БОТАНІКА 3 ОСНОВАМИ ГІДРОБОТАНІКИ - 2010
3. АНАТОМІЯ РОСЛИН
3.1. Органи рослин
Орган — це частина багатоклітинного організму, яка виконує певну функцію або групу тісно пов’язаних функцій, має певну будову і складається із закономірно утвореного комплексу тканин. Всі органи організму взаємопов'язані і взаємодіють. Зв'язок між частинами організму, що розвиваються, визначається особливостями механізмів їхнього формування.
Вегетативні органи виконують функції підтримання індивідуального життя рослини і забезпечують її живлення та ріст, а також вегетативне розмноження. У водоростей тіло (талом, слань) не почленоване на органи, а представлене однією клітиною, нитками, колоніями або пластинками. У деяких із них помітно диференціювання талому на органи, зовні подібні до стебла і листка вищих рослин. Морфологічна й анатомічна будова вегетативних органів пристосована до виконання властивих їм функцій. У разі зміни функцій відповідно змінюються і вегетативні органи (метаморфоз). Вегетативними органами рослини є корінь і пагін.
Репродуктивні органи забезпечують спорове і статеве розмноження рослин. У спорових рослин є спори, спорангії і соруси, а генеративними органами у квіткових є квітка, плід і насінина.
Генеративні органи забезпечують розмноження рослин і безперервність їх існування на Землі.
Метаморфоз у рослин — видозмінення вегетативних органів (стебла, листка або кореня) внаслідок їхнього пристосування до виконання тих чи інших функцій у певних умовах існування. Метаморфоз відбувається в онтогенезі й полягає в зміні перебігу індивідуального розвитку органа, який виробився у процесі еволюції. Ці видозмінення можуть перетворити один орган на інший (здебільшого метаморфізуються не дорослі органи, а їхні зачатки) зі зміною форми і функції, зробити його зовні невпізнанним, і лише вивчення онтогенезу видозмінених органів дає можливість з'ясувати їх походження. Наприклад, внаслідок метаморфозу листка утворилися колючки у барбарису і кактусів, вусики у гороху і чини, ловильний апарат у комахоїдних рослин тощо. Термін «метаморфоз» запровадив у науку К. Лінней (1755).
Бульбоподібні потовщення на підземних пагонах є у стрілолиста
стрілолистого, частухи подорожникової та інших водних рослин. Тут накопичується крохмаль, цукри, білки та інші речовини.
На поверхні та в товщі води різних водних об’єктів можна побачити дрібненькі рослини - це представники родини ряскових (Lemnaceae): ряски мала (Lemna minor) і триборозенчаста (L. trisulca), спіродела багатокоренева (Spirodela polyrrhiza). У цих рослин тіло представлене видозміненим пагоном, який нагадує листочок і має назву фронда. Справжні листки у ряскових редуковані. Від нижнього боку фронди може відходити один (у ряски малої) або декілька (спіродела) корінців, які виконують роль якоря.
3.2. Будова і функції кореня
Це вегетативний (переважно підземний) орган з необмеженим ростом, яким рослина закріплюється в субстраті, що забезпечує її вертикальний ріст. Він поглинає воду й розчинені в ній мінеральні та частково органічні речовини і транспортує їх нагору в стебло. У зв'язку з цим довжина і розгалуження коренів значно перевершує надземні пагони. Наприклад, довжина коренів полину може досягати 3,5 м, а висота надземної частини — 120 см.
Корінь формується ще у зародку. Корінь, як і стебло, характеризується необмеженим ростом за рахунок меристем, розташованих у конусі наростання, має циліндричну форму і радіальне розташування тканин.
На відміну від стебла, корінь не несе листків, квітів і плодів. Він пов'язаний тільки зі стеблом, тому має простішу будову. На відміну від стебла, корінь розгалужується ендогенно, тобто зачатки бічних коренів закладаються у внутрішніх тканинах.
Функції кореня. Корінь виконує запасаючу функцію. У його паренхімі відкладаються в запас поживні речовини, іноді в значній кількості (коренеплоди).
У коренях із мінеральних речовин, які поглинаються з грунту й асимілятів, що надійшли з листків, синтезується ряд необхідних рослинам речовин (амінокислоти, фітогормони, алкалоїди та ін.).
Корінь може служити органом вегетативного розмноження.
Здійснює у ґрунті взаємодію з коренями інших рослин і вступає в симбіотичні взаємини з грибами і бактеріями.
Через корінь здійснюється зв'язок рослини з мікроорганізмами ґрунту. Корінь є місцем зберігання запасних речовин, слугує для
вегетативного розмноження. У багаторічних рослин корінь втягує у ґрунт основу пагонів з бруньками відновлення. Росте в довжину лише верхівковою меристемою, виявляючи позитивний геотропізм. Тільки в деяких рослин, що ростуть на заболочених ґрунтах, що майже не містять повітря, бічні корені можуть рости вгору. На коренях немає вузлів, меживузлів, бруньок (за винятком додаткових).
Шар ґрунту, в якому містяться корені рослин, називається ризосферою. Вона збагачена кореневими виділеннями, відмерлими кореневими волосками і є поживним середовищем для бактерій, грибів, що поселяються тут. Установлено, що в ризосфері бактерій у сотні й тисячі разів більше, ніж поза нею. Завдяки цьому ґрунтові процеси відбуваються в ній інтенсивніше. Представники мікрофлори ризосфери забезпечують рослини азотом, синтезують речовини, що стимулюють ріст, мінералізують органічні сполуки, деякі з них пригнічують рослини, виділяючи токсини, або паразитують на них.
Серед сучасних вищих рослин коренів не мають тільки мохоподібні, які поглинають воду за допомогою ризоїдів. Однак є ряд представників вищих рослин, що втратили корені у зв'язку з переходом до водного способу життя (пухирник, кушир, ряска триборо-зенчаста). Рослини, що виростають в сухому, жаркому кліматі, на сухих грунтах, мають могутньо розвинену кореневу систему. У болотяних рослин коренева система розвинена слабко.
Корені й кореневі системи розвиваються не лише в ґрунті. Деякі рослини (епіфіти, зокрема, тропічні орхідні) розвивають кореневу систему в повітрі, у товщі стебел інших рослин (видозмінені корені рослин паразитів і напівпаразитів — омели, повитиці тощо).
Корені використовують у різних галузях народного господарства: в медицині — як лікарську сировину (корені валеріани, женьшеню та ін.), в цукровому виробництві (коренеплоди цукрових буряків), у їжу (коренеплоди моркви, петрушки, редьки тощо), в тваринництві як кормові культури (кормові буряки, турнепс тощо), в різних галузях промисловості: для виробництва каучуку (корені кок-сагизу, тау-сагизу, крим-сагизу тощо), рослинних барвників (корені марени красильної, кермеку тощо).
Види коренів. Типи кореневих систем. За походженням розрізняють головний, бічні й додаткові корені.
Головний корінь (вісь першого порядку) утворюється тільки із зародкового корінця насінини.
Додаткові корені беруть початок від зони кущіння зародка, від стебла і листків або їхніх видозмін. Від головного й додаткових коренів відходять бічні корені — осі другого й наступних порядків галуження. При вирощуванні рослин для збільшення маси додаткових коренів у поверхневих родючих шарах ґрунту на приземні ділянки стебел нагортають купи землі. Щоб збільшити масу бічних коренів, проводять пікірування проростків (переважно при вирощуванні культурних рослин розсадою).
Головний, бічні й додаткові корені разом становлять кореневу систему. Розрізняють стрижневу, мичкувату та змішану кореневі системи. Стрижневу кореневу систему має більшість представників двосім'ядольних рослин (люцерна, соя, горох, соняшник тощо).
Мичкувата коренева система характерна для рослин класу односім'ядольних (жито, пшениця, овес, кукурудза та ін.). У цих рослин головного кореня немає (а якщо він зберігся, то за розмірами не вирізняється серед додаткових), і система представлена мичкою додаткових коренів, що утворилися від вузла кущіння. Змішана коренева система характерна здебільшого для представників трав'янистих двосім'ядольних рослин (суниці, полуниці), в яких відносно добре розвинені усі три типи коренів.
За формою корені дуже різноманітні: ниткоподібні, конусоподібні, веретеноподібні, бульбоподібні тощо (рис. 3.1).
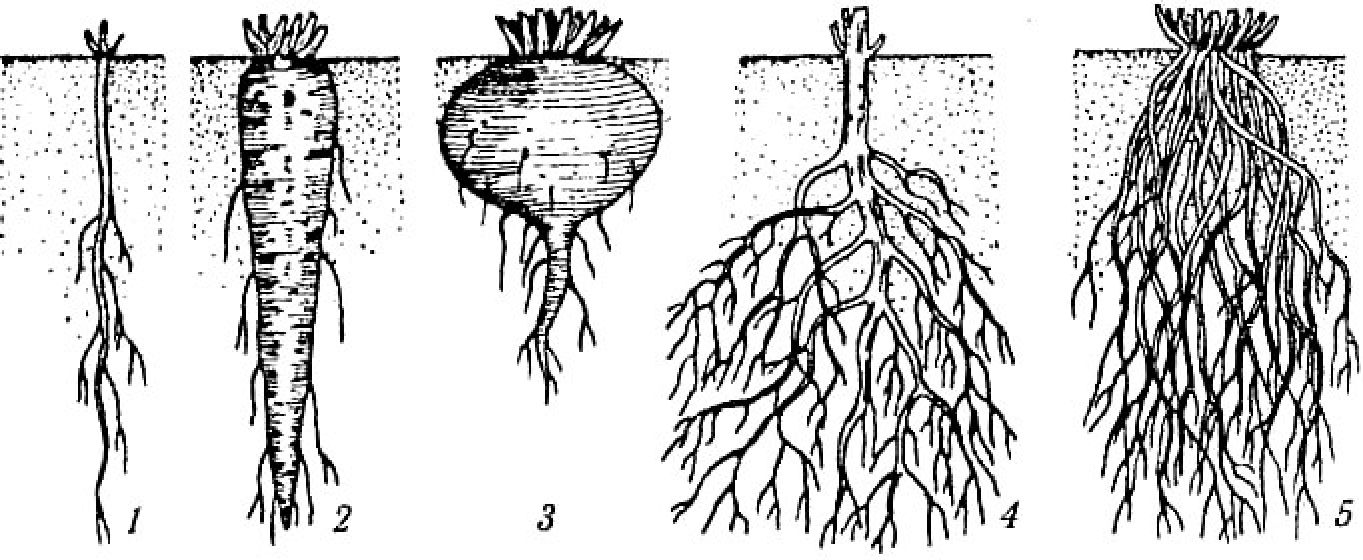
Рис. 3.1. Форми коренів:
1 — ниткоподібний; 2 — веретеноподібний; 3 — ріпчастий (2, 3 — запасальні); 4 — стрижнеподібний (типовий); 5 — мичкуватий
Корені рослин проникають у ґрунт на велику глибину, досягаючи ґрунтових вод. Наприклад, головний корінь верблюжої колючки
(рослина посушливих напівпустель з родини бобових) досягає глибини 20 м, головний корінь дворічного дуба — 2 м. На глибину 2 м проникають кореневі системи злакових рослин, а трав'янистих бобових — на 10 м і більше. Загальна довжина всіх коренів дуже велика. Коренева система рослин значно краще розвинена, ніж надземна частина. Наприклад, корені будяку заглиблюються більш як на 6 м за висоти надземної частини 1 м. За звичайних умов вирощування хлібні злаки (пшениця, овес, ячмінь) мають кореневу систему загальною довжиною 20 км, гарбузи — близько 25 км, а загальна довжина кореневої системи жита, яке вирощували в дослідних умовах, сягала 63,2 км. Діаметр кореневої системи плодових дерев у 2—5 разів більший за діаметр крони.
Корінь наростає своєю верхівкою. При обриві кореня спостерігається могутній розвиток бічного коріння. Ця особливість кореня використовується при пікіруванні. При пікіруванні пересаджують рослину і вкорочують головний корінь на 1/2—1/3 його довжини, що викликає рясне розгалуження частини, що залишилася, забезпечуючи розростання кореневої системи.
Зони кореня. На поздовжньому зрізі молодого кореня можна бачити декілька зон, які добре виявляються (рис. 3.2).
Зона поділу клітин знаходиться на самому кінчику кореня. Вона складається з меристе-матичної тканини і являє собою конус наростання кореня. Розміри її дуже малі — до 1 мм.
Зона поділу вкрита кореневим чохликом, який захищає ніжні меристематичні клітини від ушкодження твердими частинками ґрунту.
