БОТАНІКА 3 ОСНОВАМИ ГІДРОБОТАНІКИ - 2010
6. ВОДОРОСТІ (ALGI)
6.1. Загальна характеристика водоростей
Поняття «водорості» (Algae) є не систематичним, а біологічним. Слово «водорості» означає рослини, які живуть у воді. Але в ботаніці цей термін має вужче значення, і не всі рослини, які можна побачити у водоймах з наукової точки зору можна називати водоростями. З іншого боку, саме водорості ми часто просто не помічаємо у водоймах, оскільки більшість водоростей не можна побачити неозброєним оком.
Придивившись до різних водойм, особливо до озер і ставків, ми у першу чергу помічаємо насінні, або квіткові, рослини, завдяки їх розміру і великій кількості. Біля берегів ростуть очерет і рогіз, які прикріпляються корінням до дна водойми, а стебло з листям і квітками піднімаються над водою. Далі від берега можна знайти рослини з плаваючим на поверхні води листям і квітами, які лише трохи піднімаються над водою, наприклад латаття біле або глечики жовті. Іноді вся поверхня ставу буває повністю затягнута плаваючими дрібними зеленими пластинками ряски. І, нарешті, дуже численні рослини цілком занурені у воду. Деякі з них прикріплені до дна, наприклад, більшість рдесників, елодея і водопериця; інші з дном не пов'язані — пухирник і кушир. Всі ці рослини, які б вони не були різні на вигляд, є насінними, або квітковими, хоча деякі з них майже ніколи не квітнуть і не утворюють насіння, а розмножуються переважно вегетативно. Для цих рослин науковий термін «водорості» непридатний, їх називають водними рослинами.
Окрім насінних водних рослин, у водоймах можна зустріти і представників вищих спорових рослин — мохів, хвощів і папоротеподібних. Більшість мохів — вологолюбні рослини, але типово водяних, таких, що ростуть зануреними у воду, серед них не так багато. Один з найвідоміших — мох фонтиналіс, який створює у воді ставів і річок темно-зелені дерновини із стебел, вкритих листям розміром до 8 мм. Біля берегів росте водяний хвощ, з характерними кільцями бічних гілочок. На мулистих мілководдях неглибоких чистих стоячих водойм і водойм з повільною течією дуже рідко можна побачити водяну папороть сальвінію, яка занесена до Червоної книги України.
Нарешті, продовжуючи дослідження водойми можна помітити і справжні водорості. До них відносяться, наприклад, великі зелені скупчення так званої твані, що плаває в літній час поблизу від поверхні води в ставках і затишних місцях річок і озер. Різноманітні зелені і синьозелені плівки та пухкі нарости на каменях, колодах і палях також утворені водоростями. У літній час вода в ставах часто буває забарвлена в зеленуватий колір, і якщо зачерпнути її склянкою, то на просвіт можна помітити в ній найдрібніші водорості у формі плаваючих точок, пластівців або кульок. Тут можна побачити і більші за розміром водорості у вигляді простих або розгалужених ниток, або і великі харові водорості, зовні схожі на хвощ, з характерними кільцями бічних пагонів.
Крім того, значна кількість мікроводоростей зустрічаються і на суші: на поверхні і в товщі грунту, на деревах, каменях і т.п.
Чим же всі ці рослини, що об'єднуються під поняттям «водорості», відрізняються від інших рослин?
У морфологічному відношенні для водоростей найістотнішою ознакою є відсутність (навіть при дуже складній зовнішній розчленованості тіла) справжніх органів - стебел, листя і коріння, які є типовими для вищих рослин. Тіло водоростей являє собою талом або слань.
Водорості розмножуються або вегетативно, або за допомогою спор, тобто відносяться до спорових рослин (на відміну від насінних, або квіткових, рослин).
Вегетативне розмноження - брунькуванням, поділом клітини надвоє, багатоклітинні нитчасті водорості розмножуються уривками ниток, харові - розпадінням колоній і «бульбочками».
Власне нестатеве розмноження - це розмноження спорами, рухомими (зооспорами) або нерухомими (апланоспорами). Спори утворюються в спеціальних органах - спорангіях або всередині вегетативних клітин.
Статеве розмноження або відтворення за допомогою статевих клітин - гамет, які зливаючись утворюють зиготу. Після періоду спокою зигота проростає, утворюючи зооспори або нову рослину. У морських водоростей зигота проростає без періду спокою.
Є чотири типи статевого процесу у водоростей - ізогамія, гетерогамія, оогамія і кон’югація.
Розвиток спор або гамет у водоростей залежить від умов оточуючого середовища. Якщо вони сприятливі - розмноження нестатевим шляхом, у несприятливих утворюються гамети. Інколи спори і
гамети утворюються в клітинах однієї особини, але частіше спори на одних особинах (спорофітах), а гамети на гаметофітах. Гаметофіт може бути одностатевим або двостатевим. У одних водоростей спорофіт і гаметофіт зовнішньо подібні і у циклі розвитку рівнозначні, у інших переважає гаметофіт, а спорофіт - недовговічна мікроскопічна рослина або навпаки недорозвинутий гаметофіт.
У більшості водоростей редукційний поділ (мейоз) проходить при проростанні зиготи і весь життєвий цикл проходить у гаплоїдній фазі, а диплоїдна представлена зиготою. У деяких водоростей редукційний поділ проходить у гаметангіях перед утворенням гамет, тоді у їх життєвому циклі переважає диплоїдна фаза, а гаплоїдна представлена лише гаметами. Таку зміну гапло- і диплофаз називають зміною ядерних фаз.
Водорості містять хлорофіл, завдяки якому вони здатні асимілювати на світлі вуглекислий газ, тобто харчуватися фототрофно. Більшість водоростей характеризуються здатністю до фотоавтотрофного способу живлення і містять хлорофіл. Проте серед них відомі і безбарвні, і навіть беспластидні організми, що живляться гетеротрофно (сапротрофи, паразити і організми з голозойним способом живлення). Вони втратили здатність до фотоавтотрофного способу живлення в результаті пристосування до особливих умов існування в багатих органічними речовинами біотопах.
Отже, водорості — це нижчі спорові рослини, тіло яких не розчленоване на органи, а являє собою талом, або слань, вони містять у своїх клітинах хлорофіл і живуть переважно у воді. За будовою тіла водорості — від мікроскопічних організмів — одноклітинних, колоніальних і багатоклітинних, до великих за розмірами рослин різної форми. Велику різноманітність мають способи розмноження і будова органів розмноження. Навіть за забарвленням водорості неоднакові, оскільки одні містять лише хлорофіл, інші — ще ряд додаткових пігментів, що забарвлюють їх в різні кольори.
Альгологія, фікологія — це розділ ботаніки, наука, що вивчає водорості. Особливе значення для розвитку сучасної альгології мало удосконалення електронно-мікроскопічної техніки і методів біохімічного аналізу, а також розробка способів виділення чистих альгологічних культур і створення в різних країнах колекцій штамів водоростей. Все це стимулювало розвиток експериментальних робіт. Розширення об'єму альгологічних досліджень привело до істотного збільшення інформації про водорості, у результаті чого відбулася подальша диференціація альгології, і на стику альгології з іншими науками виникли нові наукові напрями (палеоальгологічні, радіобіологічні та ін.).
6.2. Будова тіла водоростей
Вегетативне тіло водоростей представлене сланню, або таломом, не диференційоване на органи — корінь, стебло, листя і квіти. Водорості - це одноклітинні, багатоклітинні, неклітинні (сифональні) і сифонокладальні організми, що живуть поодинці або створюють різноманітні угруповання. Розміри їх коливаються в широких межах: найдрібніші не перевищують 1 мкм в діаметрі, а найбільші -морські бурі водорості — досягають декілька десятків метрів у висоту. Форма і забарвлення їх також різноманітні і часто химерні. Поряд з простими округлими або видовженими зустрічаються водорості зі складно розчленованим тілом, що нагадує вищі рослини.
Величезну різноманітність водоростей можна звести до декількох типів морфологічної структури, які відповідають основним рівням морфологічної диференціації тіла водоростей у процесі їх еволюції. На даний час розрізняють такі типи структури: монадний, гемімонадний, кокоїдний, сарциноїдний, нитчастий, різнонитчас-тий, паренхіматозний, псевдопаренхіматозний, сифональний, сифо-нокладальний, амебоїдний.
Монадний (джгутиковий) тип структури. Найважливішою ознакою цього типу структури є наявність джгутиків - органоїдів руху. Рухомі джгутикові форми домінують серед евгленових, дино-фітових, криптофітових, рафідофітових, золотистих, спостерігаються у жовтозелених і зелених водоростей. У діатомових і бурих монадний тип структури у вегетативному стані відсутній, проте джгутикові стадії утворюються під час репродукції. Лише у червоних водоростей монадні форми і стадії не виявлені.
Завдяки наявності джгутиків швидкість руху монадних форм на 1—3 порядки перевищує швидкість рухомих мікроорганізмів, що не мають джгутикового апарату.
Основна форма клітини — краплеподібна, радіально- або білатерально-симетрична, зі звуженим переднім джгутиковим полюсом (рис. 6.1). Проте часто монадні організми відхиляються від цієї основної форми і можуть бути асиметричними, спіралевидними і т.д.

