дуже важлива роль у будові і життєдіяльності рослин:
Осмос визначає поглинання води і пересування її по рослині.
Осмотичні властивості забезпечують тургор рослині, особливо її м'яким органам (листки, трав'янисті стебла), що надає їм форму, міцність і дозволяє підтримувати положення в просторі.
Плазмоліз. Коли клітина контактує з гіпертонічним розчином, вода починає виходити з клітини внаслідок осмосу через плазматичну мембрану. Спочатку воду втрачає цитоплазма, а потім через
тонопласт вода виходить також із вакуолі. Об'єм внутрішньоклітинного вмісту зменшується, протопласт, оточений плазматичною мембраною, зморщується і врешті-решт плазмалема відстає від клітинної оболонки, тобто спостерігається плазмоліз.
Плазмоліз — це відокремлення пристінного шару цитоплазми від твердої оболонки рослинної клітини.
Спостерігається плазмоліз лише у живих клітинах внаслідок стискання протопласта під впливом гіпертонічного, відносно до клітинного соку, плазмолітика. За повільного плазмолізу клітини тривалий час залишаються живими. За наявності доступної для клітини води вони легко відновлюють стан тургору. Тривалий плазмоліз зумовлює загибель клітин. Явище плазмолізу використовують для визначення осмотичного потенціалу, в'язкості цитоплазми, проникності клітинних мембран тощо.
Цей процес оборотний. Якщо плазмолізовану клітину перенести в чисту воду або слабоконцентрований розчин, вода знову надходи-тиме в клітину, в результаті чого відбуватиметься деплазмоліз.
Набухання. Поряд із осмотичними силами в клітинах існують сили набухання. У деяких частин рослин вода поглинається виключно через набухання, наприклад у насіння. Вода в клітину проникає в разі набухання завдяки дифузії. Стан набухання протоплазми має вирішальне значення для інтенсивності всього обміну речовин, оскільки гідратація протоплазматичних білків необхідна для підтримки ультраструктури та функціональної активності органоїдів.
Піноцитоз — це інвагінація поверхневої мембрани, завдяки чому відбувається захоплення краплин рідини.
1.7. Клітинна оболонка
Усі клітини рослинних організмів мають тверду еластичну клітинну оболонку, або клітинну стінку, що є продуктом життєдіяльності протопласта. Наявність клітинної оболонки — істотна ознака, що відрізняє рослинну клітину від тваринної.
Клітинна оболонка визначає форму клітини.
Оболонки, особливо тверді й потовщені, служать механічною опорою клітинам і органам рослин.
Клітинна оболонка виконує різноманітні захисні функції: захищає протопласт від різних ушкоджень — механічних, шкідниками, тваринами, від проникнення патогенних мікроорганізмів, від
втрати води і висихання.
Оболонка бере участь в осмотичних властивостях клітини, обмежуючи розтягнення протопласта і запобігаючи його розриву під дією гідростатичного тиску вакуолі.
Клітинна оболонка бере участь у транспорті води і розчинених у ній речовин, що долають клітинну стінку, перш ніж потрапити в цитоплазму або при виході з неї.
Склад і будова клітинних оболонок варіює залежно від типу клітин та виду рослин. Змінюються вони й у процесі індивідуального розвитку клітини.
Склад клітинної оболонки. Основними речовинами клітинної оболонки є вуглеводи (целюлоза, геміцелюлоза, пектини), білки і вода. Целюлоза — обов'язковий компонент стінок усіх рослинних клітин. Вона утворює каркас, або кістяк, клітинної оболонки. Геміцелюлоза, пектини і білки складають матрикс оболонки, який оточує целюлозний каркас.
Целюлоза, або клітковина, — вуглевод-полімер. У клітинній оболонці молекули целюлози зібрані в пучки — мікрофібрили, у яких вони розташовуються паралельно одна до одної.
Целюлоза має дуже широке практичне застосування у целюлозно-паперовій, хімічній і інших галузях промисловості.
Геміцелюлоза — менш стійкий аморфний компонент клітинної оболонки. Входить до складу матриксу, заповнюючи проміжки між мікрофібрилами. Багато геміцелюлози в оболонках молодих клітин.
Пектини, або пектинові речовини, також мають вуглеводну природу. Перебуваючи у колоїдному стані, пектини надають клітинній оболонці еластичності.
Пектинові речовини мають желеутворюючі властивості, на яких засноване приготування варення, джему, фруктових желе, мармеладу, карамельних начинок тощо.
На частку білків у клітинній оболонці припадає 5—10 % її маси. До їх складу входить насамперед структурний білок екстенсин. Крім того, в оболонці виявлені деякі білки-ферменти.