Рис. 3.2. Зони кореня:
1 — кореневий чохлик; 2 — зона поділу; 3 — зона росту; 4 — зона всмоктування; 5 — зона проведення; 6 — кореневі волоски; 7 — зачатки бічних коренів
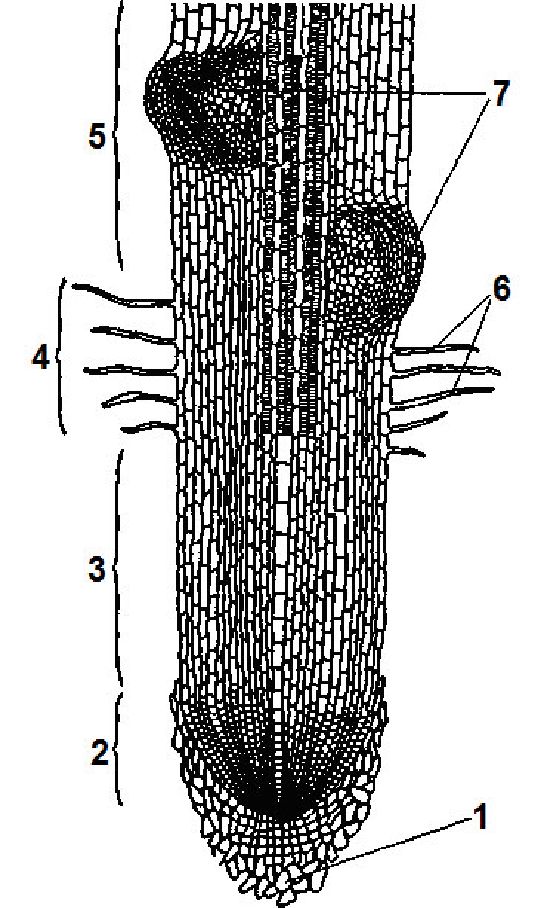
Кореневий чохлик являє собою ковпачок із паренхімних клітин. Він утворюється зовнішнім шаром клітин апікальної меристеми. Тривалість життя його клітин 2—9 діб, залежно від фізичного стану ґрунту і виду рослин. Крім захисту меристематичних клітин кореневий чохлик сприяє проникненню кореня в глибину. Він виділяє слиз, яким обволікається корінь, що полегшує його просування між частинками ґрунту, а також забезпечує реакцію кореня на гравітацію (позитивний геотропізм), яка визначає напрямок росту кореня.
Кореневий чохлик є у всіх сухопутних рослин. У водних рослин при рості їх коріння у воді чохлик не утворюється. При вкоріненні їх в грунті на кінці коріння розвивається чохлик.
Зона росту, або витягування, розташовується над зоною поділу і має довжину 2—5 мм. Тут клітини не діляться, а ростуть. їхні оболонки розтягуються, у клітинах з'являються вакуолі, спостерігається диференціація клітин у постійні тканини кореня. Тут формуються епіблема, первинна кора та центральний циліндр.
Зона всмоктування, або вбирання, має диференційовані тканини. Тут можна розглядати первинну будову кореня. Тому її ще називають зоною первинної будови. Покривна тканина на цій ділянці кореня утворює численні волоски, що забезпечують інтенсивне поглинання води і мінеральних речовин з ґрунту.
Довжина зони поглинання — від одного до декількох сантиметрів. На 1 мм2 площі поверхні кінчика кореня утворюється близько 400 волосків, залежно від виду рослини, умов зволоження, аерації й температури ґрунту. До прикладу, загальна кількість кореневих волосків у трав'янистих рослин може сягати кількох мільярдів. У нижній частині зони кореневі волоски формуються, потім функціонують, а у верхній частині руйнуються і відмирають. Тривалість життя кореневих волосків у середньому складає 10—20 днів. Тому зона всмоктування ніби переміщується протягом усього періоду росту кореня вниз і завжди перебуває поблизу кінчика кореня. Кореневими волосками верхівка кореня закріплюється в ґрунті, отже, вони сприяють її просуванню між частинками ґрунту.
Немає кореневих волосків лише у водних і болотних рослин. Корені цієї зони мають первинну анатомічну будову.
Зона проведення. Вище поглинаючої зони після відмирання кореневих волосків покривну функцію починають виконувати верхні
шари первинної кори, стінки клітин якої корковіють. У цій частині здійснюється транспорт речовин із кореня в стебло і листки. Крім того, тут закладаються і формуються бічні корені, тому цю зону називають ще зоною розгалуження. У дводольних у провідній зоні формується вторинна будова кореня. Провідна зона (зона укріплення бічних коренів) найдовша. Вона збільшується завдяки утворенню бічних коренів і розміщена між зоною всмоктування та кореневою шийкою. Ця зона виконує функцію укріплення рослини в ґрунті та надходження розчинів мінеральних речовин до наземних органів рослини, а органічних — до клітин кореня.
Особливості анатомічної будови кореня. Для кореня характерне радіальне розміщення провідних тканин. Бічні корені формуються із перициклу, тобто мають ендогенне походження.
Диференціація тканин кореня відбувається у зоні всмоктування. За походженням це первинні тканини, бо утворюються з первинної меристеми зони поділу. Тому мікроскопічна будова кореня в цій зоні називається первинною (рис.3.3). Тут розрізняють центральний (осьовий) циліндр і первинну кору, вкриту одним рядом клітин з кореневими волосками — епіблемою (ризодермою). Зовнішній шар центрального циліндра — перицикл складається з одного (рідше кількох) ряду живих паренхімних клітин. З нього утворюються бічні корені (тому перицикл називають коренерідним шаром), а при переході кореня до вторинної будови формуються вторинні твірні тканини: камбій і корковий камбій — фелоген.
Первинна кора зазвичай досить добре розвинена. На її частку припадає основна маса первинних тканин кореня. В ній розрізняють зовнішню (ектодерма), середню (мезодерма, корова паренхіма) і внутрішню (ендодерма) частини. Клітини екзодерми живі, тісно зімкнені, з потовщеними оболонками. Коли ризодерма з кореневими волосками відмирає, розміщені під нею клітини екзодерми злегка корковіють і до утворення вторинної покривної тканини виконують захисну й частково — механічну функції.
Глибокі шари кореня складаються з дрібніших клітин. Самий внутрішній шар кори утворює так звану ендодерму, що відокремлює кору від середньої частини кореня - центрального циліндра.
Центральний циліндр (стела) займає центральну частину кореня і складається із первинної твірної тканини — перициклу, судинно-волокнистого пучка, клітин основної і механічної тканин.
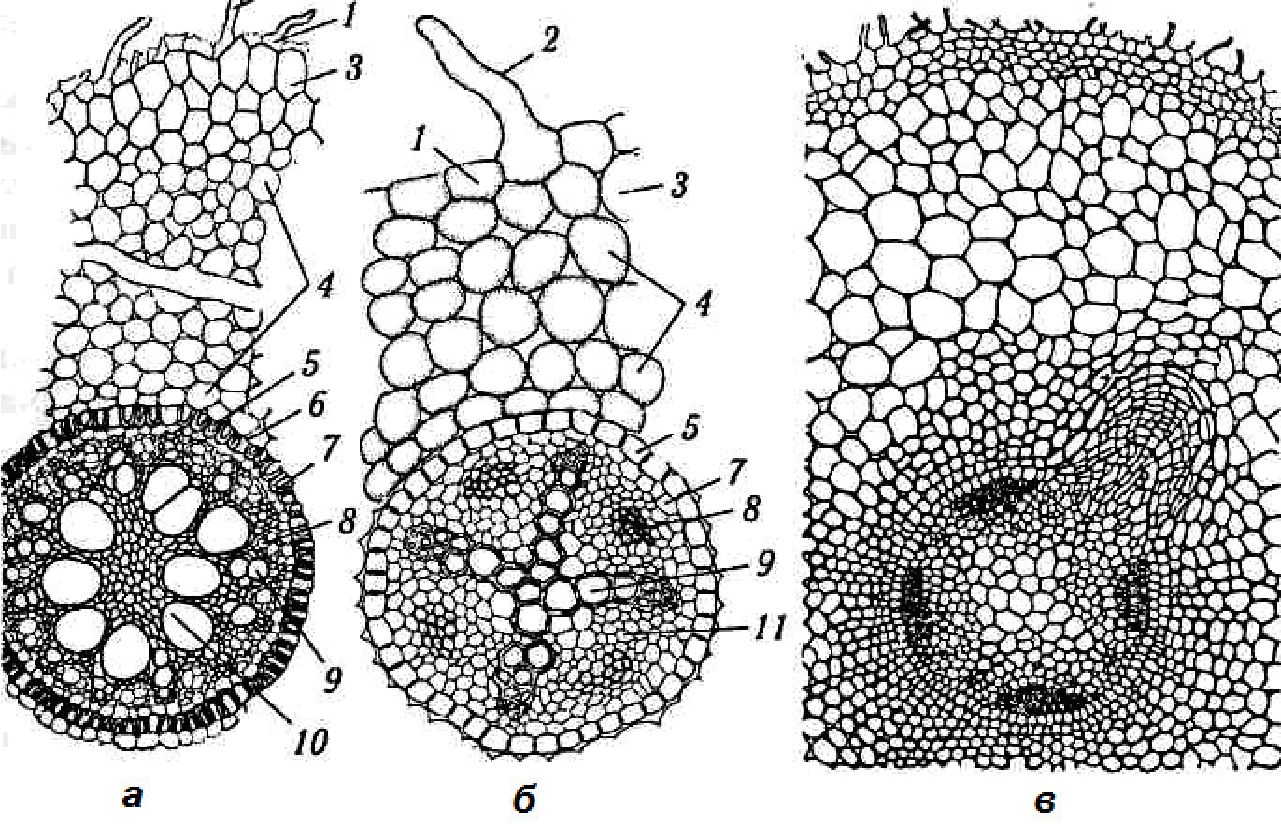
Рис. 3.3. Анатомічна будова кореня:
а — одно- і б — двосім'ядольних рослин; в — закладання бічного кореня: 1 — епіблема; 2 — кореневий волосок; 3 — екзодерма; 4 — мезодерма (корова паренхіма); 5 — ендодерма; 6 — пропускна клітина (3—6 — первинна кора); 7 — пери-цикл; 8,9 — первинна флоема та ксилема; 10 — склеренхіма; 11 — основна паренхіма (7—11 — центральний циліндр)
У судинно-волокнистому пучку чергуються ділянки ксилеми й флоеми. У корені ксилема розвинена краще, ніж флоема. Вона досягає центра кореня і розходиться від нього променями (у двосім'ядольних рослин їх 2—5, в односім'ядольних більш як 5). Флоема займає невеликі ділянки між променями ксилеми. На відміну від стебла, в корені зазвичай немає розвиненої серцевини.
В односім'ядольних і папоротей первинна анатомічна будова кореня зберігається упродовж усього життя. В голонасінних і покритонасінних двосім'ядольних у зоні закріплення будова кореня змінюється, що зумовлено утворенням вторинних твірних тканин камбію і коркового камбію.
Первинна будова кореня. У сформованій первинній будові кореня, як і в стеблі, розрізняють покривну тканину, первинну кору і центральний циліндр. Однак у корені межа між корою і центральним циліндром виявляється більш чітко, починаючи ще з меристе-матичної зони.
Покривна тканина кореня — ризодерма, або епіблема, або волос-
коносний шар, утворює кореневі волоски (рис. 3.4).