Рис. 6. 1. Монадна структура у зелених водоростей:
1 - одиночна клітина хламідомонади (Chlаmydоmоnаs);
2 - колонія піроботриса (РугоЬоІ^), утворена зростанням клітин; 3 - колонія евдорини (ЕМогіпа), утворена слизом
На передньому кінці клітини часто розташована глотка, що зазвичай виконує екскреторну функцію. Лише у небагатьох безбарвних джгутикових глотка функціонує як клітинний рот (цитостом) і служить для прийому твердої їжі. Тверді частинки їжі, що потрапили в глотку, переходять у травні вакуолі, що від’єднуються від глотки, які спускаються до клітинного анального отвору (цитопрокту), де неперетравлені частинки виштовхуються назовні.
Своєрідним органоїдом, властивим водоростям з монадною структурою, є також скоротливі вакуолі, що виконують осморегу-ляторну функцію, слизисті тільця і жалкі структури, які виконують захисну функцію. Є стигма (вічко).
Ядро займає в клітині монадних організмів осьове, часто центральне, положення. Хлоропласти, різноманітні за формою, забарвленням і тонкою структурою, можуть бути осьовими або пристінними.
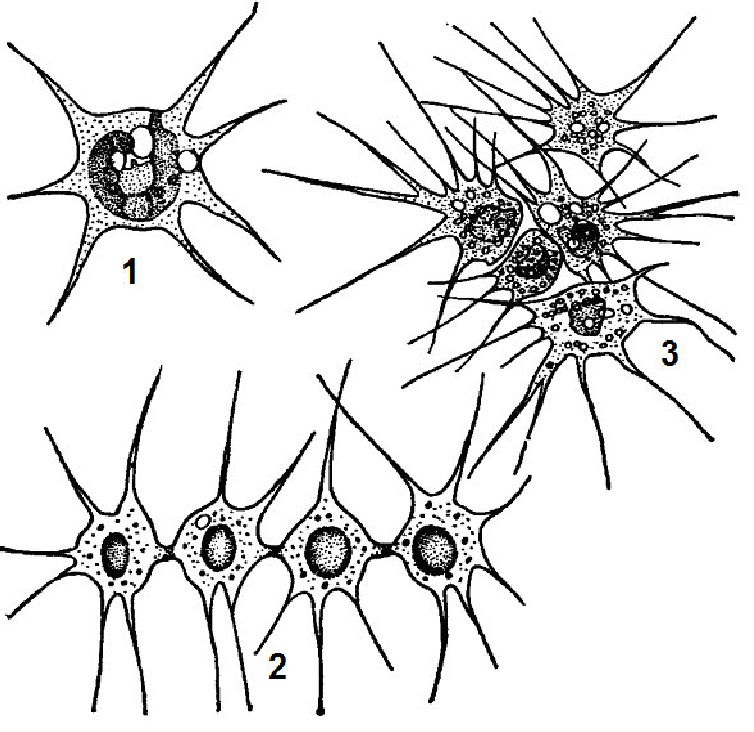
Тенденція до збільшення розмірів тіла виявляється у монадних організмів в утворенні різноманітних за формою і будовою колоній: ланцюжкових, кільцеподібних, пластинчастих, деревоподібних і ін.
Іноді клітини утримуються в колонії за допомогою тонких плазматичних ниток і слизу або без нього. Зеленим монадним організмам переважно властиві колонії типу ценобіїв з постійним для кожного виду числом клітин, не здатних до вегетативного клітинного поділу; зростання ценобіїв відбувається лише за рахунок збільшення розмірів клітин, що їх складають. На прикладі вольвоксових
(Уоіяосасеае) можна прослідкувати поступове ускладнення ценобі-їв, що досягає найвищого рівня у видів роду вольвокс (Уоіяох), між клітинами яких, диференційованих на вегетативні і генеративні, існує плазматичний зв'язок.
У несприятливих умовах монадні організми скидають або втягують свої джгутики, втрачаючи при цьому рухливість, і оточують себе густим слизом.
Монадний тип структури виявився еволюційно перспективним. На його основі розвинулися інші, складніші структури, пов'язані з втратою рухливості у вегетативному стані.
Амебоїдний (ризоподіальний) тип структури. Найхарактернішими ознаками, що визначає цей тип структури є відсутність міцних клітинних покривів і здатність до амебоїдного руху. Під амебоїдним рухом розуміють всі форми переміщення організмів за допомогою тимчасових утворень на їх поверхні — цитоплазматичних виростів — псевдоподій.
У клітинах амебоїдних водоростей є ядра, пластиди і інші органоїди, властиві еукаріотам; нерідко спостерігаються також скоротливі вакуолі, стигми і базальні тіла, здатні виробляти джгутики.
Багато амебоїдних організмів ведуть прикріплений спосіб життя. Вони можуть будувати будиночки різної форми, тонкі, ніжні, або товстостінні, грубі, безбарвні або забарвлені.
Нерідко амебоїди утворюють колонії різноманітних розмірів і форми, як з допомогою слизу, так і без нього (рис. 6.2).
Рис. 6. 2. Амебовдна структура у золотистих водоростей: 1 - одиночні клітини хризамеби ( Chrysamoeba); 2 - рядове об'єднання клітин хризидіаструм (Chrysidiastrum); 3 - групове обє-днання клітин рнзохризнс (Rhizochrysis)
Амебоїдний тип структури розповсюджений не так широко, як монадний. Він спостерігається лише в тих відділах (золотисті, жов-тозелені і динофітові), в яких різноманітно представлені монадні форми, причому між монадними і амебоїдними організмами існує поступовий перехід. Амебоїдний тип структури у водоростей є, на думку багатьох учених, виразом крайньої спеціалізації, еволюційною безвихіддю, що виникла в результаті пристосування монадних форм до особливих умов існування в біотопах, багатих органікою.
Гемімонадний (пальмелоїдний) тип структури. Характерним для цього типу структури є поєднання нерухомого рослинного способу життя з наявністю клітинних органоїдів, властивих монадним організмам: скоротливих вакуолей, стигм, джгутиків або їх похідних. Так, вегетативні клітини можуть мати джгутики, за допомогою яких вони обмежено пересуваються в межах колоніального слизу ^Іоеососсш), або джгутики зберігаються в нерухомих клітинах в дуже редукованому стані (Astеrососсus, СйІогапріеІІа).
Клітини вкриті слизом або без нього, одиночні або в колоніях, прикріплені (мають спеціальні пристосування у вигляді слизистих подушечок, стебел або підошов) або вільноживучі.
Виникнення гемімонадного типу структури було важливим етапом на шляху морфологічної еволюції водоростей від рухомих монадних до типово рослинних нерухомих форм.
Кокоїдний тип структури об'єднує одноклітинні і колоніальні водорості, нерухомі у вегетативному стані. Клітини кокоїдного типу оточені оболонкою і мають протопласт рослинного типу (з тоноп-
У нейстонних водоростей спостерігаються спеціальні пристосування, що утримують їх в поверхневій плівці води, у вигляді плавальних ковпачків (рис. 6.3).
Рис. 6. 3. Г емімонадна (пальмелоїдна) структура і пальмелоїдний стан:
1 - пальмелоїдна структура у золотистої водорості гідруруса (Нуйгигш); 2 - пальмелоїдний стан у зеленої водорості хламідомонади (Chlamydomonas) ластом, без скоротливих вакуолей, стигм, джгутиків).
Водорості кокоїдного типу мають різноманітну, часто химерну форму клітин від початкової кулястої до еліпсоїдної, веретеновидної, циліндричної, яйцевидної, грушовидної, спіралевидної, серцеподібної, блюдцеподібної, зірчасто-лопатевої і т.д. їх різноманіття збільшується завдяки різним виростам — гранулам, шпилькам, шипикам, щетинкам і ін.
Клітини кокоїдних водоростей мають одне або багато ядер. Хлоропласти різноманітні за формою і положенням у клітині, іноді вони відсутні.
Кокоїдні водорості утворюють колонії різноманітної форми, в яких клітини об'єднуються за допомогою слизу або без нього (рис. 6.4).
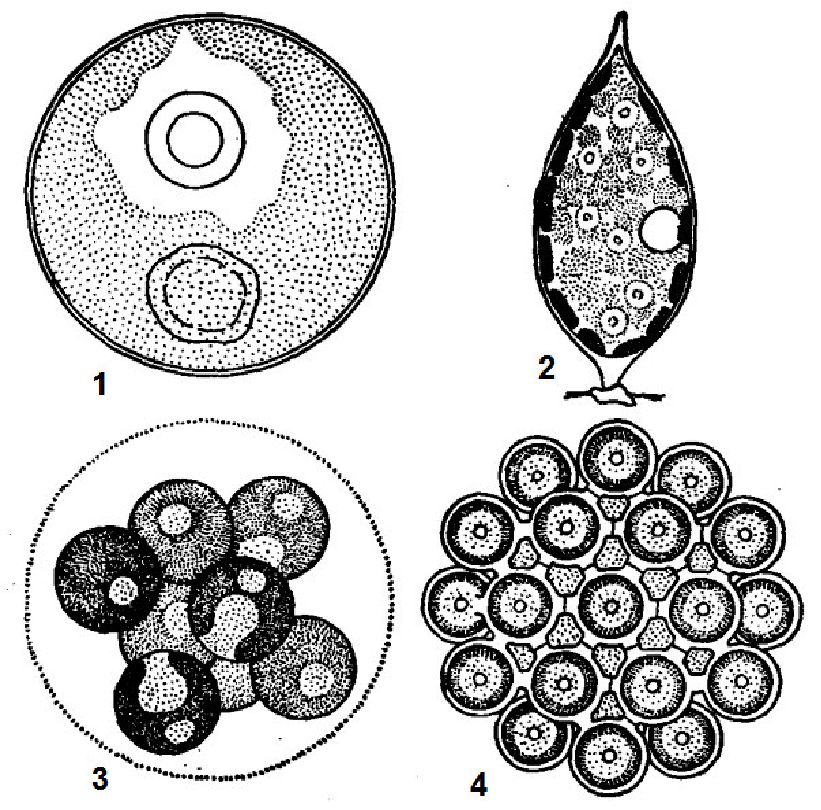
Рис. 6. 4. Кокоїдна структура у зелених водоростей:
1 - одиночна клітина хлоро-коку(СЫогососсит); 2 - одиночна клітина харціуму (Сйагасшт); 3 - колонія сфе-роцистиса (Sphaerосystis), утворена слизом; 4 - колонія, утворена зростанням клітин (Соеіазішт)
Кокоїдний тип структури широко розповсюджений майже у всіх відділах еукаріотичних водоростей (за винятком евгленових і рафі-дофітових).