Вода в клітинній оболонці відіграє роль розчинника і забезпечує транспорт речовин через клітинну стінку: чим більше води в оболонці, тим вища її проникність. Вона утворює колоїдний розчин пектинових речовин.
Структура клітинної оболонки. У процесі цитокінезу між дво
ма дочірніми клітинами спочатку утворюється пектинова серединна пластинка, а потім на неї з боку кожної клітини накладається первинна оболонка. В одних тканинах вона зберігається протягом усього життя клітин (меристеми, кореневі волоски, мезофіл листка), в інших — після припинення росту клітин на первинну оболонку накладається вторинна (механічні тканини, ксилема).
Первинна і вторинна клітинні оболонки розрізняються за своїм складом і будовою.
Первинна клітинна оболонка дуже тонка — 0,1-0,5 мкм. До її складу входять целюлоза (5-15 %), геміцелюлоза (до 30 %), пектини (до 5 %), білки (5-10 %) і багато води (60-90%). У зв'язку з низьким вмістом целюлози її мікрофібрили в первинній оболонці розташовуються пухко і пов'язані між собою молекулами матриксу. У наш час учені пропонують кілька різних схем молекулярної структури первинної клітинної оболонки. Вважається, що сітка, утворена мік-рофібрилами целюлози і молекулами геміцелюлози, забезпечує міцність клітинної оболонки, а пектинові речовини — її еластичність.
Первинна оболонка неоднакова за товщиною на всій своїй поверхні. Вона має тонкі ділянки, де мікрофібрили розташовані особливо пухко і через які проходять плазмодесми, що з'єднують цитоплазми сусідніх клітин. Ці ділянки називають первинними поровими полями. Над ними у вторинній оболонці утворюються пори.
Клітини меристеми, які постійно діляться, а також більшість спеціалізованих зрілих клітин (наприклад, мезофілу листка) мають лише первинні клітинні оболонки. Такі клітини здатні змінювати свою форму, знову ділитися і диференціюватися в новий тип клітин. Вони беруть участь у заживленні ран і регенерації у рослин. Первинні клітинні оболонки різні за товщиною, вони мають і тонкі ділянки, які називають первинними поровими полями. Через них проходять плазмодесми.
Однак у більшості клітин на внутрішню поверхню (назовні від плазматичної мембрани) відкладаються додаткові шари целюлози й утворюється вторинна клітинна оболонка. Вміст целюлози у вторинній оболонці досягає 60 %, її фібрили в кожному шарі лежать паралельно, а в сусідніх — під кутом один до одного, в матриксі з'являються різні включення — лігніни, суберин, кутин, віск тощо. Цим досягається значна міцність і твердість вторинної оболонки, вона втрачає еластичність, а клітина — здатність до подальшого росту. Головний компонент жорсткої оболонки — лігнін, присутність якого типова для вторинних стінок клітин деревини. Лігнін скріплює целюлозні волокна й утримує їх у певному місці. Елементи ксилеми (трахеї, трахеїди), механічні клітини склеренхіми зазнають інтенсивної лігніфікації. Суберин у поєднанні з воском спричинює опробковіння клітинної оболонки, що знижує її проникність. Кутин формує кутикулярний шар назовні стінки, завдяки чому вона стає майже непроникною.
Функції клітинної оболонки. Клітинна оболонка є секреторним продуктом протопласту і в зрілій клітині, ймовірно, нежива. Однак в ній є білки, серед яких і ті, що мають ферментативну активність.
Клітинна оболонка забезпечує окремим клітинам і рослині в цілому механічну міцність і опору. Вона визначає розмір, форму та стабільність рослинної клітини, захищає протоплазматичну мембрану від руйнування в разі дії гідростатичного тиску, що формується всередині клітини. Клітинна оболонка є протиінфекційним бар'єром, бере участь в поглинанні, транспортуванні та виділенні речовин. Через пори в клітинних оболонках проходять цитоплазматичні тяжі, які зв'язують вміст окремих клітин, тобто об'єднують всі протопласти в одну систему — симпласт.
Є дані, що вуглеводні компоненти клітинної оболонки, взаємодіючи із фітогормонами, викликають фізіологічні зміни у клітині.
Пори. Вторинна клітинна оболонка не відкладається над первинними поровими полями, а їх оточує. Виникають поглиблення у вторинній оболонці, які називають порами. Звичайно пори утворюють пари, тому що в суміжних стінках сусідніх клітин вони розташовуються одна проти одної. За формою каналів пори поділяють на прості й облямовані.
Прості пори мають циліндричний канал, діаметр його на всій відстані приблизно однаковий. Такі пори характерні для паренхіми, механічної тканини та ін.