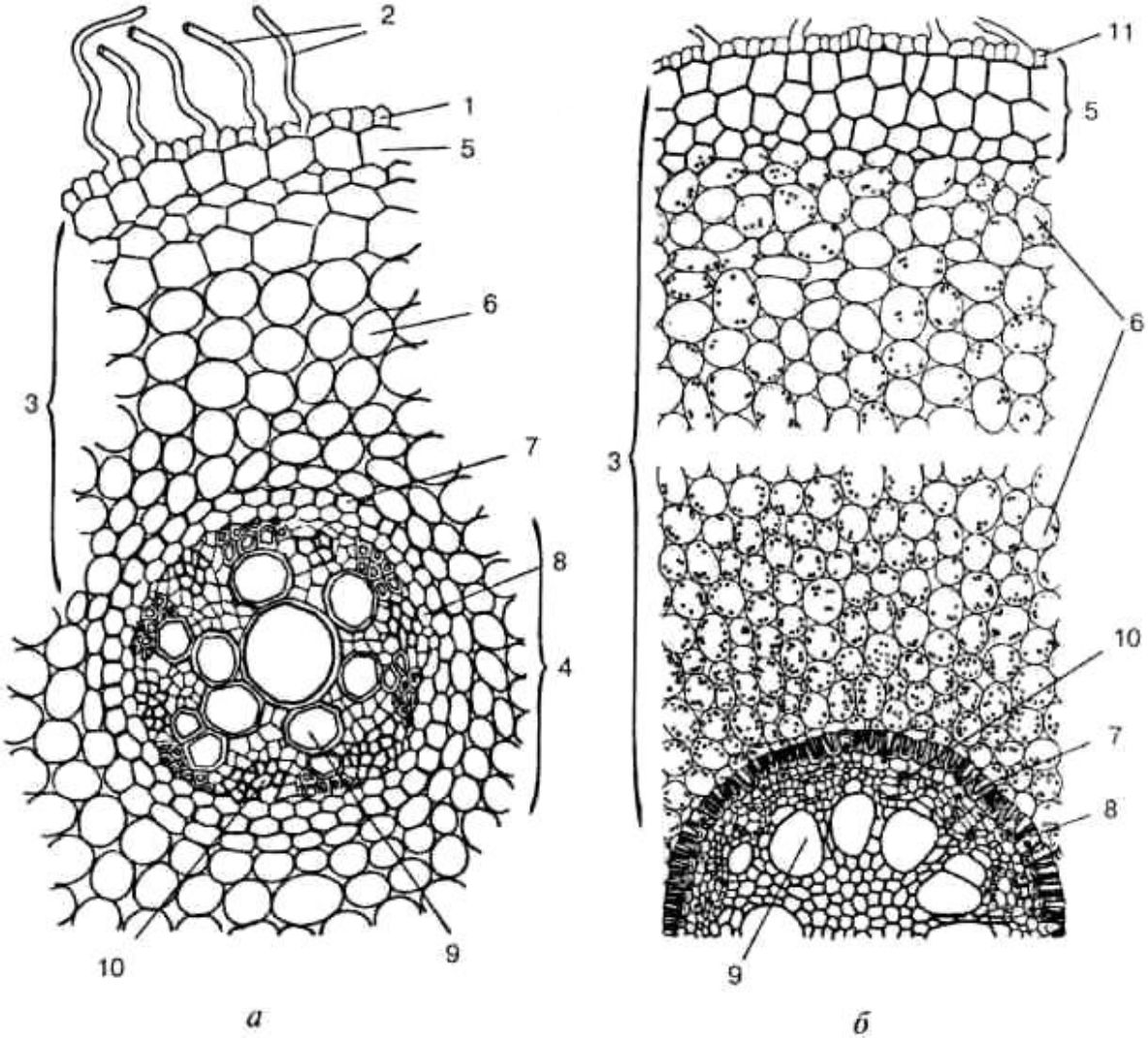
Рис. 3. 4. Поперечний зріз кореня півників:
а — у зоні всмоктування; б — у зоні проведення; 1 — ризодерма; 2 — кореневі волоски; 3 — первинна кора; 4 — центральний циліндр; 5 — екзодерма; 6— мезодерма; 7 — ендодерма; 8 — перицикл; 9 — судини ксилеми; 10 — флоема; 11 — залишки ризодерми після злущування кореневих волосків
Ризодерма першою починає диференціюватися в нижній частині зони поглинання, що дуже важливо для розвитку кореня і всієї рослини, тому що волоски одразу починають поглинати воду і мінеральні речовини, подаючи їх у провідну систему кореня. Кореневі волоски розвиваються досить швидко — протягом 30-40 годин. Довжина їх у середньому від 0,15 до 8 мм, але можуть досягати і 1 см. Причому кореневі волоски в трав'янистих рослин довші, ніж у деревних. Кількість волосків у перерахунку на 1 мм2 залежить від виду рослин і вологості ґрунту. Найбільша щільність кореневих волосків у рослин, що ростуть на досить зволожених ґрунтах. У водних рослин волосків мало або вони зовсім відсутні. У таких рослин поглинаюча поверхня зростає за рахунок розгалуження коренів.
Кореневі волоски створюють величезну поглинаючу поверхню, забезпечуючи інтенсивне усмоктування води і мінеральних речовин. Довжина волосків однієї рослини може досягати декількох десятків кілометрів. Наприклад, у пшениці, яка вирощена за оптимальних умов, довжина всіх кореневих волосків склала 10 000 км, загальна їх площа — 400 м2. Крім поглинаючої, кореневі волоски виконують закріплювальну функцію.
Як відомо, кореневі волоски недовговічні. Вони злущуються від тертя об тверді частинки ґрунту і відмирають разом із клітинами епіблеми, що їх утворили. Це відбувається звичайно у верхній частині зони поглинання.
Первинна кора — багатошарове утворення з паренхімних клітин. У коренях вона товстіша, ніж у стеблах. Причому її діаметр в однодольних більший, ніж у дводольних. У первинній корі є три шари: екзодерма, мезодерма, або корова паренхіма, і ендодерма.
Екзодерма — зовнішня частина первинної кори, що складається з одного або декількох шарів щільно зімкнутих клітин, часто багатогранної форми, іноді з потовщеними стінками. Під ризодермою вона виконує провідну функцію, транспортуючи воду з розчиненими мінеральними речовинами від волосків далі по первинній корі. Коли ж волосконосний шар відмирає, екзодерма стає покривною тканиною. Стінки її клітин корковіють, і вона захищає корінь від механічних впливів і проникнення мікроорганізмів.
Серед мертвих окорковілих клітин зберігається деяка кількість живих. Це пропускні клітини, які після втрати кореневих волосків забезпечують у невеликих розмірах поглинаючу функцію в цій частині кореня. В однодольних екзодерма має кілька шарів клітин, а в дводольних — частіше один. У деяких рослин екзодерми немає.
За екзодермою розміщується корова паренхіма — мезодерма. Вона могутня, особливо в однодольних. Клітини в ній розташовані пухко зі значною кількістю міжклітинників. У коровій паренхімі можуть знаходитися різні вмістища, а іноді й групи механічних волокон. Мезодерма здійснює транспорт речовин у радіальному напрямку. У ній можуть відкладатися запасні речовини, а також синтезується ряд речовин, необхідних рослині.
Закінчується первинна кора одношаровою ендодермою, яка оточує центральний циліндр. Спочатку вона складається з живих тонкостінних, чотирикутних на поперечному зрізі клітин. Її основна функція — регуляція транспорту речовин і води з кори в судини центрального циліндра. У своєму розвитку ендодерма проходить три стадії, що підсилюють її регуляторну функцію.
На першій стадії відбуваються зміни оболонок в екваторіальній частині її клітин. Клітинні оболонки дещо потовщуються і просочуються суберином — корковіють. Утворюються потовщені непроникні для води пояски, які оточують клітину по екватору - пояски-Каспарі. Пояски сусідніх клітин стикаються.
Вода і розчинені в ній речовини, що рухаються по апопласту первинної кори, можуть дійти тільки до поясків, а далі, щоб потрапити в центральний циліндр, повинні перейти в симпласт, тобто в цитоплазму клітин ендодерми через плазмалему.
У дводольних при формуванні вторинної будови кореня первинна кора відмирає і злущується. Тому в тих рослин, у яких вторинні зміни починаються рано, диференціювання ендодерми закінчується на першому етапі. При пізніших змінах у ендодермі деяких дводольних відбувається другий етап диференціювання. Він полягає в тому, що на всіх клітинних стінках відкладається шар, що складається із суберину і целюлози. Це робить клітинні оболонки непроникними. Однак не всі клітини піддаються таким змінам. Залишаються пропускні клітини, розташовані проти променів ксилеми. Вони мають тільки пояски Каспарі й продовжують здійснювати транспорт речовин із первинної кори в центральний циліндр.
В однодольних рослин, у яких відсутні вторинні зміни і первинна кора в корені зберігається впродовж усього їх життя, здійснюється третя стадія змін в ендодермі. Радіальні та внутрішні стінки її клітин потовщуються і дерев'яніють. Здерев'янінню піддаються і тонкі зовнішні оболонки. У результаті клітини ендодерми набувають підковоподібної форми. Протопласти клітин відмирають, і ендодерма виконує механічну функцію. Для транспорту речовин із кори в центральний циліндр в ендодермі залишаються пропускні клітини з поясками Каспарі і живим протопластом, розташовані напроти судин ксилеми.
У цілому первинна кора кореня виконує ряд важливих функцій.
- По ній здійснюється радіальний транспорт води і мінеральних речовин від кореневих волосків до ксилеми й асимілятів від флоеми до кореневих волосків.
- Після відмирання ризодерми первинна кора (екзодерма) функ
ціонує як покривна тканина, а поглинаюча функція частково здійснюється пропускними клітинами.
- У паренхімі первинної кори можуть відкладатися в запас поживні речовини. У її клітинах первинної кори (мезодерми) синтезується ряд важливих для рослин сполук (алкалоїди, глікозиди).
- У первинну кору проникають гіфи грибів, утворюючи мікоризу. Центральний циліндр, або стела, кореня складається з перициклу і складного радіального провідного пучка.
Перицикл є зовнішнім шаром, що оточує центральний циліндр. Він складається з дрібних живих тонкостінних клітин. Частіше перицикл одношаровий, що скорочує шлях речовин від кореневих волосків через первинну кору до судин ксилеми, але є і багатошаровий. Наприклад, у голонасінних (гінкго, саговники), у деяких квіткових (агава, драцена, волоський горіх, хміль). У ряду рослин кільце перициклу переривається променями ксилеми, які стикаються безпосередньо з ендодермою і поділяють його на сегменти (злаки), що також скорочує шлях транспорту речовин у корені до трахеаль-них елементів. Іноді в перициклі, звичайно проти променів ксилеми, утворюються вмістища виділень, що також робить його переривчастим (зонтичні). У коренях водних рослин перицикл відсутній.
Перициклу в коренях належить дуже важлива роль. Назвемо його основні функції:
- у перициклі закладаються бічні корені;
- він бере участь в утворенні камбію в коренях дводольних;
- з нього в корені виникає фелоген;
- із перициклу формуються додаткові камбіальні кільця;
- у перициклі закладаються додаткові бруньки;
- у перициклі утворюються різні вмістища.
Центральну частину стели кореня займає провідний пучок, що складається з радіальних променів ксилеми і флоеми, які чергуються. Як відомо, у радіальному пучку кількість променів ксилеми і флоеми є однаковою. Кількість променів у провідній тканині в пучках може бути різною залежно від виду рослин. Так, у буряку їх два, у бобових і гарбузових — чотири, в ірисів — багато. Кількість променів в однієї рослини в різних її коренях може змінюватися внаслідок їх редукції. Так, у бобових в одних корінцях виявляється чотири тяжі провідних тканин, в інших — три.
Провідний пучок у корені утворюється з прокамбію. Першою з усіх його елементів на периферії прокамбіального тяжу диференціюється протофлоема, необхідна для подачі до точки росту кореня асимілятів. Потім формується протоксилема. Оскільки ксилема і флоема — складні тканини, вони містять певну кількість паренхіми, а іноді і механічні волокна.
Як правило, в корені серцевини немає. На її місці звичайно знаходяться судини ксилеми. Однак у деяких рослин у центрі кореня можна спостерігати паренхімні клітини або механічну тканину (кукурудза, іриси). Але це не справжня серцевина, тому що її клітини утворюються з прокамбію, а не з основної меристеми конусу наростання, як це відбувається в стеблі.
Закладення і розвиток бічних коренів. У більшості рослин бічні корені закладаються в перициклі в провідній зоні. Це має певне біологічне значення для рослин. При утворенні кореневих зачатків на коренях виникають горбики (рис. 3.5). Якби це відбувалося в зоні поглинання, то шлях речовин від кореневих волосків до центрального циліндра подовжувався б. Закладення ж бічних корінців у провідній зоні не заважає процесу поглинання і радіальному транспорту речовин. Ендогенне розгалуження коренів захищає їх зачатки на початкових етапах розвитку від зіткнення з ґрунтом.
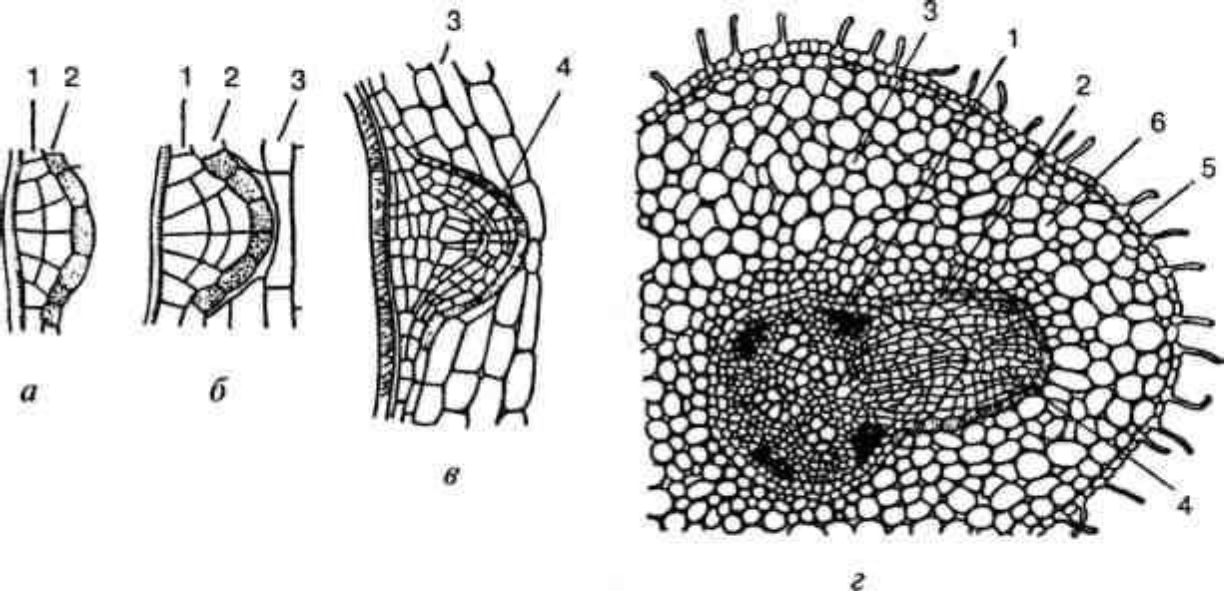
Рис. 3.5. Закладення і розвиток бічного кореня:
а, б, в — послідовні етапи розвитку бічного корінця; г — зачаток бічного корінця на поперечному зрізі кореня соняшника; 1 — перицикл; 2 — ендодерма; 3 — паренхіма первинної кори; 4 — конус наростання зачаткового корінця; 5 — епіблема; 6 — первинна кора
У кореневому зачатку формуються всі анатомічні структури і зв'язуються з відповідними структурами головного кореня. Цьому сприяє перицикл, клітини якого діляться і диференціюються в елементи провідних тканин і паренхіми і зв'язують провідні системи обох коренів. Кореневий чохлик також формується на кінчику зачатка і після розриву кишеньки починає виконувати свої функції.