Сарциноїдний тип структури є поєднанням кокоїдного зовнішнього вигляду із здатністю до вегетативного клітинного поділу, що відбувається в різних площинах, завдяки чому утворюються скупчення з багатьох клітин, що згодом легко розпадаються. Це одноклітинні або колоніальні організми з неполярною будовою клітин і міцною клітинною оболонкою. Сарциноїдний тип структури властивий зеленим і жовто-зеленим водоростям.
Поява здатності до вегетативного клітинного поділу була однією з найбільш значних подій в еволюції рослинного світу, з яким пов'язано виникнення багатоклітинних макроскопічних рослин.
Нитчастий (трихальний) тип структури має найхарактернішу особливість - ниткоподібне розташування нерухомих клітин, що утворюються в результаті клітинного поділу, що відбувається переважно в одній площині. Їм властива найважливіша властивість рослинних організмів — здатність до необмеженого росту і можливість виникнення багатоклітинних макроскопічних таломів.
У простих випадках таломи нитчастої структури складаються з морфологічно подібних клітин. У багатьох водоростей морфологічні відмінності часто мають клітини на ділянках стоншення нитки, кінцеві клітини. При прикріпленому способі життя нижня клітина перетворюється на безбарвний ризоїд або стопу, позбавлену хлоропластів. Нитки можуть бути простими або розгалуженими, одно-або багаторядними, вільноживучими або прикріпленими, одиночними або об'єднаними в угруповання різного типу (рис. 6.5).

Рис. 6. 5. Нитчаста структура у синьозелених водоростей:
1 - найпростіша будова нитки з дифузним ростом у осциляторії (Oscillatoria); 2 -нитка з диференційованою основою у ендонеми (ЕМопета); 3 - верхівка нитки у ривулярії ^ітоіагіа), витягнута у волосок; 4 - інтеркалярний ріст у глеотрихії (ОІоеоігісШа); 5 - апікальний ріст на кінцях гілочок у сцитонеми (суіопета)
Нитчастий тип структури представлений серед синьозелених, зелених, червоних, жовтозелених, золотистих водоростей.
Різнонитчастий (гетеротрихальний) тип структури виник на основі нитчастого унаслідок морфологічної диференціації різних ділянок у зв'язку з пристосуванням їх до виконання різних функцій: прикріплення, опорної, асиміляційної, відтворення і ін.
Різнонитчастий талом складається переважно з горизонтальних ниток, що стелються по субстрату і виконують функцію прикріплення, і вертикальних ниток, що піднімаються над субстратом. Вони виконують асиміляційну функцію й несуть органи розмноження.
У деяких водоростей вертикальні нитки диференційовані на міжвузля і вузли, від яких відходять кільця бічних гілочок. Функцію прикріплення до субстрату виконують безбарвні ризоїди. Така будова спостерігається у харових водоростей.
Різнонитчаста структура спостерігається у багатьох зелених, бурих, червоних, деяких синьо-зелених, жовтозелених і золотистих водоростей (рис. 6.6).
Виникнення і розвиток диференціації нитчастого талому було важливою подією в морфологічній еволюції водоростей, що зумовило появу нових великих таксонів, завоювання нових екологічних ніш і послужило відправним пунктом для розвитку паренхіматозного і псевдопаре-нхіматозного типів структури.
Псевдопаренхіматозний тип структури. Характерною особливістю псевдопаренхіматозного типу структури є утворення великих об'ємних багатоклітинних таломів, у результаті зрощення ниток розгалуженого різнонитчастого талому, що часто супроводжується морфофункціональною диференціацією «тканин».
Оскільки останні за способом утворення відрізняються від справжніх тканин, їх називають несправжніми тканинами. Псевдопаренхіматозний тип структури — тупикова гілка в морфологічній еволюції водоростей.

Рис. 6. 6. Різнонитчаста структура у синьозеленої водорості фішерели (РІ8еЬеге11а)
Паренхіматозний (тканинний) тип структури. Подальша еволюція нитчастого і різнонитчастого таломів була пов'язана з виникненням здатності до поділу клітин первинних ниток в різних площинах з утворенням паренхіматозних таломів. Здатність до необмеженого росту і поділу клітин у різних напрямах призвела до утворення об'ємних макроскопічних таломів (рис. 6.7). В межах паренхіматозного типу структури спостерігається поступове ускладнення таломів від простих недиференційованих пластинок з дифузним ростом (Рогрйуга, Шяа) до складних таломів із тканинами, що виконують асиміляційну, провідну, запасаючу, опорну функції, і примітивними багатоклітинними органами.
Тканинна структура представлена у бурих, червоних і зелених водоростей. Виникнення її було великою еволюційною подією. Воно привело до формування різноманітних морфологічних варіантів на даному рівні організації, до завоювання нових біотопів, закріпивши панування водоростей в рослинному світі водного середовища, подібно до домінування вищих рослин на суші. Паренхіматозний тип Рис. 6. 7. Паренхіматозна (пластинча- структури — найвищий зі всіх ста) структура у зеленої водорості пра- досягнутих в процесі еволюції зіоли (Pгаslоlа): однорядна нитка, що
розростається у одношарову пластинку ступенів морфологічної диференціації тіла водоростей.
Сифональний тип структури характеризується відсутністю всередині талому клітинних перегородок за наявності великої кількості органоїдів. Саме тіло водорості досягає порівняно великого розміру і певного диференціювання. Перегородки можуть з'являтися лише випадково, при пошкодженні талому або при утворенні репродуктивних органів. У обох випадках процес утворення перегородок відрізняється від способів їх формування в багатоклітинних рослинах.
Цей напрям морфологічної еволюції водоростей виявився тупиковим, таким, що не витримав конкуренції з магістральним еволюційним шляхом, що виник на основі вегетативного поділу клітинСифональний тип структури представлений у деяких зелених і жовтозелених водоростей (рис. 6.8).
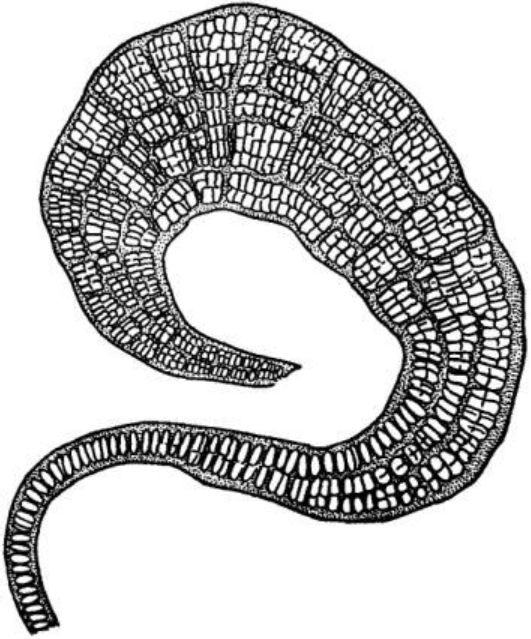

Рис. 6. 8. Сифональна структура у морської зеленої водорості каулер-пи (Саиіегра) (частина талому)
Сифонокладальний тип структури. Основною ознакою сифо-нокладального типу структури є здатність до утворення з первинного неклітинного талому в результаті сегрегативного поділу складно влаштованих таломів, що складаються з первинно багатоядерних сегментів. Характерною особливістю сегрегативного поділу, що лежить в основі формування сифонокладального талому, є роз'єднаність процесів мітозу і цитокінезу.
Сифонокладальний тип структури відомий лише у зелених водоростей. Цей напрям морфологічної еволюції, що виник на базі сегрегативного поділу, хоч і призвів до формування нових таксонів і освоєння нових біотопів і екологічних ніш, але все-таки виявився ще однією тупиковою гілкою морфологічної еволюції рослин.
6.3. Екологічні групи водоростей
Для водоростей як фототрофних організмів основною умовою існування є наявність світла, джерел вуглецю і мінеральних речовин, а основним середовищем життя для більшості водоростей є вода. Крім того, на життя водоростей мають великий вплив і інші фізико-хімічні чинники зовнішнього середовища — хімічний склад субстрату, температура і т. д., які дуже відрізняються в різних місцях. Пристосованість водоростей до різних екологічних умов справді величезні. Водорості зустрічаються в різних водних, наземних і ґрунтових біотопах по всій земній кулі. Через широку пристосованість до зовнішніх умов, водорості розподіляються в різноманітні екологічні угруповання, що пристосовані до певної амплітуди екологічних чинників.