В облямованих порах канал має лійкоподібну форму. Він звужується усередину клітини у зв'язку з ростом в товщину вторинної оболонки. Облямовані пори властиві елементам, які проводять воду (ксилема). Елементи ксилеми мертві, вони втрачають живий вміст. Одночасно руйнуються і плазмодесми в порових мембранах, а також з них зникають геміцелюлози і пектини, залишається тільки сітка з мікрофібрил целюлози, через яку проходить струм води. У багатьох хвойних рослин посередині замикаючої плівки облямованих йор утворюється потовщення — торує.
Іноді замикаюча плівка пори руйнується і утворюється наскрізний отвір — перфорація. Причому на поровій мембрані може виникати багато дрібних перфорацій.
Фізико-хімічні видозміни клітинної оболонки. У деяких клітин оболонки залишаються незмінними, целюлозними до кінця життя. Однак у багатьох тканин під час диференціювання клітин мають місце фізико-хімічні видозміни: здерев'яніння, окорковіння, кутинізація, ослизнення, мінералізація.
Здерев'яніння, або лігніфікація, — найпоширеніша видозміна оболонок клітин вищих рослин, які закінчили ріст. Вона відбувається при інкрустації їх лігніном. Це складний полімер фенольної природи, дуже стійкий і важкорозчинний. Здерев'яніння надає клітинній оболонці твердості та міцності, хоча значно зменшує її еластичність. Клітини зі здерев'янілими стінками втрачають живий вміст, відмирають. Здерев'яніння зазнають насамперед механічні тканини і провідна тканина ксилеми.
Окорковіння відбувається при нагромадженні в клітинних оболонках суберину — стійкої полімерної жироподібної речовини. Стінки, що окорковіли, стають непроникними для води і газів, і вміст клітин відмирає. Корковіють звичайно стінки клітин покривних тканин і клітин, що відокремлюють ушкоджені ділянки органів.
Кутинізація — це відкладання в клітинній оболонці жироподібної речовини кутину. Кутин звичайно відкладається на зовнішніх стінках клітин шкірки у вигляді плівки — кутикули. Кутикула непроникна для води і газів і тому захищає листки і молоді стебла від зайвого випаровування. Крім того, вона перешкоджає проникненню в рослину патогенних грибів та інших мікроорганізмів. Кутикула може мати різну товщину. Найтовстіша вона на шкірці листків рослин посушливих місць.
У деяких рослин до складу кутикули і кутикулярних шарів входить віск. Восковий наліт підсилює захисні властивості кутикули.
Ослизнення клітинних оболонок насіння сприяє закріпленню його у ґрунті й поглинанню з нього води, полегшує вихід зародка з насіння при проростанні. Слизуваті полісахариди містяться в клітинних стінках насіння, кореневих чохликів і волосків, листків деяких рослин посушливих місць. Слиз, що виділяється на поверхні
кореневого чохлика і кореневих волосків, допомагає кореню просуватися в ґрунті, контактувати з ґрунтовими частинками, поглинати з них воду і мінеральні речовини.
1.8. Поділ ядра і клітини
Мітоз — найпоширеніша універсальна форма поділу ядер як рослинних, так і тваринних клітин. У результаті мітозу дочірні клітини одержують ту саму кількість хромосом, яку мала материнська.
Життєвий цикл клітин здатної до поділу, який називають ще клітинним циклом, — це період її існування від одного поділу, у результаті якого вона виникла, до іншого, коли з неї утворюються дві дочірні. У ньому визначають два періоди: довгий період між поділами — інтерфазу і короткий період поділу ядра і клітини — мітоз.
Тривалість клітинного циклу в рослин залежить від типу клітин і тканин та дії факторів зовнішнього середовища (температура, умови мінерального живлення тощо). Однак інтерфаза завжди довша за мітоз. Вона складає від 8-10 годин до декількох діб, у той час як мітоз — від декількох хвилин до 2-3 годин.
Інтерфаза включає процеси підготовки до поділу ядра і клітини. У ній умовно визначають три періоди, які позначають G1, S і G2.
в1 - постмітотичний період - збільшується кількість цитоплазми і органоїдів, підготовка до поділу ДНК. S - синтетичний період, подвоюється ДНК. G2 - постсинтетичний, синтез РНК і білків клітини, структур, які беруть участь у мітозі.
Мітоз - це безперервний процес поділу ядра, за якого генетичний матеріал поділяється між ядрами дочірніх клітин. Мітоз умовно поділяють на чотири фази: профазу, метафазу, анафазу і телофазу.