Не всі кореневі зачатки виходять назовні, досягають повного розвитку і перетворюються в бічні корені. Деякі з них відмирають, а інші затримуються у своєму розвитку і стають сплячими. Останні можуть продовжувати ріст ще тривалий час.
Вторинна будова кореня. У папоротей і однодольних первинна будова зберігається в коренів упродовж усього життя. У голонасінних і дводольних вона змінюється вторинною будовою, яку можна спостерігати в провідній зоні.
Спочатку в коренях дводольних камбію немає, він виникає перед вторинними змінами і формує їх. Камбій закладається дугами під променями первинної флоеми, утворюється з розташованої там паренхіми. Оскільки центр кореня займає первинна ксилема, камбіальна дуга опиняється між ксилемою і флоемою. Далі камбіальні дуги подовжуються уздовж променів ксилеми, доходячи до перицик-лу, клітини якого також починають ділитися, утворюючи камбій.
Камбіальні клітини починають працювати в дугах під променями флоеми ще до замикання повного кільця. Звичайно, вторинної деревини утворюється більше, ніж лубу, і камбіальне кільце поступово випрямляється. Зрозуміло, що воно неоднорідне за походженням: одні його ділянки виникли з паренхіми провідного пучка, інші — з перициклу. Функціонують ці ділянки також по-різному: похідні паренхіми утворюють елементи вторинного лубу і вторинної деревини, а похідні перициклу — паренхімну тканину. Іншими словами, меристема, утворена з паренхіми, працює як пучковий камбій, а утворена з перициклу — як міжпучковий.
У центрі кореня залишається первинна ксилема. Тяжі паренхіми йдуть від променів первинної ксилеми, розсікаючи вторинну деревину і вторинний луб. Вони аналогічні серцевинним променям стебла, але, оскільки в коренях немає серцевини, тут їх називають па-ренхімними променями (рис. 3.6).
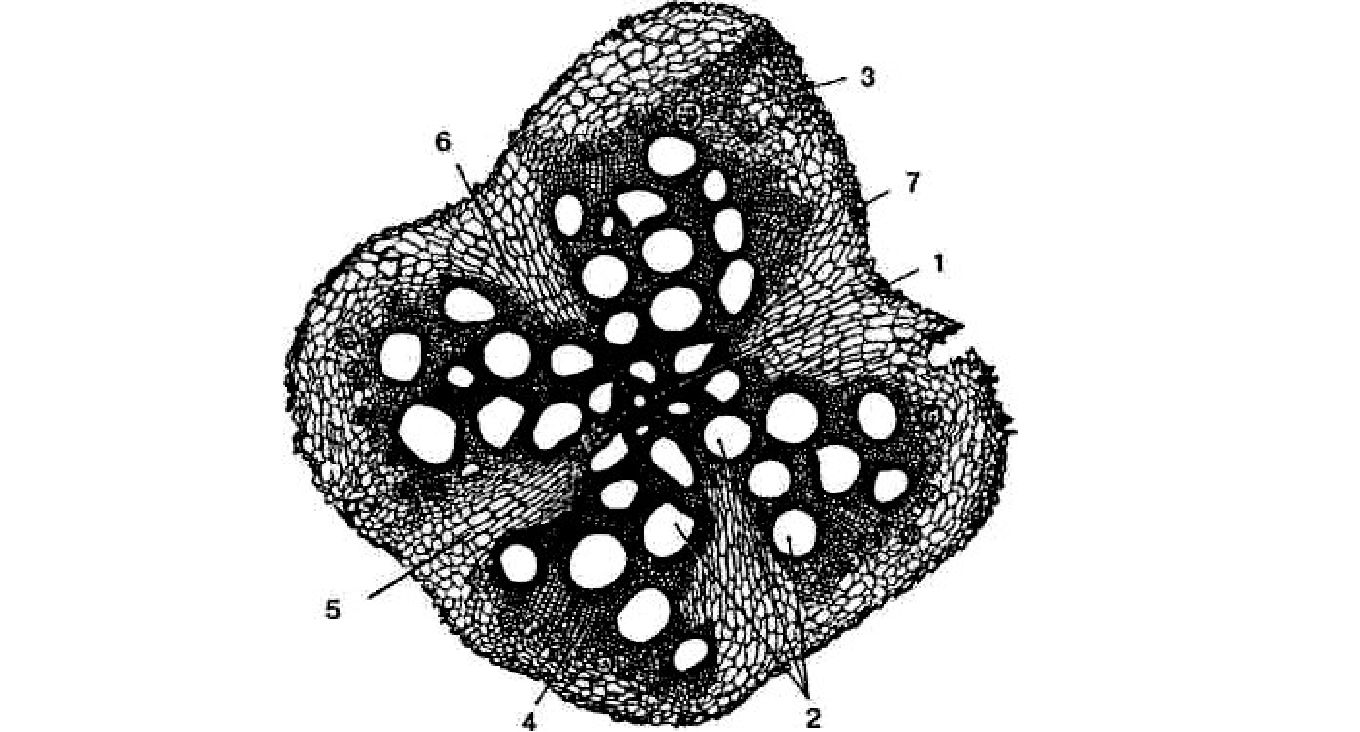
Рис. 3.6. Вторинна будова кореня гарбуза:
1 — первинна ксилема; 2 — вторинна ксилема; 3 — вторинна флоема; 4 — пучковий камбій; 5 — міжпучковий камбій; 6 — паренхімні промені; 7 — перидерм
Первинна флоема при формуванні вторинної структури кореня відтискується вторинним лубом далеко на периферію, де втрачає свою функцію і сплющується. У коренях унаслідок сезонних змін утворюються річні кільця, але вони більш вузькі, ніж у стеблах, і межі між ними виражені слабко.
У формуванні вторинної будови кореня бере участь не тільки камбій, що утворює луб і деревину, але і корковий камбій.
Вторинні зміни в коренях однодольних. Корені однодольних, як і їх стебла, зберігають первинну будову протягом усього життя. При цьому з віком відбувається склерифікація паренхіми, що підсилює механічну міцність рослин. Однак у ряді деревних однодольних, у яких відбувається потовщення стебла, росте в товщину і корінь. У корені, як і в стеблі, це відбувається за рахунок роботи додаткових камбіальних кілець, які формуються частіше з перициклу, рідше — з паренхімних клітин первинної кори (драцена, юка). При цьому перше додаткове камбіальне кільце доцентрово відкладає багато паренхіми і закриті провідні пучки, а назовні — нове камбіальне кільце, що працює в такому ж режимі. Зверху такий корінь покривається перидермою, а іноді пізніше формується кірка.
Метаморфози коренів. Видозміни, або метаморфози, коренів відбуваються звичайно у зв'язку з пристосуванням рослин до умов навколишнього середовища або з посиленням будь-якої однієї з його функцій, наприклад запасаючої. При цьому відбуваються значні зміни в структурі кореня.
Потовщуватися для запасання можуть головний, бічні й додаткові корені. При потовщенні головного кореня утворюються коренеплоди (морква, буряк); при потовщенні бічних і додаткових коренів — кореневі бульби (жоржини).
Коренеплоди залежно від місця розвитку і походження запасаючої паренхіми поділяють на три групи.
Запасаюча паренхіма в основному зосереджена у вторинному лубі. У таких рослин у коренях діаметр лубу великий і перевищує вторинну деревину (морква, петрушка).
Запасаюча паренхіма знаходиться у вторинній деревині, в той час як луб представлений тонким шаром (редиска, редька, ріпа).
Запасаюча тканина утворюється в результаті діяльності додаткових камбіальних кілець. Така будова притаманна коренеплодам буряка.
У деяких рослин корені потовщуються за рахунок розвитку паренхіми в первинній корі, а іноді в первинній корі і серцевині. У багатьох болотяних рослин корені потовщені, але це пов'язано не з відкладенням поживних речовин, а з розвитком у первинній корі аеренхіми.
Повітряні корені утворюються в деяких тропічних рослин (орхідні, ароїдні). Звичайно такі корені притаманні епіфітам — рослинам, що використовують стовбури і гілки дерев для прикріплення. їхні корені вільно висять у повітрі. Зверху повітряні корені вкриті багатошаровою покривною тканиною веламеном, яка захищає їх від висихання і перегріву. Він складається з мертвих клітин. Повітряні корені за допомогою веламену поглинають воду з повітря, але не осмотично, як звичайні корені, а капілярним шляхом.
Симбіоз коренів рослин. У коренів рослин зустрічаються різні види симбіозу. Так, у бобових на коренях утворюються бульбочки. Це прояв симбіозу коренів із азотфіксуючими бактеріями, які через кореневі волоски проникають у паренхіму первинної кори і викликають там активний поділ клітин. Це сприяє утворенню на коренях здуття — бульбочок. Бактерії потрапляють в клітини і утворюють там бактероїди і бактероїдну тканину.
Бульбочкові бактерії здатні засвоювати атмосферний азот тільки
при взаємодії з рослиною і використанні її органічних речовин. Фіксований азот включається в амінокислоти рослини.
Робота бульбочкових бактерій ефективна. Бактерії-симбіонти витрачають на фіксацію 1 г атмосферного азоту всього 3—4 г вуглеводів, тоді як вільноіснуючі азотфіксатори використовують на засвоєння тієї ж кількості азоту 50—100 г вуглеводів.
Бульбочкові бактерії забезпечують азотом рослини навіть на грунтах, бідних його доступними формами. Після відмирання бобових рослин або їх частин грунт збагачується доступним азотом. Серед представників родини бобових лише 10% видів мають бульбочки, і вони відрізняються за інтенсивністю фіксації азоту.
Бульбочки з азотфіксуючими бактеріями зустрічаються не тільки на коренях бобових, але і деяких інших рослин (вільха, обліпиха).
Мікориза — симбіоз грибів із коренями рослин. Звичайно вона виникає в зоні поглинання. Гіфи гриба можуть обплітати корінь зовні — ектомікориза, або проникати в клітини кори кореня — ендо-мікориза. Іноді поєднуються обидва типи мікоризи (рис.3.7).
Корені з ектомікоризою не мають волосків, їх заміняють гіфи гриба, що залишаються назовні та виглядають як кореневі волоски. Мікориза затримує ріст коренів, але на кінчику головного кореня вона не утворюється, і він продовжує нормально рости в довжину.
Гіфи ендомікоризних грибів розвиваються усередині клітин кори, а іноді і ризодерми, але кореневі волоски при цьому утворюються і функціонують нормально. Співіснуючи з рослиною, гриби одержують від неї органічні речовини, утворені в процесі фотосинтезу.
Рослини також мають користь від симбіозу з грибами. Гіфи гриба утворюють велику поглинаючу поверхню і забезпечують рослину мінеральними речовинами.
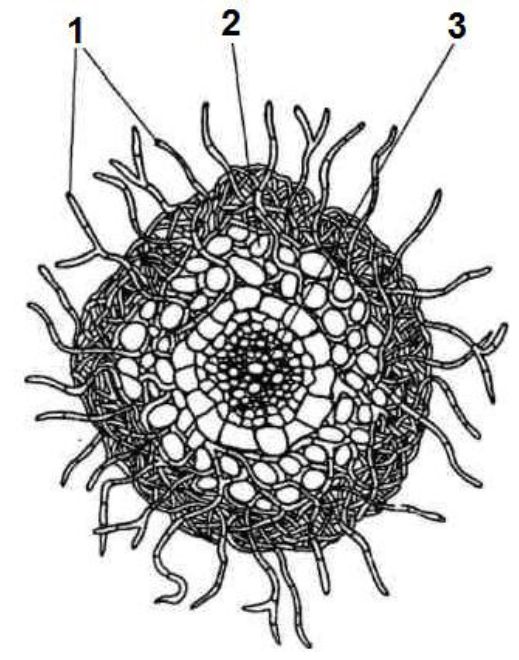
Рис. 3.7. Мікориза на коренях дуба: 1
— гіфи гриба; 2 — міцелій гриба у первинній корі кореня; 3 — центральний циліндр кореня
Гриби, як сапротрофи, розкладають органічні залишки і мінералізують їх, поліпшуючи постачання рослин мінеральними елементами і насамперед азотом і фосфором. Інакше кажучи,мікоризні гриби покращують мінеральне живлення рослин.