У межах кожної водойми водорості можуть населяти товщу води, вільно плавати в ній, складаючи угруповання планктону, або поселятися на дні, утворюючи бентос.
Проте водорості можуть жити не лише у водоймах. За наявності періодичного зволоження багато водоростей успішно розвивається на різних наземних предметах — скелях, корі дерев і т.д. Сприятливим середовищем для розвитку водоростей служить також ґрунт. Таким чином, виділяють різні екологічні групи цих рослин: планктонні і бентосні, наземні і ґрунтові водорості, водорості снігу і льоду, водорості солоних водойм, водорості, що живуть у вапняковому субстраті. При проведенні флористичних досліджень зазвичай застосовують більш загальні класифікації, наприклад, водорості не водних місцепроживань ділять на 4 екологічні угруповання: 1) повітряні водорості (аерофітон); 2) наземні водорості (геофітон); 3) грунтові водорості (едафон) і 4) водорості усередині вапняного субстрату (ендолітофітон). У зарубіжних роботах все різноманіття неводних поселень водоростей відносять до двох типів: аерофільні і грунтові водорості, причому розростання водоростей на грунті відносять або до першого, або до другого типу.
Водорості водних місцезростань (гідрофітон). Водне середовище неоднорідне і має ряд особливостей, до яких пристосувалися різні живі організми.
Залежно від поширення у водному середовищі гідробіонтів поділяють на тих, які живуть у товщі води (планктон, нектон і нейстон) і тих, які поширені на дні водних об’єктів (бентос). Екологічні групи включають організми різного систематичного положення (водорості і вищі водні рослини, тварини, гриби, бактерії), які мають пристосування до життя у певній екологічній зоні.
Організми планктону не здатні протистояти течії, але мають спеціальні пристосування для зависання у товщі води - газові і ліпідні включення, які легші за воду і дозволяють зменшити загальну вагу тіла; збільшення площі поверхні тіла за рахунок зменшення розмірів, сплющування і видовження, формування виростів або щетинок, що збільшує тертя об воду (рис. 6.9). За систематичною ознакою планктон підрозділяють на фітопланктон, бактеріопланктон і зоопланктон. Отже, фітопланктоном називають сукупність вільноплаваючих (у товщі води) дрібних, переважно мікроскопічних, рослин, основну масу яких складають водорості. Відповідно кожен окремий організм зі складу фітопланктону називають фітопланк-тером.

Рис. 6. 9. Пристосування до планктонного способу життя у водоростей із різних систематичних груп
1-4 - шипуваті форми (1 - маломонас (Mallomonas), одноклітинна джгутикова золотиста водорість з окремими лусочками на оболонці з відростками; 2 - колоніальна зелена водорість иедіаструм (Pediastrum) з шипами на крайових клітинах; 3 -одноклітинна зелена водорість голенкінія ^оіепкіпіа) з шипами, які всіюють оболонку; 4 - одноклітинна діатомея коретрон (Согеїтоп) з трьома віночками відростків на панцирі); 5-6 - парашутні форми (5 - зірчаста колонія діатомеї астеріонели (Astеriоnеllа) зі слизистими тяжами між клітинами, що утворюють парашут; 6 -одноклітинна діатомея планктоніелла (Ріапкіопіеііа) з плоскою формою панциру)
Полегшується маса клітини також шляхом зменшення її розмірів: розміри клітин у планктонних видів, як правило, менші, ніж у близькоспоріднених донних водоростей. У планктоні широко розповсюджені і найдрібніші організми, розміром у декілька мікрометрів, які створюють так званий нанопланктон.
Фітопланктон у житті великих водойм виконує ту ж роль, що і рослини на суші, тобто виробляє первинну органічну речовину, за рахунок якої прямо або опосередковано (через ланцюг живлення) існує решта живих організмів.
Видовий склад фітопланктону відрізняється у різних водних об’єктах, а також в одному і тому ж водному об’єкті у різні сезони року. Він залежить від гідрофізичного і гідрохімічного режиму.
Прісноводний фітопланктон має величезну різноманітність зелених і синьозелених водоростей. Особливо численні серед зелених одноклітинні і колоніальні вольвоксові і протококові: види хламідомонад (Chlamydomonas), гоніуму (Gonium), вольвоксу (Volvox), педіаструму (Pediastrum), сценедесмуса (Scenedesmus), ооцистіса (Oocystis), сфероцистіса (Sphaerocystis) і ін. Серед синьозелених численні види анабени (Anabaena), мікроцистіса (Microcystis), афа-нізоменона (Aphanizomenon), глеотрихії (Gloeotrichia) і ін.
Видова різноманітність діатомових тут нижча, ніж в морях (якщо не враховувати велику різноманітність тимчасово планктонних видів); за продуктивністю на одиницю поверхні води роль діатомових в прісних і морських водах у середньому співрозмірна.
Характерний у морському фітопланктоні рід хетоцерос (Chaetoceros) в озерах і ставах зовсім відсутній, а численні в морях ризосоленії (Rhizosolenia) у прісних водах представлені лише декількома видами.
У прісних водах відсутні кремнієві джгутикові і дуже рідкісні коколітофориди, зате деякі інші джгутикові представлені різноманітно і нерідко у великій кількості. В основному це хризомонади — види дінобріону (Dinobryon), маломонасу (Mallomonas), уроглени (Uroglena) і ін., а також евгленові — евглена (Euglena), трахеломо-нас (Trachelomonas) і факус (Phacus); перші переважно в холодних водах, а другі — в теплих.
У кожен сезон переважаючого розвитку набуває одна з груп водоростей (діатомові, синьозелені, золотисті, евгленові або зелені), а в періоди інтенсивного розвитку часто домінує лише один вид. Особливо це характерно для прісноводних водойм. Так, взимку під кригою фітопланктон дуже бідний або майже відсутній через нестачу світла. Вегетаційний розвиток водоростей планктону як угруповання починається навесні, коли рівень сонячного випромінювання стає достатнім для фотосинтезу водоростей навіть під кригою. В цей час з'являються досить численні дрібні джгутикові — евгленові, динофітові, золотисті, діатомові. У період від скресання криги до встановлення температурної стратифікації, що зазвичай буває при прогріві верхнього шару води до 10—12°С, починається бурхливий розвиток холодолюбних діатомових. Влітку при температурі води вище 15°С максимальний розвиток синьозелених, евгленових і зелених водоростей, аж до можливого «цвітіння» води (рис. 6.10).

Рис. 6. 10. Планктонні синьозелені водорості з газовими вакуолями в клітинах, які викликають «цвітіння» води:
1 - дві колонії мікроцистіса (Microcystis aeruginosa), утворені безструктурним слизом; 2 - колонія вороніхії (Woronichia naegeliana) з штрихуватим зовнішнім слизом; 3,4 - афанізоменон (Aphan^omenon flos-aque) (3 - лусочки з ниток в натуральну величину, 4 - ділянки ниток при великому збільшенні); 5 - зібрані в клубочок нитки анабени (Anabaena lemmermannii); 6 - плаваючі окремі нитки анабе-HH(Anabaena scheremetievii); 7,8 - колонія і окрема нитка глеотрихії (Gloeotrichia echinulata) при різному збільшенні
Морський фітопланктон складається в основному з діатомових і динофітових водоростей. З діатомових особливо численні представники класу центричних. Дуже різноманітний у морському фітопланктоні динофітові водорості. Ця група і в прісноводному фітопланктоні досить різноманітна, але налічує менше число видів, ніж у морському, а деякі роди представлені тільки в морях (Dinophysis, Goniaulax і ін.). Численні в морському фітопланктоні також вапнякові джгутикові — Coccolithophoridophycidae, представлені в прісних водах лише кількома видами, і кремнієві джгутикові, або силі-кофлагелляти, що зустрічаються виключно в морському планктоні.
Термін «планктон» за першим визначенням означав сукупність організмів, завислих у товщі води. Зараз до планктону відносять і організми нейстону.
Нейстон - сукупність гідробіонтів, які живуть на межі з повітряним середовищем біля плівки поверхневого натягу води. Одні з організмів нейстону існують над плівкою води - епінейстон, інші -під плівкою - гіпонейстон.
Великі концентрації нейстоних організмів спочатку були виявлені в дрібних водоймах (ставах, ямах, в невеликих затоках озер) у тиху погоду при спокійній поверхні води. Пізніше різноманітні ней-стонні організми, в основному дрібні тварини, були знайдені і в великих водоймах, зокрема в морях. До складу прісноводих водоростей нейстону входять види різних систематичних груп. Тут виявлені представники золотистих — хромуліна (Сйготиііпа), кремастохри-зис (Kгemаstоchгysіs); з евгленових — евглена (Еиріепа), трахело-монас (Tгachеlоmоnаs), а також деякі зелені — хламідомонада (Chlаmydоmопаs), кремастохлорнс (Kгеmаstоchlогіs), окремі види жовтозелених і діатомових водоростей.
У деяких видів нейстонних водоростей існують характерні пристосування до існування біля поверхні води. Наприклад, у видів нау-тококкуса (Каиіососсш) є слизисті парашути, що утримують їх на поверхневій плівці. У кремастохризиса для цього служить лускатий парашут; у одного виду із зелених водоростей такий мікроскопічний парашут виступає над плівкою поверхневого натягу у вигляді конусовидного ковпачка (рис. 6.11).