Профаза — перша фаза мітозу. Ядро збільшується в розмірі. З ниток хроматину утворюються хромосоми. При цьому нитки спіра-лізуються, коротшають і ущільнюються, стають видимими під світловим мікроскопом. Хромосоми — нитчасті структури нуклеопро-теїнової природи. Кожна хромосома в профазі складається з двох хроматид, розташованих поруч і паралельно одна до одної. Хрома-тиди в хромосомі з'єднані в одному місці вузькою ділянкою — центромерою, що поділяє хромосому на два плеча різної довжини.
Навколо ядра збираються мікротрубочки. Спочатку вони розташовуються безладно, а потім організовуються в пучки біля полюсів ядра. Це — початкова стадія формування ахроматиноеого веретена
(веретена поділу). Ядерце поступово втрачає чіткість обрисів і зовсім зникає. Наприкінці профази ядерна оболонка теж зникає.
Метафаза характеризується повним розвитком ахроматинового веретена. Починається рух хромосом, що складаються з двох хро-матид. Вони переміщуються в екваторіальну частину клітини і прикріплюються центромерами до ниток напівверетена, які йдуть від протилежних полюсів. До кінця метафази хромосоми розташовуються на екваторі ахроматинового веретена і готові до поділу.
Анафаза починається з поділу центромери. Хроматиди цілком відокремлюються одна від одної, і тепер їх називають дочірніми хромосомами. Починається розтягнення дочірніх хромосом до протилежних полюсів клітини. У кінці анафази біля полюсів клітини збираються дві однакові групи дочірніх хромосом.
Телофаза — заключна фаза мітозу, в ході якої відбувається формування дочірніх ядер біля полюсів материнської клітини. Хромосоми деспіралізуються, гідратуються, набрякають і подовжуються. Поступово вони втрачають чіткі контури, зливаються в загальну масу й утворюють малопомітну тонку сітку хроматину. В ядрах з'являються ядерця. Одночасно навколо груп дочірніх хромосом виникає ядерна оболонка з елементів ЕПС.
Цитокінез — це поділ цитоплазми й утворення двох дочірніх клітин слідом за поділом ядра.
Мейоз — це особливий тип непрямого поділу ядра, що відбувається у всіх живих організмів, які розмножуються статевим шляхом. У мейозі відбувається редукція, тобто зменшення кількості хромосом удвічі. Тому мейоз називають ще редукційним поділом.
Мейоз складається з двох поділів ядра, які швидко йдуть один за одним. У першому поділі, який називають гетеротипним, відбувається редукція кількості хромосом, до полюсів клітини розходяться гомологічні хромосоми, в результаті чого дочірні ядра стають різними і гаплоїдними. У другому поділі, який називають гомеотип-ним і який дуже подібний до звичайного мітозу, утворюються однакові гаплоїдні ядра. Кожен з двох поділів мейозу умовно поділяють на ті ж чотири фази, які характерні для мітозу: профазу, метафазу, анафазу, телофазу.
У рослин мейоз відбувається при формуванні пилкових зерен і зародкового мішка. З нього починається гаметофітна фаза життєвого циклу рослини.
Контрольні запитання і завдання
1. Дайте визначення клітини. Чим рослинна клітина відрізняється від тваринної?
2. Що визначає форму й розміри рослинних клітин? Які клітини називають паренхімними, а які прозенхімними?
3. Розкажіть про склад рослинної клітини. Які органоїди належать до корпускулярних, а які до мембранних?
4. Які будова і функції плазмалеми і тонопласта?
5. Охарактеризуйте ендоплазматичну сітку. Які функції вона виконує?
6. Розкажіть про будову і значення плазмодесм.
7. Розкажіть про структуру, функції і походження хлоропластів.
8. Порівняйте три типи пластид.
9. Що ви знаєте про структуру і функції мітохондрій?
10. Яка роль лізосом і локального автолізу в рослинній клітині?
11. Що ви знаєте про апарат Гольджі і його роль у формуванні системи ендомембран?
12. Розкажіть про мікрочастинки рослинної клітини.
13. Які речовини відкладаються в запас у рослинних клітинах? Які форми їх відкладення?
14. Розкажіть про утворення і функції вакуолей.
15. Розкажіть про склад клітинного соку.
16. Розкажіть про склад клітинної оболонки та її функції.
17. Порівняйте склад і будову первинної і вторинної клітинної оболонки.
18. Що таке пори? Які типи пор ви знаєте?
19. Які ви знаєте фізико-хімічні видозміни клітинної оболонки? Яке значення вони мають для рослин?
20. Які існують типи поділу ядра? Яке їх значення?
21. У чому полягають особливості поділу рослинної клітини?