Гриби сприяють підвищенню концентрації клітинного соку рослин за рахунок поглинання мінеральних речовин з ґрунту і виділяючи в рослинну клітину гідролітичні ферменти, підсилюють розщеплення крохмалю до цукрів, частина яких надходить у вакуолю. У результаті зростає осмотичний тиск і сисна сила клітин кореня, а отже, і поглинання ним води, тобто мікориза поліпшує водопостачання рослин.
Мікориза захищає корені рослин від проникнення патогенних організмів, виробляючи антибіотичні речовини.
Завдяки користі, що рослини мають від симбіозу з грибами, мікориза широко розповсюдилася в природі. Понад 70 % голонасінних і покритонасінних і 60 % вищих спорових рослин мають мікоризу. Не утворюють її рослини деяких родин (хрестоцвіті, осокові, макові, кропивові тощо), відсутня вона й у мохів.
Кожен вид рослин пристосований до співіснування з певним видом гриба. Відсутність мікоризи в багатьох рослин гальмує їх ріст і розвиток.
Кореневе живлення рослин. Живлення рослин полягає у поглинанні ними із навколишнього середовища речовин, потрібних для процесів життєдіяльності, а також розподіл та використання їх в обміні речовин. У процесі фотосинтезу рослинні організми синтезують органічні речовини, частину яких використовують для побудови самого організму, а частину витрачають як джерело енергії. До складу органічних речовин входять різні хімічні елементи, які надходять до рослин із ґрунту.
Ґрунт є активним середовищем живлення рослин і складається з органічних, мінеральних і органомінеральних компонентів, із яких під дією абіотичних і біологічних процесів продукуються доступні для рослин поживні речовини. Поживні елементи — основна складова частина, що характеризує родючість ґрунту. Родючість зумовлюється здатністю ґрунту забезпечувати рослини водою, повітрям (киснем), теплом (для коренів) і сприятливими фізичними та фізи-ко-механічними умовами для росту і розвитку рослин. Родючість ґрунту — основна якісна ознака, яка відрізняє ґрунт від гірської породи та пасивного субстрату.
Під час кореневого живлення рослини в основному поглинає з ґрунту хімічні елементи (макро-, мікро- та ультрамікроелементи).
Макроелементи — це хімічні елементи, вміст яких у рослині від десятків відсотків до сотих часток відсотка (С, О, Н, І, 8і, К, Mg, Р, 8). Залізо міститься на межі між макро- та мікроелементами.
Мікроелементи — це елементи, вміст яких у рослині становить від сотих до стотисячних часток відсотка. До цієї групи належать марганець, бор, хлор, мідь, цинк, нікель, молібден, кобальт тощо.
Ультрамікроелементи — це хімічні елементи, вміст яких у рослині становить від мільйонних часток відсотка. До цієї групи належать цезій, кадмій, срібло, радій тощо.
Елементи мінерального живлення забезпечують у рослинній клітині як структурну, так і каталітичну функції.
У значній кількості йони елементів мінерального живлення входять до складу органічних сполук рослинної клітини.
Органогени — вуглець, водень, кисень — універсальні компоненти майже всіх органічних сполук; азот і сірка — компоненти білків, нуклеїнових кислот, порфіринів.
Метали — залізо, марганець, цинк, молібден і кобальт входять до складу ферментів або їхніх кофакторів. Молібден і кобальт входять до складу ферментів азотфіксації. Крім того, молібден бере участь у відновленні нітратів, а марганець — у фотолізі води. Залізо необхідне для синтезу хлорофілу. Йони Са2+, СІ- беруть участь у процесах фотосинтетичного виділення кисню.
Такі елементи, як залізо, марганець, мідь, молібден, кобальт, входять до складу активних груп або компонентів простетичних груп ферментів, особливо оксидоредуктаз, які забезпечують процеси фотосинтезу, дихання (флавопротеїни, фередоксини, цитохроми, пластоціанін, фенолоксидази та ін.). Як кофактори перелічені елементи забезпечують утворення хелатів, поєднання ферментів або коферментів із субстратами (наприклад, марганець, магній, цинк).
Фосфор і бор трапляються у вигляді залишків фосфатної та борної кислот (АТФ, фосфати цукрів, нуклеїнові кислоти).
Калій, магній та кальцій впливають насамперед на гідратацію колоїдів протоплазми. Калій впливає на активність майже 60 ферментів. Солі кальцію та магнію входять до складу серединних пластинок (пектатів, фітину). Магній часто функціонує як стабілізатор структури в рибосомах, кальцій виконує цю саму функцію в хромосомах і мембранах.
Отже, основними функціями йонів у метаболізмі є структурна й каталітична.
Мінеральні елементи поглинаються рослинним організмом одночасно з водою за допомогою кореневої системи. В невеликих кількостях мінеральні елементи можуть надходити і через листки, тому позакореневе підживлення рослин, особливо мікроелементами, — поширений агрозахід.
Більшість рослин поглинають воду пасивно — силою, що утворюється завдяки різниці між осмотичним і тургорним тиском. Рослини, що адаптувалися до існування на засолених субстратах, використовують активне транспортування води проти градієнта концентрації солей, витрачаючи на це значну частину продуктів асиміляції. Через це вони завжди низькорослі. Мінеральні речовини рослини поглинають активним всмоктуванням. Проте рослини здатні не тільки поглинати мінеральні речовини із ґрунтового розчину, а й розчиняти нерозчинні у воді сполуки. Цьому сприяють виділювані рослиною органічні кислоти — яблучна, лимонна та ін.
Через різницю в концентрації солей ґрунтового розчину і цитоплазми клітин епіблеми виникає осмос — переміщення розчинника з ґрунту у волосконосні клітини. Відомо, що концентрація речовин у клітинах кореня зростає від периферії до центра (градієнт концентрації). Внаслідок цього вода й розчинені в ній речовини переміщуються до судин центрального циліндра кореня, і виникає кореневий тиск, під дією якого розчин рухається до стебла. Крім кореневого тиску (нижній водяний насос) рух розчину по судинах підтримує також процес транспірації в листках (верхній водяний насос). Кореневим тиском вода ніби «закачується» до ксилеми, а транспірація забезпечує її транспортування на потрібну висоту.
Роль мінеральних речовин у процесах життєдіяльності рослин в той чи інший період вегетації визначають методом водних культур. Водна культура — це рослини, вирощені без ґрунту в посудинах з водними розчинами мінеральних солей при надходженні в розчин повітря (аерація розчину). При цьому використовують різні варіанти поживних середовищ, змінюючи в них вміст тих чи інших компонентів і порівнюючи характер вегетації рослин на цих середовищах з вегетацією культур, для вирощування яких використовують«стандартний» набір речовин.
Рух неорганічних та органічних речовин по кореню. Рух води і розчинених у ній речовин у рослині відбувається переважно двома шляхами: дифузією та потоком. Дифузія води і речовин відбувається за градієнтом концентрації, а рух потоком — за градієнтом гідростатичного тиску. По судинах вода рухається, як по трубах, за загальними законами гідродинаміки, а в паренхімних клітинах — осмотичним шляхом, причому пересування води в живих клітинах значно утруднене.
У корені рух води та розчинених у ній речовин починається із всмоктування її кореневими волосками. Із волосків до ксилеми центрального циліндра вода надходить через цитоплазму живих клітин кори кореня, а також по клітинних стінках. Таким шляхом вода пересувається повільно й на невелику відстань. Нарешті, вода й розчинені в ній речовини надходять до ксилеми (ксилемний сік), а далі ксилемний сік по судинах ксилеми рухається завдяки кореневому тискові. По ксилемі кореня можуть переміщуватися й органічні речовини, наприклад запасні речовини кореня навесні.
Рослина поглинає воду за допомогою кореня, який нагнітає воду до пагонів та листків, що її випаровують. Нижній кінцевий двигун — це кореневий тиск, в основі якого лежить явище осмосу. Осмотична концентрація пасоки в багато разів вища, ніж зовнішнього середовища, що й спрямовує рух води в клітини кореня. Основним осмотичним компонентом пасоки є йони К+, які зумовлюють 3/4 її загального осмотичного потенціалу. Вода і розчинний у ній К+ постійно рухаються від кореня до судин стебла та листків. Для того, щоб надходження води було неперервним, калій має постійно надходити в клітини кореня та активно транспортуватися в судини.
Верхній кінцевий двигун транспортування води — це транспірація крізь продиховий апарат. Вирішальне значення в регуляції продихового апарату належить, знову ж таки, йонам К+. Вночі, коли продихи закриті, калій рівномірно розподіляється між усіма клітинами епідермісу. Перші промені сонця слугують сигналом для відкривання продихів. Розпочинають функціонувати мембранні йонні насоси, що перекачують калій в замикаючі клітини продихів із сусідніх клітин. Вже протягом кількох хвилин концентрація К+ в замикаючих клітинах, а отже й осмотичний потенціал їхнього клітинного соку, зростає в 4-5 разів. У результаті цього замикаючі клітини продихів вбирають воду, набухають, і продихи відкриваються.
У темряві йонні насоси не функціонують, і надлишок калію переноситься за градієнтом концентрації із замикаючих клітин продиху до найближчих сусідніх. Вбирання води клітинами продихів послаблюється, тургор в них спадає, продихи закриваються.
Таким чином, транспортування йонів (у даному разі К+) є тим привідним механізмом, який контролює рушійні сили водного обміну рослин — кореневий тиск і транспірацію.
Добрива. З кожним урожаєм із ґрунту виноситься певна частина мінеральних речовин, і він поступово виснажується. Запас необхідних елементів поповнюється мінеральними (амонію сульфат, калію хлорид, суперфосфат, фосфоритне борошно; калієва, кальцієва і натрієва селітри та ін.) та органічними (перегній, торф, торфокомпос-ти, зелені добрива, пташиний послід) добривами.
Кількість добрив, які потрібно внести в ґрунт, визначають за допомогою хімічного аналізу ґрунту. Як надлишок окремих елементів у ґрунті, так і їх нестача можуть негативно впливати на врожайність культур. Строки внесення добрив визначають з урахуванням їх здатності розчинятись у воді. Важкорозчинні (фосфатні) й нерозчинні (органічні) добрива вносять восени, щоб до весни вони під дією ґрунтових організмів розклалися до розчинних у воді мінеральних сполук і з талими водами надійшли в ґрунт. Добрива можна вносити в окремі фази розвитку рослин як підживлення.
3.3. Будова і функції пагона
Стебло з розміщеними на ньому листками називають пагоном. У процесі еволюції пагін сформувався як комплексний орган, пристосований для ефективного фотосинтетичного процесу, транспірації, утворення репродуктивних органів, виконання опорної і транспортної функцій. При цьому відбулися відособлення осьової стеблової структури з відгалуженнями і формування плоских бічних виростів з обмеженим ростом — листків, які забезпечили можливість ефективного контакту рослин з повітряним середовищем і поглинання світлової енергії. Характерною структурною особливістю типового пагона вищих рослин є наявність на ньому бруньок — зачаткових пагонів, здатних тривалий час зберігати життєздатність меристем і забезпечувати їх захист від несприятливих факторів.
Термін «стебло» використовують лише в розумінні його як осьової частини пагона. Даючи анатомічну характеристику, наприклад, структур крони дорослого дерева, зважають на те, що вони є результатом перетворення системи пагонів, які виникли раніше, розгалужувались і з часом втратили недовговічні листки, а з вегетативних бруньок сформувалися нові пагони. Саме тому брунька є зачатком не стебла, а цілого пагона. З огляду на це можна дати таке визначення пагона: це нерозгалужене стебло з листками і бруньками на ньому. Річним називають пагін, що розвинувся з вегетативної бруньки за один вегетаційний період.
Класифікація та типи бруньок. Брунька — це зачаток нового пагона. Це укорочений пагін з тісно притисненими один до одного зачатками листя.
Бруньки поділяються на вегетативні та генеративні. Розрізняють бруньки за місцем їх розміщення — верхівкові й пазушні — та функціональним призначенням — вегетативні, квіткові, сплячі, додаткові. Верхівкові бруньки містяться на верхівках стебла та його бічних відгалужень. Зовні бруньки захищені лусками (видозмінені листки). Зовнішні луски зимуючих бруньок щільні, шкірясті, можуть бути вкриті кутикулою або клейкими смолянистими речовинами. Під лусками є вкорочений зародковий пагін із тісно зближеними зачатковими листками, які вкривають твірну тканину конуса наростання. У пазухах зачатків нижніх листків розміщені зачатки пазушних бруньок, із яких розвиваються бічні пагони й листки.