Рис. 6. 11. Водорості нейстону:
1 - лусочка крематохризису (КгетазіосЬгузіз), що плаває на поверхні води, з декількох «па-рашутиків» з клітинами, що висять під ними; 2 - конічні «пара-шутики» кремастохлорісу (КгетазіосШогіз) на поверхні води з підвішеними до них клітинами
В окремих випадках нейстонні організми розвиваються в такій кількості, що вкривають воду суцільною плівкою. Часом і планктонні водорості (особливо синьозелені) у період масового розвитку спливають до поверхні води, утворюючи величезні скупчення.
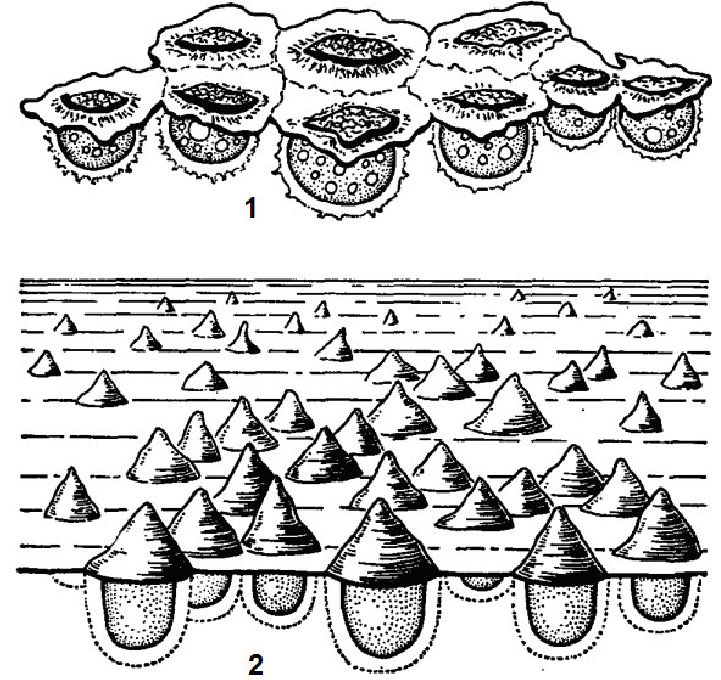
До екологічної групи плейстону (від грец. pleusis — плавання, ріео — пливу) відноситься сукупність водних організмів, що тримаються на поверхні води або напівзанурених у неї (тобто мешкають одночасно у водному і повітряному середовищі). Як правило, до складу плейстону відносять тваринні організми, а з рослин, наприклад, саргасові водорості.
До бентосних (донних) водоростей належать рослини, пристосовані до існування в прикріпленому стані на дні водойм і на різноманітних предметах і організмах, що знаходяться у воді. Залежно від місця росту серед бентосних водоростей розрізняють:
1) епіліти, які ростуть на поверхні твердого грунту (скелі, каменях і т. д.);
2) епіпеліти - населяють поверхню пухких грунтів (пісок, мул);
3) епіфіти - живуть на поверхні інших водних рослин;
4) ендоліти, або свердлячі водорості, що угвинчуються у вапняний субстрат (скелі, черепашки молюсків, панцирі ракоподібних);
5) ендофіти - поселяються в таломах інших водоростей, але, на відміну від паразитичних видів, мають нормальні хлоропласти;
6) паразити, що живуть в таломах інших водоростей, хлоропласти в клітинах не виражені.
Іноді водорості, що ростуть на предметах, споруджених людиною у воді (судна, плоти, буї), відносять до групи перифітону. Виділення цієї групи обґрунтовується тим, що ці організми (водорості і тварини) живуть на рухомих предметах або обтікаються водою. Крім того, ці організми віддалені від дна і, отже, знаходяться в умовах іншого світлового і температурного режимів, в інших умовах надходження біогенних речовин, джерелом яких служать донні відкладення. Іноді виділення перифітону обґрунтовують ще і практичними міркуваннями: це обростання, які можуть заподіювати практичний збиток, — зменшувати швидкість суден, засмічувати водозабірні отвори і трубопроводи. У перифітоні розвиваються водорості з різних систематичних груп (переважно зелені, синьозелені, діатомові і жовтозелені), що зазвичай володіють спеціальними органами (органоїдами) прикріплення у вигляді підошви, стопи, слизистих ниток. Наприклад, види родів улотрикс (Шоїйгіх), едогоніум (Оейоропіиш), афанохета (Арйапосйаеіе), гідрурус (Нуйгигщ) і ін. Часто зустрічаються також синьозелені водорості, що прикріплюються до підводних предметів за допомогою слизу - види родів лін-
гбія (Lyngbya), осциляторія (Oscillatoria), ривулярія (Rivularia), гле-отрихія (Gloeotrichia), носток (Nostoc) і ін., і діатомові, що щільно прилягають до субстрату безпосередньо нижньою стулкою зі швом - наприклад, види роду коконеїс (Cocconeis) і ін.
Між епілітами, епіпелітами і епіфітами часто немає чіткої грані, особливо це стосується мікроскопічних бентосних водоростей. Правда, існують види, які живуть лише на інших водоростях і лише на певних видах. Наприклад, полісифонія шерстиста (Polysiphonia lanosa) росте виключно на аскофіллюмі Ascophyllum nodosum).
Для росту бентосних водоростей особливо важливе світло. Але ступінь його використання залежить від інших екологічних чинників: температури, вмісту біогенних речовин, кисню і неорганічних джерел вуглецю, а головне — від темпів надходження цих речовин в тіло рослини, що залежить від концентрації речовин і швидкості руху води. Як правило, місця з інтенсивним рухом води відрізняються значним розвитком бентосних водоростей.
Інтенсивному розвитку бентосних водоростей сприяє помірний вміст у воді біогенних речовин, джерелом яких служать поверхневий стік і донні відкладення. У прісноводних екосистемах такі умови створюються в неглибоких ставах, у прибережній зоні озер, у річкових затоках, а в морях — в дрібних затоках. Якщо в таких місцях є достатнє освітлення, твердий грунт і помірна течія, то створюються оптимальні умови для життя фітобентосу. За відсутності руху води і при недостатньому забезпеченні біогенними речовинами бентосні водорості ростуть погано.
Крім абіотичних чинників, розвиток бентосних водоростей залежить від присутності рослиноїдних тварин — морських їжаків, черевоногих молюсків, ракоподібних, риб. У тропічних морях у деяких місцях риби повністю виїдають м'які зелені, бурі і червоні водорості. Черевоногі молюски, повзаючи по дну, поїдають мікроскопічні водорості і дрібні проростки макроскопічних видів.
Переважаючими бентосними водоростями континентальних водойм є діатомові, зелені, синьозелені і жовтозелені нитчасті водорості, прикріплені або не прикріплені до субстрату. Це види таких родів: навікула (Navicula), діатома (Diatoma), гіросигма (Gyrosigma), кладофора (Cladophora), едогоніум (Oedogonium), улотрикс (Ulothrix), спірогіра (Spirogyra), мужоція (Mougeotia), зигнема (Zygnema), осциляторія (Oscillatoria), лінгбія (Lyngbya), формідіум
(Phormidium), трибонема (Tribonema), вошерія (Vaucheria) і ін.
Основні бентосні водорості морів і океанів — бурі і червоні, іноді зелені макроскопічні, прикріплені сланеві форми. Це види родів бангія (Bangia), фукус (Fucus), порфіра (Porphyra), філофора (Phyllophora), ентероморфа (Enteromorpha), ламінарія (Laminaria) і ін. Всі вони можуть обростати дрібними діатомовими, синьозеле-ними і іншими водоростями.
Водорості снігу і льоду. Серед кріофільних водоростей переважають зелені, синьозелені і діатомові. Розвиваючись в масовій кількості, вони можуть викликати зелене, жовте, блакитне, червоне, коричневе, буре або чорне «цвітіння» снігу або льоду. Зелене забарвлення снігу викликає рафідонема (Raphidonema nivale), червоне — хламідомонада (Chlamydomonas nivalis), коричневе — анцилонема (Ancylonema nordenskioeldii). Ці водорості знаходяться в поверхневих шарах снігу або льоду та інтенсивно розмножуються при температурі близько 0°С. Лише небагато з них мають стадії спокою, більшість позбавлені будь-яких спеціальних морфологічних пристосувань для переживання низьких температур.
Аерофільні водорості (аерофітон). Основним життєвим середовищем аерофільних водоростей є повітря. Число видів, виявлених в аерофільних угрупованнях, наближається до 300. Типові місця поширення — поверхня різних твердих субстратів (скелі, камені, кора дерев і т. д.). Залежно від рівня вологості їх підрозділяють на дві групи: повітряні водорості, що мешкають в умовах лише атмосферного зволоження і, отже, зазнають постійної зміни зволоження і висихання; водно-повітряні водорості, що зазнають постійного зволоження (під бризками водоспаду, прибою і т. д.).
Водорості, що мешкають в умовах виняткового атмосферного зволоження, вимушені часто переходити із стану надлишкового зволоження (наприклад, після зливи) в стан мінімальної вологості в посушливі періоди, коли вони висихають настільки, що можуть бути розтерті в порошок.
Аерофільні угруповання переважно зазнають постійних коливань температури. Вдень вони сильно прогріваються, вночі охолоджуються, взимку промерзають. До несприятливих умов існування пристосувалися порівняно небагато зелених і синьозелених водоростей, представлених мікроскопічними одноклітинними, колоніальними і нитчастими формами, і, в значно меншій мірі, діатомових.