Пазушні бруньки дерев і кущів бувають вегетативними (ростовими) із зачатками листків і стебла, та квітковими із зачатками квіток або суцвіть. Вони відрізняються за формою: ростові — видовжені, із загостреною верхівкою, а квіткові — округлі й більші за розміром.
Деякі пазушні бруньки можуть залишатися в стані спокою неви-значено довго. Це сплячі бруньки. Вони розвиваються при пошкодженні верхівкових бруньок, зламуванні стебла над ними.
Велике значення має формування додаткових бруньок, які закладаються на стеблах, листках, коренях і слугують для вегетативного розмноження квіткових рослин.
Зовнішня будова і функції стебла. Стебло як осьова частина пагона забезпечує зв'язок усіх частин рослини, збільшує її поверхню за рахунок галуження, утворює і несе на собі бруньки й листки, здійснює транспортування води, мінеральних і органічних речовин,
слугує для вегетативного розмноження і фотосинтезу, запасає поживні речовини. Стебла без листків, хоча б зародкових або рудиментарних, як і листків без стебла, хоча б укороченого, не буває.
В оптимальних умовах стебло росте постійно, даючи бічні відгалуження і збільшуючи листкову поверхню. На пагоні є вузли й міжвузля. Вузол — це місце прикріплення листків до стебла. Міжвузля — відстань між сусідніми вузлами. Міжвузля можуть бути довгими, тоді пагін називається видовженим (ростовим). Вкорочений пагін має короткі міжвузля. На стеблах деяких рослин з дуже короткими міжвузлями зближені листки утворюють прикореневу розетку (кульбаба, морква). На плодових деревах і кущах утворюються пагони обох типів: вкорочені з малим приростом, на яких формуються квіти, а потім плоди, і видовжені, зазвичай безплідні.
Стебло характеризується необмеженим ростом. Воно росте все життя за рахунок верхівкової, рідше вставної меристеми.
На стеблі у покритонасінних утворюються квіти, а потім плоди з насінням.
У більшості випадків стебло має циліндричну форму і радіальне розташування тканин.
Стебло розгалужується екзогенно.
Стебло виконує декілька функцій.
Провідна. Стебло зв'язує дві площі живлення — корінь і листки. По стеблу проходить течія розчинених у воді мінеральних речовин із кореня до листків і течія асимілятів із листків у корінь.
Запасаюча. Накопичує в тканинах поживні речовини.
Опорна. Виносить листки до світла й орієнтує їх у просторі у певному положенні.
Стебло може бути органом вегетативного розмноження.
Молоді зелені стебла виконують функцію фотосинтезу.
Розміри стебла від 1,5 мм у ряски до понад 100 м у евкаліптів і мамонтового дерева.
За формою поперечного розрізу стебло буває округле (злаки, жовтецеві), трикутне (осоки), чотирикутне (губоцвіті), плоске (півники болотні), діжкоподібне (баобаб). Є чотири типи галуження стебла: дихотомічне, моноподіальне, симподіальне і несправжньо дихотомічне (рис. 3.8).
При дихотомічному типі галуження конус наростання ділиться на дві рівні частини (роздвоюється).
У ялини можна дослідити монопо-діальне галуження з необмеженим верхівковим ростом пагонів. При такому галуженні верхівкова брунька розвивається навесні в новий річний пагін, який продовжує наростання пагона попереднього року. З пазушних бруньок утворюються бічні пагони, але вони в процесі росту ніколи не переганяють головний.
При симподіальному галуженні наростання пагонів у довжину кожен наступний рік відбувається за рахунок розкриття бічної бруньки, розташованої під верхівковою. Головна вісь у результаті має звивисту форму. Цей тип галуження характерний для черемухи, вишні, яблуні, картоплі. Із водних рослин симподіальне галуження стебла у рдесників (родина Potamogetonaceae).
На гілочці каштана кінського можна побачити відмерлу верхівкову бруньку і два бічних пагони, що виникли з супротивних бічних бруньок. У результаті такого галуження дворічна гілка набуває вил-частої форми. Вказаний тип галуження відрізняється від симподіа-льного тим, що в ріст рушають одночасно дві бруньки. Таке галуження називають несправжньо дихотомічним.
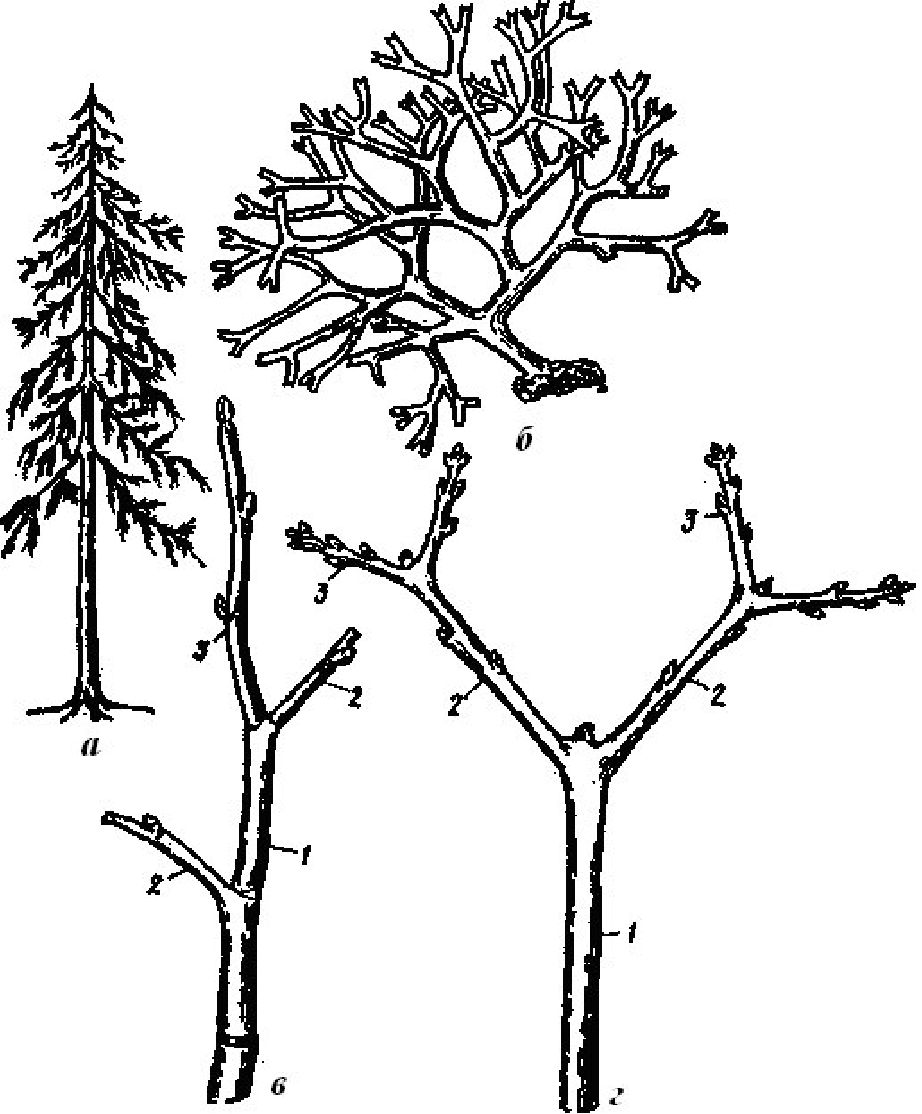
Рис. 3.8. Типи галуження: а - моноподіальне у ялини; б - дихотомічне водорості диктіоти; в - симподіальне черемухи; г - несправжньо дихотомічне клену татарського (1-2-3 і т.д. - осі першого і наступних порядків)
Листорозміщення буває спіральне (чергове - слива, льон), супротивне (гвоздика) і кільчасте (елодея канадська, м’ята кільчаста).
Анатомічна будова стебла. На верхівці стебла (головного і бічних) знаходиться верхівкова брунька, в яку замкнений конус наростання, або точка росту, що складається з меристематичної тканини. У бруньці він захищений від несприятливих зовнішніх впливів листковими зачатками і бруньковими лусками. Конус наростання дуже малий: його довжина в середньому 1—1,5 мм. Верхівкова меристема конуса наростання формує первинну будову стебла і визначає його ріст у довжину, а також утворює листки, бічні пагони і генеративні органи.
Унаслідок диференціювання верхівкових меристем виникає первинна будова стебла. В однодольних рослин вона зберігається впродовж усього життя, у дводольних, завдяки діяльності камбію, слідом за первинною формується вторинна будова стебла.
Первинна і вторинна будова стебла
При первинній будові в осьових органах рослин — стеблі й корені звичайно розрізняють три частини: 1) покривну тканину (епідерму), 2) первинну кору, 3) центральний циліндр, або стелу.
1) Епідерма оточує стебло зовні.
2) Первинна кора розташовується під епідермою. Вона утворена багатошаровою паренхімною тканиною, клітини якої часто містять хлоропласти і здатні до фотосинтезу. Крім паренхіми в первинній корі можуть бути присутніми й інші тканини. Тканини первинної кори можуть мати міжклітинники, а у водних рослин вони такі великі, що первинна кора перетворюється на аеренхіму.
3) Центральний циліндр, розташований усередину від первинної кори, звичайно складається з трьох частин — перициклу, провідних тканин і серцевини.
Перицикл зовні оточує центральний циліндр. Він представлений одним або декількома шарами паренхімних клітин. У перициклі можуть закладатися додаткові бруньки, додаткові корені, утворюватися вторинні меристеми (корковий камбій, додаткові камбіальні кільця). У деяких рослин перицикл у стеблі відсутній.
Провідні тканини розташовуються під перициклом у вигляді судинно-волокнистих пучків або суцільного кільця.
У центрі стебла знаходиться серцевина. У клітинах серцевини можуть відкладатися в запас поживні речовини (крохмаль, олії). У
деяких рослин серцевина довго залишається живою, в інших — її клітини швидко відмирають і замість запасаючої функції починають виконувати опорну, а стінки клітин дерев'яніють.
Вторинна будова стебел дводольних. У стеблах дводольних при первинній будові присутній камбій, у результаті роботи якого утворюються вторинні стебла.
Кожна камбіальна клітина ділиться тангентально. Одна з двох дочірніх клітин залишається ініціальною, здатною до необмеженої кількості поділів, а інша після декількох поділів диференціюється в елемент вторинного лубу, якщо це відбувається з клітиною, розташованою назовні від шару камбію, або вторинної деревини, якщо перетворенню піддається клітина, що знаходиться до середини від камбіального кільця. При цьому в бік вторинної деревини звичайно відчленовується більша кількість (у 3—5 разів) похідних камбію, тому шар вторинної деревини, особливо в деревних рослин, набагато ширший, ніж шар вторинного лубу.
Камбій разом із пов'язаними з ним по обидва боки найближчими похідними клітинами, подібними до нього за зовнішніми ознаками, називають камбіальною зоною. Власне ініціальний камбій у цій зоні представлений одним шаром клітин.
У зонах із помірним кліматом камбій у деревних рослин працює періодично — за сезонами. Активний поділ камбіальних клітин починається навесні, влітку уповільнюється, а восени зовсім припиняється. Отже відкладення нових шарів вторинного лубу і вторинної деревини також відбувається по сезонах.
Анатомічна будова стебел трав'янистих рослин після вторинних змін має свої особливості. Це, по-перше, наявність великої кількості паренхіми (первинна кора, серцевина, серцевинні промені); по-друге, великий діаметр центрального циліндра, який значно перевищує діаметр первинної кори; по-третє, розміщення механічної тканини (частіше коленхіми) на периферії стебла.
Вторинні зміни в стеблах однодольних
Як правило, однодольні зберігають первинну будову стебла на все життя. Однак існують деревні однодольні, стебла яких ростуть у товщину (драцена, юка, алое). У цих рослин у стеблі є постійно діюча меристематична зона, що розташовується між первинною корою і центральним циліндром і відповідає кільцю перициклу.
Будова стебла деревних рослин. У стеблах деревних рослин камбій працює впродовж усього життя, відчленовуючи все нові й нові шари елементів вторинної будови. Для деревних стебел характерна присутність великої кількості мертвих клітин зі здерев'янілими стінками.
Зовні деревного стебла знаходиться 1) покривна тканина, під нею — 2) первинна кора, далі розташовується 3) вторинний луб (вторинна кора з елементами флоеми: провідні елементи представлені ситоподібними трубками і клітинами-супутницями), за ним йде 4) камбіальна зона. Усередину від камбіальної зони розташовується могутня 5) вторинна деревина (провідна тканина -ксилема - вторинної деревини представлена трахеями (судинами) і трахеїдами), а в центрі стебла — 6) серцевина. Через вторинну деревину і вторинний луб тягнуться серцевинні (лубодеревні) промені.
Унаслідок періодичної, сезонної роботи камбію щороку відкладається новий шар лубу на старий, торішній. Однак межа між річними шарами, як правило, не помітна.