Аерофільні форми відомі і серед червоних водоростей - види родів порфіридіум (Porphyridium), родоспора (Rhodospora); вони зустрічаються на каменях, старих стінах оранжерей і т.д. При масовому розвитку аерофільних водоростей, вони мають вигляд порошкоподібних або слизистих нальотів, м'яких або твердих шкірочок.
На корі дерев найчастіше поселяються зелені водорості (представники родів хлорела (Chlorella), хлорокок (Chlorococcum) і ін. Синьозелені і діатомові зустрічаються на деревах значно рідше.
Існує дуже своєрідна група водоростей, що ростуть на мохах. У ній переважають бріофільні діатомові, зустрічаються специфічні види зелених (представники родів мезотеніум (Mesotenium), пеніум (Penium) і ін.) і жовтозелених водоростей.
На поверхні голих скель розвиваються діатомові і деякі, переважно одноклітинні, зелені водорості, але найбільш звичайні для цих місцезростань представники синьозелених водоростей (види родів глеокапса (Gloeocapsa), хамесифон (Chamaesiphon), толіпотрикс (Tolypothrix), сцитонема (Scytonema) і ін.). Водорості і супутні бактерії утворюють «гірський загар» (наскельні плівки і шкірочки). Особливо рясно розростаються водорості на поверхні вологих скель. Вони утворюють плівки і нарости різного кольору - яскраво-зелені, золотисті, бурі, синьо-зелені, коричневі, майже чорні, залежно від видів, що їх утворюють.
Ґрунтові водорості (едафон). Основним середовищем існування едафофільних водоростей є поверхня і товща ґрунту.
Непрозорість ґрунту має вирішальний вплив на розвиток водоростей. Інтенсивний розвиток водоростей можливий лише в межах проникнення світла. У цілинних ґрунтах це поверхневий шар завтовшки до 1 см, в оброблюваних ґрунтах він дещо товстіший. Деякі водорості переходять у темряві до гетеротрофного живлення. Багато водоростей зберігаються в ґрунті у стані спокою.
Для попередження висихання ґрунтові водрості мають здатність до утворення слизу — слизистих колоній, чохлів і обгорток. Завдяки наявності слизу водорості швидко поглинають воду при зволоженні і запасають її, уповільнюючи висихання.
Характерною рисою ґрунтових водоростей є «ефемерність» їх вегетації — здатність швидко переходити зі стану спокою до активної життєдіяльності і навпаки. Вони також здатні витримувати різкі коливання температури ґрунту. Діапазон виживання деяких видів лежить у межах від -200 до +84 °С і вище. Наземні водорості складають важливу частину рослинності Антарктиди. Вони забарвлені майже в чорний колір, тому температура їх тіла вища за температуру навколишнього середовища.
Переважна більшість ґрунтових водоростей — мікроскопічні форми, проте часто їх можна побачити на поверхні ґрунту неозброєним оком. Наприклад, в грунтах степової зони України широко розповсюджений носток (Nostoc commune), що створює на поверхні ґрунту плівку темно-зеленого або (в сухий сезон) шкірку грифельно-чорного кольору. Масовий розвиток мікроскопічних форм викликає позеленіння схилів ярів і узбіччя лісових доріг.
За систематичним складом ґрунтові водорості досить різноманітні. Найбільшою кількістю видів представлені синьозелені і зелені водорості. Меншу різноманітність мають представники відділів жо-втозелених і діатомових водоростей. Ще рідше зустрічаються евгленові, золотисті, динофітові і червоні.
Літофільні водорості (ендолітофітон). Основним життєвим середовищем літофільних водоростей є непрозорий щільний вапняковий субстрат. Літофільні водорості живуть у глибині твердих порід певного хімічного складу. Залежно від фізіологічних особливостей, розрізняють дві групи літофільних угруповань: свердлячі водорості, що активно угвинчуються в кам'янистий субстрат і заселяють дрібні ходи та пори, утворені ними в кам'янистій породі; туфоут-ворюючі водорості, що відкладають навколо свого тіла вапно і мешкають у периферичних шарах цього середовища, в межах доступності світла і води. У міру наростання відкладень ці ценози поступово відмирають.
Водорості-паразити. Безбарвні паразитичні види, які мешкають у кишечнику червів, копепод, амфібій, на зябрах риб, відомі серед евгленових і динофітових водоростей (види родів бластодиніум (Blastodinium), синдініум (Syndinium), іхтіодиніум (Ichtyodinium), трихомонас (Trichomonas) і ін.)
Водорості-ендосимбіонти. Зелена водорість з роду картерія (Carteria), поселяється в епідермальних клітинах війчастого черва Convoluta roscoffensis, один вид роду хлорела (Chlorella) — у вакуолях деяких інфузорій, а види роду хлорокок (Chlorococcum) — у клітинах криптофітової водорості ціанофори (Cyanophora paradoxa). Ендосимбіонти зазвичай зазнають значних морфологічних змін у порівнянні з вільноживучими представниками того ж роду (клітинна оболонка редукується, будова джгутиків спрощується), проте вони не втрачають здатності до фотосинтезу і розмноження усередині клітин господаря.
6.4. Класифікація водоростей
Сучасна класифікація водоростей побудована на особливостях їх забарвлення. На тлі розвитку хлорофілу, пігментного компоненту всіх водоростей, відмінностях біохімічних особливостей і морфологічної будови утворилися пігментні групи: синьо-зелена, зелена, бура, жовта, червона. Є прості одноклітинні і багатоклітинні водорості, які мають складну будову, що нагадує будову вищих наземних рослин. Існують мікроскопічні форми і гіганти, розміри яких досягають десятків і навіть сотень метрів. Підводні ліси, що формуються морськими бурими ламінарієвими водоростями (алярії, ламінарії, макроцистіса, нереоцистіса, лесонії), нагадують джунглі. У посібнику прийнято такий поділ водоростей на відділи: Прокаріотичні водорості Відділ 1. Синьозелені водорості — Cyanophyta Еукаріотичні водорості Відділ 1. Евгленові водорості — Euglenophyta Відділ 2. Динофітові водорості — Dinophyta Відділ 3. Криптофітові водорості — Cryptophyta Відділ 4. Рафідофітові водорості — Raphidophyta Відділ 5. Золотисті водорості — Chrysophyta Відділ 6. Діатомові водорості — Bacillariophyta Відділ 7. Жовтозелені водорості — Xanthophyta Відділ 8. Червоні водорості — Rhodophyta Відділ 9. Бурі водорості — Phaeophyta Відділ 10. Зелені водорості — Chlorophyta Відділ 11. Харові водорості — Charophyta
Детальна характеристика відділів водоростей подана нижче.
6.4.1. Синьозелені водорості або ціанеї (Cyanophyta)
До відділу належать близько 2 тис. видів, широко розповсюджених у водних і неводних біотопах. Це найдавніші водорості, прокаріоти. Є одноклітинні і багатоклітинні, одиночні і колоніальні синьозелені водорості. Вони можуть бути прикріпленими і неприкріп-леними до субстрату, нерухомими або рухливими, проте джгутиків і війок ніколи не утворюють. Синьозелені водорості переважно мікроскопічні, але деякі колоніальні види утворюють скупчення розміром у декілька сантиметрів. Колонії водоростей іноді займають великі простори і продукують значну кількість слизу.
Синьозелені водорості переважно мають специфічний синьо-зелений колір. Проте їх забарвлення може сильно варіювати залежно від комбінації пігментів — бути майже зеленим, оливковим, жовтувато-зеленим і ін. Для одноклітинних ціаней характерна кокоїд-на форма будови тіла. Для багатоклітинних — нитчаста (трихома-льна), рідше різнонитчаста. Дуже рідко спостерігається певна тенденція до пластинчастого або об'ємного розташування клітин. Обов'язковою складовою частиною тіла особин гормогонієвих водоростей є трихом — ниткоподібне (нерозгалужене або розгалужене) утворення, що складається з одного, рідше з кількох рядів фізіологічно взаємозв'язаних клітин. Ниткоподібними у ціаней бувають і колонії одноклітинних форм. Плазматичний взаємозв'язок між клітинами в цьому випадку відсутній. У гормогонієвих водоростей утворюються «нитки», які складаються з одного або кількох трихо-мів у слизистому чохлі і можуть мати інші утворення (гетероцисти і ін.).
Справжніх ядер, хлоропластів і вакуолей з клітинним соком у клітинах ціаней немає. Нерідко зустрічаються газові вакуолі. Оболонка клітин багатошарова, з внутрішнім муреїновим шаром. З хлорофілів є лише хлорофіл а. Хлорофілу Ь і бактеріохлорофілу немає. Продуктом асиміляції є глікогеноподібний полісахарид.
Мітозу і мейозу у ціаней немає. Розмноження вегетативне, рідше власне нестатеве (екзо- і ендоспорами). Розмножуються поділом (дробленням) клітин, розпадом нитчастих форм на багатоклітинні ділянки - гормогонії. Розрив ниток проходить біля особливих більш товстостінних мертвих клітин - гетероцист. Статеве розмноження відсутнє. Деякі форми у несприятливих умовах утворюють спори, як і бактерії. При цьому потовщується оболонка, зменшується кількість води у цитоплазмі, утворюється велика кількість запасних поживних речовин. Спори можуть витримувати тривале висушування, а у сприятливих умовах - проростають у нову особину.
Ціанеї переважно мешкають у водоймах, багатих органічними речовинами. Є планктонні і бентосні форми. Зустрічаються у ґрунті, першими заселяють нові площі після вулканічних вивержень. Деякі живуть у гарячих джерелах, на снігах Арктики і Антарктиди.
Є три класи ціаней: хроококові, хамесифонові і гормогонієві.
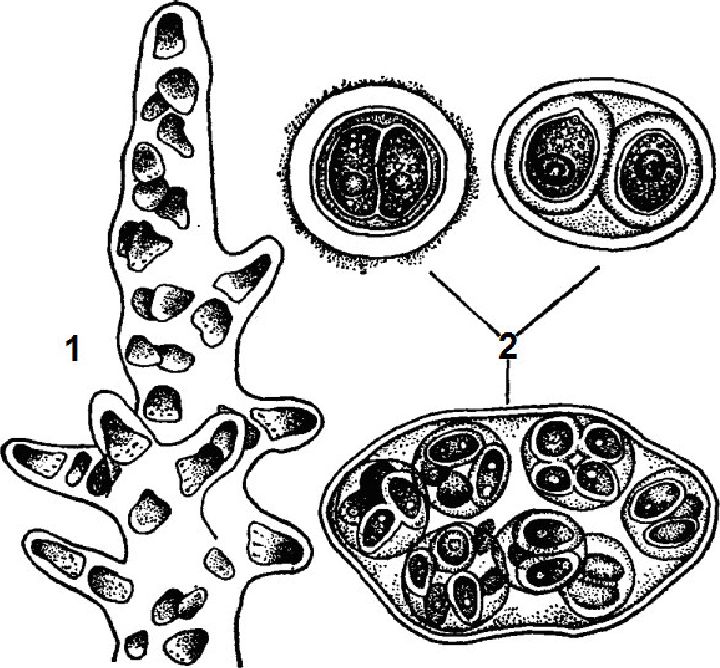
Представники класу хроококових (Скгоососсоркусеае) зустрічаються у вигляді окремих клітин або частіше утворюють колонії, оточені слизом. Колонії бувають «простими» і складними. «Прості» колонії складаються з окремих клітин «простих» ціаноїдів, сполучених між собою слизом. Складні колонії складаються з двох і більшої кількості ціаноїдів, які зовні схожі на «прості» колонії (рис. 6.12).

Рис. 6. 12. Хроококові (Chroococcophyceae): 1 - Merismopedia glauca, 2 -Chroococcus dimidiatus, 3 - Eucapsis, 4 - Gloeocapsa, 5 - Microcystis aeruginosa
Колоніальні і одноклітинні водорості одиночні або утворюють різноманітні угруповання: випадкові скупчення, своєрідні скупчення (агрегати) і поселення. Будова і форма колоній залежить від способу поділу клітин і особливостей утворення слизу. При поділі клітин у двох взаємно перпендикулярних площинах утворюються одношарові пластинчасті колонії (Merismopedia). Поділ клітин у трьох взаємно перпендикулярних площинах веде до утворення колоній у вигляді куба (Eucapsis). Якщо при поділі в трьох взаємно перпендикулярних площинах дочірні клітини зміщуються, то утворюються колонії типу глеокапси (Gloeocapsa).
У видів роду мікроцистіс (Microcystis), які часто зумовлють «цвітіння» континентальних водойм, поділ клітин відбувається в багатьох площинах, і тому клітини розташовуються безладно по всій товщині слизу, а форма колоній може бути найрізноманітнішою. Колоніальний слиз сполучає прості і складні ціаноїди колоній.
Цим водоростям властиве вегетативне розмноження. Найчастіше розмноження відбувається поділом колоній - фрагментацією (розрив на декілька частин), перешнуровуванням видовженої материнської колонії і її брункуванням.
Клас хамесифонові (Chamaesiphonophyceae) об’єднує одноклітинні, колоніальні та нитчасті форми, що утворюють нитчасті тало-ми, які переважно прикріплені до субстрату. Характерне утворення спор нестатевого розмноження.
Клітини в колоніях нерідко об’єднані у глеокапсоподібні групи або розташовані неправильно; часто вони дуже стиснуті і утворюють несправжню паренхіму.
Розмноження вегетативне і власне нестатеве (за допомогою екзо-і ендоспор). За рівнем організації хамесифонові займають проміжне положення між хроококовими і гормогонієвими водоростями.
Хамесифонові ростуть на рослинах або на каменях, віддають перевагу чистим і бідним на поживні речовини гірським струмкам, де їх таломи утворюють кольорові шкірочки або плями на каменях, черепашках і рослинах. Деякі види мешкають в морях.
В межах класу виділяють чотири порядки: плеврокапсові (Pleurocapsales), дермокарпові (Dermocarpales), сифононемові (Siphononematales) і ендонемові (Endonematales).
Гормогонієві водорості (Hormogoniophyceae) - це найбільший клас синьозелених водоростей, що включає більшу частину видів, які характеризуються нитчастою (трихомальною) будовою тіла і здатністю до утворення гормогоніїв - спеціальних фрагментів та-ломів, які здатні до активного руху і проростання в нові особини.
Водорості багатоклітинні, «прості» або колоніальні (з багатоклітинними ціаноїдами).
До особливих клітин належать гетероцисти і акінети, які утворюються з вегетативних клітин.
Акінети — це особливі одноклітинні утворення, які виконують функцію збереження життя в несприятливих умовах, а у деяких випадках також функцію розмноження. Оболонка акінет потовщена.
Розмноження у гормогонієвих водоростей, як правило, вегетативне (випадковою фрагментацією, гормогоніями, акінетами і ін.). Ендоспори виявлені лише у окремих представників. Статеве розмноження і джгутикові стадії відсутні.
Найхарактерніше для гормогонієвих водоростей розмноження за допомогою гормогоніїв, які у багатьох форм утворюються по всій довжині трихома шляхом розпадання його на частини. Часто гормогонії утворюються в певних частинах нитки (наприклад, на верхівці) і є відносно диференційованими утвореннями.
Найхарактернішими для водойм України є роди осциляторія, анабена, афанізоменон, носток.
Особини осциляторії (Oscillatoria) складаються з одного ряду морфологічно однотипних (часом за винятком кінцевих) клітин, поодинокі або утворюють переважно плівчасті структури. Осциляторія має нерозгалужені нитки (рис. 6.13), для яких характерний повільний і плавно хвилястий (або осцилюючий) рух.

Рис. 6. 13. Осциляторія (Oscillatoria): а — будова клітини (1 — центроплазма, 2 — хроматоплазма); б, в — осциляторія
Розмноження у осциляторі! вегетативне. Є понад 100 видів, поширених по всій земній кулі, в Україні близько 70 видів. Деякі види спричинюють «цвітіння» води. Найхарактернішим видом для водойм України є осциляторія (OsdПatoria ^ш^).
Рід анабена (АпаЬаепа) включає водорості одиночні або у вигляді плівчастих дерновинок (рис. 6.14). Розмноження уривками ниток і акінетами. Є близько 100 видів, поширених у прісних і солоних континентальних водоймах, опріснених ділянках морів, гарячих джерелах, на ґрунті. Здатні фіксувати молекулярний азот. Викликають «цвітіння» води. Найхарактернішою для водойм України є анабена (АпаЬаепа Поз-ациае).
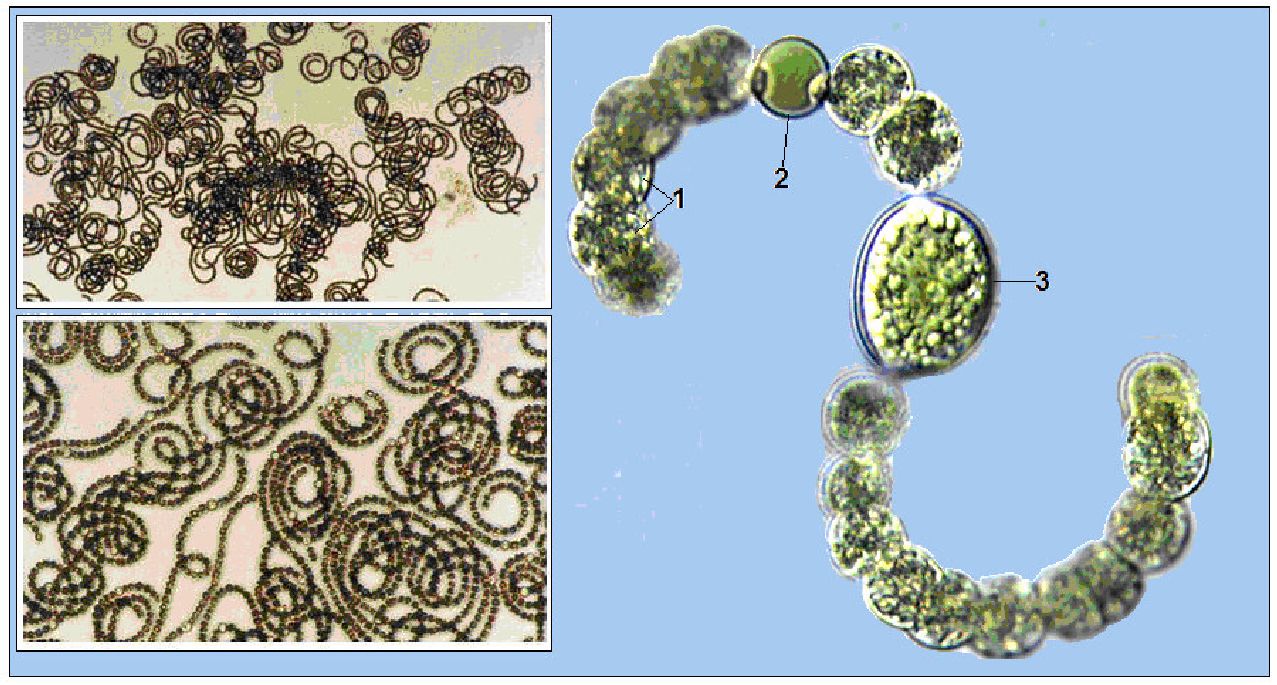
Рис. 6. 14. АпаЬаепа зрігоійез:
1 - вегетативні клітини, 2 - гетероциста, 3 - спора (акінета)
Рід афанізоменон (АрЬапігошепоп) включає 10 видів. Трихоми у водоростей з цього роду прямі або зігнуті, одиночні або сполучені у вільноплаваючі пучки, що досягають значних розмірів. Клітини в середній частині трихомів короткоциліндричні, з газовими вакуолями, на кінцях видовжені і звужені, часто безбарвні. Гетероцисти і спори інтеркалярні, різноманітної форми. Найпоширеніший вид — Aphamzomenon і^-а^ае з циліндричними спорами, звичайний збудник «цвітіння» води в евтрофних водоймах.