На відміну від вторинного лубу, сезонні зміни в деревині добре видно. Робота камбію пов'язана з листками. Він починає активно ділитися навесні, коли розпускаються листки. У цей час камбій утворює в основному широкопросвітні судини з не дуже потовщеними стінками, необхідні рослині для забезпечення листків, що розвиваються, водою і мінеральними речовинами. Коли листки закінчують свій ріст, ділення камбіальних клітин уповільнюється, і влітку камбій продукує в основному механічну тканину і в меншій кількості — товстостінні вузькопросвітні судини. Восени діяльність камбію припиняється. Наступною весною на осінню деревину накладається весняна із широкопросвітними трахеями. У результаті добре видно щорічні зони приросту у вигляді концентричних кілець, які називають річними кільцями. Вони характерні для дерев помірного клімату, де існує сезонність і камбій працює періодично. У рослин вологих тропіків, де пори року майже не розрізняються за температурою і кількістю опадів, річні кільця не утворюються.
Серцевина — це залишок первинної тканини в центрі стебла. З віком її клітини склерифікуються і запасаюча функція змінюється опорною.
Будова видозмінених стебел. У стебла такі зміни спостерігаються у зв'язку з розвитком запасаючої функції. У результаті утворюються такі видозміни стебла, як кореневища і бульби. Вони ви
конують також функцію вегетативного розмноження і забезпечують перенесення несприятливих умов (зима), залишаючись у ґрунті після відмирання надземної частини і даючи нові пагони при настанні сприятливих умов.
Кореневище зовні схоже на корінь (звідси його назва), але як стебло воно відрізняється від кореня тим, що несе листки, хоча і зародкові у вигляді лусочок, у пазухах яких знаходяться бруньки, що дають початок надземним і підземним пагонам. Кореневища мають багато водних і прибережно-водних рослин: лепеха, рогіз, очерет звичайний, хвощ водяний та ін.
Ще більшою спеціалізацією характеризуються бульби як підземні, так і надземні. Вони утворюються на осі головного стебла (капуста кольрабі) або на його бічних відгалуженнях — столонах.
Особливості будови стебла водних рослин. На будові стебел водних рослин відбиваються слабка освітленість, збіднення киснем і вуглекислим газом, забезпеченість водою, досить велика щільність води, що підтримує рослини. У зв'язку з цим стебла рослин, які занурені у воду, мають цілий ряд відмінностей від стебел сухопутних рослин. Епідерма в них слабко диференційована, продихи не розвиваються, тому що розчинені у воді кисень і вуглекислий газ стебла поглинають усією поверхнею.
Часто замість замикаючих клітин продиху видно їх ініціальну клітину. Клітини епідерми містять хлоропласти і здійснюють фотосинтез, використовуючи промені світла, що проникають крізь шар води. Первинна кора займає велику частину діаметра стебла. Паренхіма кори пухка, з великими міжклітинниками, нерідко перетворюється в аеренхіму. Часто повітроносні порожнини розділяються перегородками з дрібних клітин із хлоропластами, а довгі міжклітинники — діафрагмами, у яких між клітинами є невеликі повітряні порожнини. Вони пропускають повітря, але перешкоджають проходженню води, що у випадку ушкодження тканин стебла не дозволяє воді заповнити повітряні порожнини.
Діаметр центрального циліндра у водних рослин невеликий. Провідні пучки сильно зближені й у більшості водних рослин перетворилися на один пучок, що складається з тяжа ксилеми і тяжа флоеми. Причому ксилема звичайно розвинута слабко або зовсім відсутня, і на її місці утворюється повітряна порожнина. Це пов'язано з тим, що стебло поглинає воду всією поверхнею і не треба проводити її по стеблу. Механічної тканини в таких стеблах мало або вона зовсім відсутня. Рослину підтримує вода, тим більше, що велика кількість повітроносних порожнин надає їй плавучості.
Камбій у стеблах, занурених у воду, працює дуже слабко, і вторинна будова, як правило, не формується.
Будова і функції листка. Листок — бічний орган рослин. Як відомо, листки зі стеблом утворюють єдину систему — пагін.
Основні функції листка — фотосинтез і транспірація. Хлорофілоносна тканина забезпечує фотосинтез, а елементи системи провітрювання і покривна тканина — випаровування води і його регуляцію. Провідна система бере участь в обох процесах. У деяких рослин листки виконують додаткові функції. Наприклад, запасна функція характерна для листків сукулентів, у клітинах яких накопичується вода (алое, агава). Листки деяких рослин можуть бути органами вегетативного розмноження (бегонія, узумбарська фіалка).
Листок складається з листкової пластинки, черешка, а в ряду рослин також і прилистків. Найважливіша частина — листкова пластинка, саме в ній здійснюються фотосинтез і транспірація. Листкова пластинка, як правило, плоска і має дорзовентральну будову, тобто верхній і нижній її боки мають певні структурні та функціональні відмінності. Будова черешка подібна до стебла, а прилистків — до листкової пластинки.
Листок, на відміну від осьових органів, характеризується обмеженим ростом. Камбій у листку не утворюється, і тому він складається тільки з первинних тканин.
За будовою розрізняють прості і складні листки. Прості за формою листкової пластинки - округлі, овальні, стрілоподібні, ланцетні і ін. Також розрізняють листки за формою вершка і основи листкової пластинки (тупі, гострі, серцеподібні і ін.). За формою краю (цілокраї, зубчасті і ін).
Складними називаються листки, які на спільному черешку мають прості листочки, кожен з яких опадає самостійно (трійчастий -конюшина, горобина - непарноперистоскладний).
Існують різні види жилкування листків: просте (одна жилка) - у мохів, плаунів, хвощів і хвойних; дихотомічне - у гінкго дволопатевого; сітчасте - у дводольних; дугове і паралельне - у однодольних.
Розвиток та анатомічна будова листка. Листки закладаються в конусі наростання стебла екзогенно у вигляді листкових горбків. У їх утворенні беруть участь мантія і корпус. Недиференційовані листкові зачатки називають примордіями.
У багатьох деревних рослин листки, що розпускаються навесні, проходять два етапи розвитку: внутрішньобруньковий (закритий) і позабруньковий (відкритий). У процесі внутрішньобрунькового розвитку формуються в основному всі тканини листкової пластинки. Позабруньковий етап починається навесні після опадання брунькових лусок. Відбуваються поділ і ріст клітин, причому не тільки поверхневого шару, але і м'якуша листка. Пластинка збільшується в розмірах — у площині й у товщину. З основи листка інтеркалярно виростає черешок. Ріст листка однодольних більш тривалий, ніж у дводольних, особливо це характерно для листків злаків. їх вставні меристеми довго зберігають активність, що забезпечує також відростання листків після ушкодження або поїдання їх тваринами. Однак і в однодольних ріст листків обмежений.
Сьогодні відома тільки одна рослина, листки якої ростуть протягом усього її життя. Це голонасінна рослина вельвічія дивовижна, що росте в пустелі Наміб у Південно-Західній Африці. Вона має всього два листки, у основі яких постійно функціонує вставна мери-стематична зона. Довгі листки стеляться по землі. їх верхівки постійно старіють і відмирають, а фотосинтезуюча частина досягає кількох метрів у довжину.
Будова пластинки листка. Листкові пластинки зовні дуже різноманітні за розмірами, формою, навіть за забарвленням. Внутрішня структура їх більш однорідна, тому що забезпечує виконання їх основних функцій. У пластинці листка розрізняють чотири частини: покривну тканину, м'якуш, або мезофіл, провідну систему і систему механічних тканин (рис. 3.9).
Листок зверху і знизу вкритий епідермою. Звичайно епідерма одношарова, зрідка зустрічається дво-, тришарова (фікус, олеандр).
Епідерма захищає листок від механічних пошкоджень, проникнення патогенних мікроорганізмів, висихання. Разом з тим вона забезпечує фотосинтез, тому що, з одного боку, прозора і вільно пропускає сонячні промені до хлорофілоносних тканин, з іншого — через її продихи відбуваються газообмін, а також транспірація. Остання допомагає транспорту води і мінеральних речовин у листок і сприяє його охолодженню.
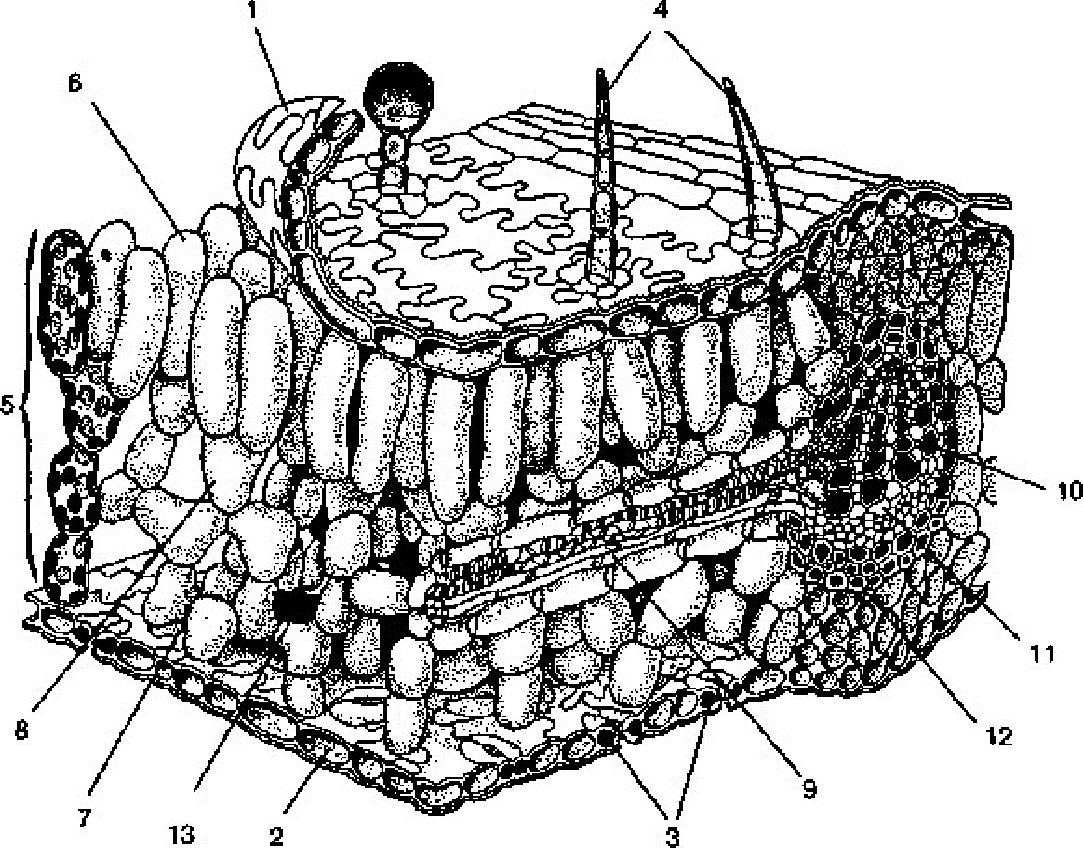
13 2 З
Рис. 3.9. Об'ємне зображення пластинки листка:
1 — верхня епідерма; 2 — нижня епідерма; 3 — продихи; 4 — волоски; 5 — мезофіл; 6 — стовпчаста тканина; 7 — губчаста тканина; 8 — збірні клітини; 9 — жилка; 10 — ксилема; 11 — флоема; 12 — склеренхіма; 13 — міжклітинники
М'якуш листка, або мезофіл, розташовується в листковій пластинці між верхнім і нижнім шарами епідерми. Мезофіл представлений хлорофілоносною тонкостінною паренхімою. Розрізняють два типи мезофілу: однорідний, або недиференційований, і диференційований. Недиференційований зустрічається в основному в однодольних і голонасінних рослин. Він складається з приблизно однакових за формою паренхімних клітин. До недиференційованого також належить складчастий мезофіл більшості хвойних.
Диференційований мезофіл представлений двома типами хлорофілоносної тканини — стовпчастою, або палісадною, і губчастою, або пухкою. Стовпчаста (палісадна) тканина звичайно розташовується під верхньою епідермою у вигляді одного шару дещо видовжених, щільно зімкнених клітин. Це основна фотосинтезуюча тканина листка. У деяких рослин є 2—3 шари палісадної тканини під верхньою епідермою (брусниця, лимон) або по одному шару під верхньою і нижньою епідермами (евкаліпт).
Між стовпчастою тканиною і нижньою епідермою розташована губчаста паренхіма. Вона багатошарова, її клітини мають округлу або лопатеву форму. Ця тканина містить багато міжклітинників.
Крім фотосинтетичної, вона виконує функцію газообміну і транспірації. Хлоропластів у клітинах губчастої тканини менше, ніж у стовпчастої, у середньому в 2—5 разів. У зв'язку з цим забарвлення нижнього боку листка світліше, ніж верхньої.
Провідна система листка представлена жилками.
У листках дводольних є одна головна жилка 1-го порядку. Жилки 1-го порядку розгалужуються, утворюючи жилки 2-го порядку, які, у свою чергу, розгалужуючись, дають жилки 3-го порядку, і так далі, аж до 7-го і 8-го порядків у деяких рослин. Більш великі жилки утворюють ребра — виступи в основному на нижньому боці листка. Дрібні жилки видно, якщо листок розглянути на світлі, тоді як дрібніші можна побачити тільки під мікроскопом. Дрібні жилки з'єднуються перемичками-анастомозами.
В однодольних є кілька головних жилок. Вони можуть мати приблизно однакову товщину, або товсті жилки можуть чергуватися з тонкішими. Жилки звичайно проходять уздовж пластинки листка паралельно (злаки) або дугами (конвалія), з'єднуючись у її верхівці. Головні жилки зв'язуються перемичками, що розташовуються поодинці або групами.
Найбільші жилки містять кілька провідних пучків, дрібніші — по одному.
Провідні пучки листкової пластинки через черешок зв'язуються з пучками стебла і далі — кореня й утворюють єдину провідну систему рослини.
Тканини провідних пучків у листку безпосередньо не контактують з клітинами мезофілу. Вони оточені одним шаром паренхімних витягнутих уздовж жилки клітин, які називають обкладкою. У більшості рослин клітини обкладки не містять хлоропластів і служать для проведення асимілятів у флоему.
У рослин зі зниженою транспірацією кінцеві жилки містять одну флоему. Узагалі, в листках рослин вологих місць, особливо у водних, ксилема розвинена слабше, ніж флоема.
Вплив факторів довкілля на будову і функціонування листка
Світлові й тіньові листки. У кронах дерев і кущів формуються світлові й тіньові листки. Світлові звичайно утворюються на периферії крони, де краще освітлення, а тіньові — усередині крони, куди потрапляє менше світла. Світловий і тіньовий тип будови пластинки листка розвивається також у трав'янистих рослин, що ростуть на відкритих місцях і в тіні.
Пластинка світлових листків характеризується більшою товщиною і жорсткістю. Стінки клітин епідерми мають дуже потовщену зовнішню оболонку. У них могутніші кутикула і восковий наліт, густіша опушеність. Вони мають більш тонкі та густіше розташовані жилки, дрібніші продихи, але їх більше на одиницю площі поверхні. У цих листків у порівнянні з тіньовими краще розвинута стовпчаста тканина, але вони бідніші на хлорофіл. У світлових листків переважає механічна тканина, а в тіньових більше міжклітинники.
Особливості будови світлових і тіньових листків обумовлені не лише різницею в освітленні, але і неоднаковими умовами водопостачання: листки, розташовані на поверхні крони, забезпечуються водою гірше, ніж ті, що ростуть внизу або всередині крони.
Умови освітлення визначають ступінь диференціювання мезофілу, розвиток стовпчастої тканини, вміст хлорофілу. Густота і товщина жилок, кутикула й опушеність епідерми, кількість продихів і інші особливості визначаються умовами водозабезпечення рослин.
Формування світлових і тіньових листків залежить також від фізіологічного стану рослини. Молоді дерева спочатку утворюють листки тіньового типу, і лише в зрілому віці розвиваються світлові.
Ярусність у будові листків. У рослин спостерігаються деякі розходження в будові пластинки листка залежно від ярусності, тобто положення їх на стеблі. Виявилося, що за будовою листки верхніх ярусів дуже схожі на світлові, а нижніх — на тіньові. Від нижнього до верхнього ярусу ознаки тіньових листків зменшуються, а світлових — зростають. Фактори, що впливають на особливості будови листкової пластинки, ті самі — освітлення і водопостачання. Зрозуміло, що листки верхніх ярусів знаходяться в більш сприятливих умовах освітлення, але потерпають від нестачі води.
У рослин, що потерпають від нестачі вологи, в епідермі можуть виникати такі пристосування:
- у клітинах епідерми розвивається товста зовнішня стінка, могутня кутикула і восковий наліт.
- шкірка, особливо нижня, часто має сильну опушеність.
- у деяких рослин епідерма складається з декількох шарів клітин.
- продихи частіше розташовані в заглибинах, тобто нижче рівня епідерми; над ними звичайно нависають виступи кутикули.
Крім змін в епідермі, у рослин розвиваються й інші пристосу
вання, що допомагають їм перенести водний дефіцит.
Сукуленти — м'ясисті соковиті рослини, які пристосувалися до запасання води у водоносних тканинах (кактус, алое, агава, молодило, очиток). Алое, агава, очиток водоносну тканину містять у листках, кактуси — у стеблах. У період дощів корені сукулентів активно поглинають воду і запасають її у вакуолях великих клітин водоносної тканини, а в посушливий період дуже ощадливо її витрачають.
У кактусів для зменшення випаровування води редукуються листки — вони перетворюються на колючки, а функцію фотосинтезу бере на себе стебло, що сильно розростається від водоносної тканини, площа якого невелика. Епідерма стебла кактуса, має всі перераховані вище пристосування до посушливих умов. Продихів у шкірці мало, та й вдень вони звичайно закриті. Під епідермою розташовані кілька рядів клітин хлорофілоносної паренхіми, глибше — безбарвна крупноклітинна тканина, що запасає воду.
Сукулентні рослини за малої кількості продихів, до того ж закритих удень, мало випаровують води. Це, з одного боку, запобігає її втраті, а з іншого — не забезпечує охолодження, що спричиняє сильне їх нагрівання — до 50—60 °С. Однак цитоплазма сукулентів пристосована до перенесення таких температур без коагуляції колоїдів і втрати життєдіяльності.
Пристосування до посушливих умов мають ксерофіти — рослини сухих степів і пустель. У них вузькі довгі (злаки) або дрібні листки. Ксерофіти в період посухи втрачають до 50 % води, але колоїди цитоплазми при цьому не коагулюють, тому що пристосовані до сильного зневоднення.
Деякі рослини сухих степів і пустель інакше пристосувалися до перенесення посушливих умов. Наприклад, верблюжа колючка на листках має тонку епідерму і кутикулу, але в неї довгий корінь, що досягає ґрунтових вод і забезпечує рослину водою.
Є група рослин із коротким вегетаційним періодом — ефемери і ефемероїди. За 20—30 днів вони встигають закінчити життєвий цикл — від насіння до насіння (тюльпани). У цих рослин є свої особливості в будові листків. За короткий період формується примітивніша листкова пластинка без будь-яких пристосувань до посушливих умов. Ефемери переживають посушливий період у вигляді насіння, а в ефемероїдів після відмирання надземної частини в грунті залишаються бульби або цибулини, які й переносять посуху.
Листопад — масове опадання листків у деревних, звичайно дводольних, рослин. Він є важливим пристосуванням до перенесення несприятливих умов (зима, посуха). Скидаючи листя, рослини різко скорочують випаровувальну поверхню, що важливо в зимовий період, коли корені не поглинають воду, а транспірація продовжується. Звільняючись від листя, дерева уникають поломок під вагою снігу. Листопад у зонах із різко вираженим посушливим періодом допомагає деревам пережити посуху, коли в ґрунті різко зменшується кількість води, а висока температура підсилює випаровування.
Ще одне важливе значення листопаду полягає в тому, що, скидаючи листя, рослина звільняється від ряду речовин, накопичених у великій кількості, гальмуючи її життєдіяльність і насамперед фотосинтез (кальцій, кремній та ін.). Саме це є основною причиною скидання листя у рослин теплого і вологого клімату.
Особливості будови листків водних рослин. Листки водних рослин можуть бути розташовані над водою, лежати на воді, бути цілком зануреними в неї. Прикладом рослини, що має всі три типи листків, є стрілолист. Його стрілоподібні листки стирчать з води, лежать на поверхні, а занурені у воду мають стрічкову форму. Два типи листків — на воді і під водою — мають латаття і глечики. Водне середовище певною мірою відбивається на будові листкової пластинки.
Надводні листки практично не відрізняються від листків рослин, що зростають на суші в досить вологих умовах. Листки, що лежать на воді, звичайно мають товсту, іноді шкірясту пластинку (латаття). Верхня епідерма, як правило, товстостінна, покрита добре розвиненою кутикулою. Очевидно, кутикула захищає листя від змочування водою. Продихи у верхній епідермі численні, а в нижній їх немає: вони там не потрібні, тому що листок поглинає гази з води. Кутикула на нижньому боці листка тонка або зовсім відсутня. Мезофіл у таких листків може бути диференційованим. Наприклад, у латаття стовпчаста тканина багатошарова і складається з дрібних клітин із великою кількістю хлоропластів. Губчаста тканина листків, які лежать на воді, містить великі міжклітинники. У мезофілі часто присутні великі астросклероцити.
У листків, цілком занурених у воду, насамперед змінюється форма пластинки. Вона стає тонкою, стрічковою або розсіченою, що збільшує площу контакту листка з водою, з якої він одержує кисень,
вуглекислий газ і мінеральні речовини. У покривній тканині відсутні кутикула й продихи, але в її клітинах є хлоропласти. Остання особливість пов'язана з тим, що вода поглинає частину сонячних променів, тому світло, яке дійшло до рослини, повинне використовуватися максимально вже в клітинах епідерми.
Мезофіл у занурених листків не диференційований. Він складається з губчастої тканини з великими міжклітинниками. Об'єм повітряних порожнин дуже великий: він може дорівнювати половині об'єму листка і більше. Наявність великих міжклітинників забезпечує запасання вуглекислого газу і кисню, що пов'язано з їх малою розчинністю у воді.
У підводних листків слабко розвинуті жилки, в них дуже мало ксилеми, оскільки листок поглинає воду всією поверхнею і нема потреби в її транспорті. Іноді на місці ксилеми в пучках утворюється повітряна порожнина, флоеми в них теж менше, ніж у повітряних листків. Це пов'язують із низькою інтенсивністю фотосинтезу занурених у воду листків.
В епідермі багатьох водних рослин, листки яких стикаються з водою, утворюються спеціальні клітини — гідропоти (рис. 3.10). За формою і функціями вони відрізняються від епідермальних. Клітинні стінки в них целюлозні, звивисті, клітини багаті цитоплазмою з дрібними хлоропластами, їх цитоплазма добре проникна для води і мінеральних речовин. Вважають, що гідропоти залежно від потреби рослин можуть поглинати воду або виділяти її надлишок.
Гідропоти частіше розташовуються в епідермі дифузно — по одній або групами (жабурник, латаття, глечики), а в рдесника, наприклад, уся нижня епідерма представлена гідропотами.
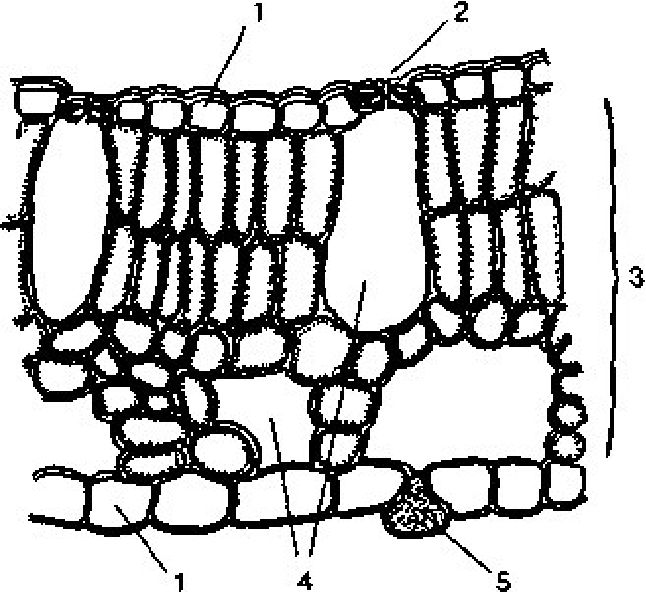
Рис. 3. 10. Поперечний зріз пластинки листка жабурника: 1 —
епідерма; 2 — продихи; 3 — мезофіл; 4 — повітряні порожнини; 5
— гідропот
Контрольні запитання та завдання
1. Що таке орган? Які органи є у рослин? Що таке метаморфози органів?
2. Які функції виконують корені?
3. Які існують види коренів? Порівняйте типи кореневих систем.
4. Опишіть зони кореня.
5. Де розташована зона всмоктування кореня і яка її будова?
6. У чому полягають особливості анатомічної будови кореня?
7. Охарактеризуйте первинну і вторинну будову кореня.
8. Які існують метаморфози коренів? У яких рослин утворюються повітряні корені?
9. Що називають симбіозом? Який симбіоз коренів рослин?
10. Що таке мікориза? Яке значення мікоризи для рослин?
11. У чому полягає кореневе живлення рослин? Як класифікують елементи мінерального живлення і які їх функції в організмі рослин?
12. Як відбувається рух неорганічних та органічних речовин по кореню?
13. Розкажіть про класифікацію та типи бруньок.
14. Охарактеризуйте будову і функції пагона.
15. Яка будова видозмінених стебел?
16. Анатомічна будова стебла.
17. Особливості будови стебла водних рослин.
18. Як відбувається розвиток листка? Опишіть анатомічну будову листка.
19. Охарактеризуйте вплив факторів довкілля на будову і функціонування листка
20. Якими є особливості листків водних рослин?