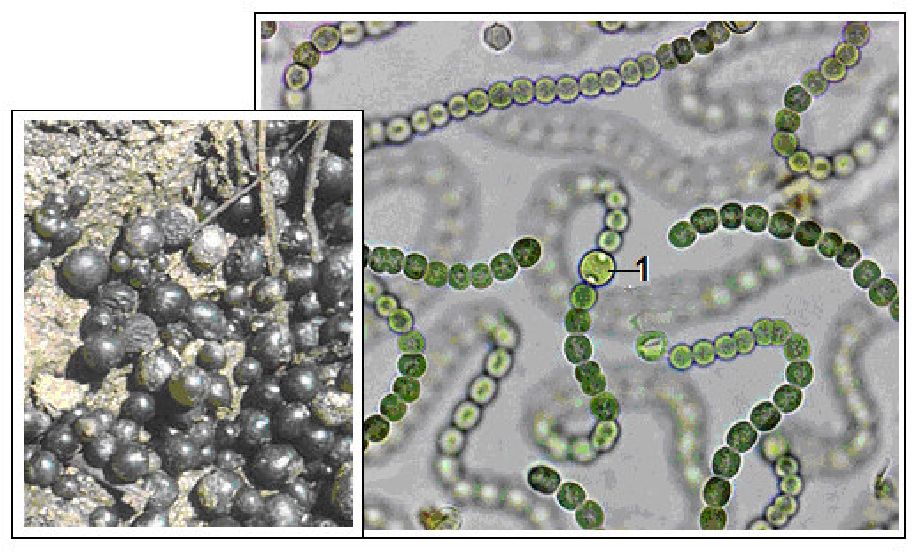
Рис. 6. 15. Носток сливовидний (Nostoc pruniforme):
1 - гетероциста
До роду носток (Nоstоc) відносяться водорості, які утворюють колонії — кулясті (діаметром до 5 см) (рис. 6.15), шкірковидні або кущисті у вигляді дерновинок (діаметром 0,5 м і більше). Трихоми з гетероцистами. Розмноження уривками ниток і акінетами. Є близько 40 видів, широко розповсюджені майже у всіх кліматичних зонах. Мешкають у прісних водоймах, на ґрунті, в кореневих потовщеннях деяких вищих рослин (саговник, конюшина); це компонент лишайників. Здатні до азотфіксації. У країнах Східної Азії вживаються в їжу.
Рід спіруліна (Spirulina) досить близький до осциляторії, але відрізняється спіральними трихомами. У дрібних форм поперечні перегородки бувають зовсім непомітними. Широко розповсюджені Spirulina jenneri і S. major. Тропічний вид S. platensis вирощують як об’єкт масової культури.
Значення синьозелених водоростей. Синьозелені водорості відіграють важливу роль у природі і дуже важливі в господарському відношенні. Завдяки своїй здатності засвоювати вільний азот з атмосфери і значній стійкості до дії несприятливих чинників багато ціаней живуть в умовах, непридатних для розвитку еукаріот. Вони поселяються на голих скелях, продуктах виверження вулканів та інших безплідних субстратах і беруть участь у процесі ґрунтоутворення - засвоюють вільний азот з повітря і збагачують ним ґрунт.
У результаті їх життєдіяльності помітно змінюються фізико-хімічні умови навколишнього середовища, що позначається на функціонуванні екосистем. Біомаса, утворена планктонними ціанеями в період інтенсивного «цвітіння» води, може складати 80—90 % біомаси всіх водоростей. Червоне море отримало свою назву через інтенсивний розвиток червоної осциляторії (Oscillatoria erythraea).
Проте інтенсивний розвиток синьозелених водоростей спостерігається далеко не у всіх сучасних біотопах. У розвинених фітоценозах вони поступаються вищим рослинам, а за певних умов навіть відсутні (при pH нижче 4-5, у швидких гірських річках і ін.).
Синьозелені водорості належать до найдревніших організмів, їх викопні рештки і продукти життєдіяльності виявлені в породах, що утворилися 2,3 (а може й 2,7—3,2) млрд. років тому, тобто ще в архейську еру. Вважають, що перші на Землі (докембрійські) екосистеми складалися лише з прокаріотичних організмів, зокрема ціаней. Інтенсивний розвиток ціаней у докембрійських морях мав величезне значення для розвитку життя на Землі, і не лише через накопичення ними органічної речовини, але і у зв'язку із збагаченням первинної атмосфери киснем.
Господарське значення ціаней пов'язане з їстівністю деяких їх представників (Nostoc pruniforme, Spirulina platensis і ін.), можливістю використання азотфіксуючих форм для підвищення родючості ґрунтів, необхідністю запобігання надмірному «цвітінню» води у водоймах, нерідко обумовлюваного токсичними формами, такими як мікроцистіс (Microcystis aeruginosa), анабена (Anabaena flos-aquae), афанізоменон (Aphanizomenon flos-aquae) і ін. Розробляються способи масового (промислового) культивування деяких видів ціаней, наприклад Spirulina platensis. Є можливість використання азотфіксуючих форм синьозелених для підвищення родючості грунтів. У Японії з метою отримання «живого добрива» у напіввиробни-чих умовах вирощують толіпотрикс (Tolypothrix tenuis). У В'єтнамі як добриво для полів здавна використовують водну папороть азоллу (Azolla), у симбіозі з якою живе анабена (Anabaena azollae).
6.4.2. Евгленові водорості (Euglenophyta)
Евгленові водорості є своєрідною групою нижчих рослин, що перебуває на межі рослинного і тваринного світу. Відділ об'єднує близько 1 тис. видів мікроскопічних (4-500 мкм), одноклітинних, переважно джгутикових, еукаріотичних водоростей зеленого, зрідка червоного кольору. Є багато безбарвних евгленових водоростей.
Це одноклітинні організми з монадним типом структури, мають один або декілька джгутиків, за винятком невеликої групи безджгу-тикових евгленових, а також форм, що ведуть переважно нерухомий спосіб життя і створюють колонії. Форма клітин різноманітна,
але найпоширенішою вважається веретено видна, або, як її часто називають, евгленоподібна, добре пристосована до руху у воді. Клітини без целюлозних оболонок. Під плазмалемою розташована пе-лікула - це щільний і еластичний шар протопласту білкової природи. Види з тонкою і гнучкою пелікулою не мають постійної форми, їм властива мінливість форми тіла. Під пелікулою у багатьох евгленових є слизові тільця, які виділяють слиз крізь пори. У представників родів трахеломонас (Trachelomonas) і стробомонас (Strombomonas) окрім иелікули, навколо клітини утворюється твердий мінеральний будиночок, що складається з солей заліза і марганцю.
На передньому кінці клітини є мішковидне заглиблення - глотка - органоїд видільної системи. У розширену частину глотки — резервуар — виливається вміст скоротливих вакуолей.
Рух евгленових водоростей здійснюється за допомогою джгутика. Джгутик прикріплений своєю основою до базального тіла на дні резервуару. Через глотку він виходить назовні.
У більшості видів джгутик один; деякі евгленові забезпечені двома джгутиками рівної або нерівної довжини.
Рух більшості евгленових — це активне плавання. Працюючи джгутиком, клітина одночасно обертається навколо своєї осі і ніби угвинчується у воду. У деяких безбарвних представників евгленових, що живуть на дні серед мулу, поряд із плаванням (Регапета, Неїегопета) спостерігається і повзання по дну або іншому субстрату (Petalomonas). Клітина повзає, ніби обмацуючи дорогу попереду своїм джгутиком.
Зелене забарвлення хлоропласту обумовлене присутністю хлорофілів а і Ь. Це ті ж самі форми хлорофілу, які є і у вищих рослин. Крім хлорофілу присутні й інші пігменти: декілька каротинів і ксантофілів.
За формою хлоропласти евгленових досить різноманітні: дископодібні, пластинчасті, стрічкоподібні, зірчасті, лопатеві тощо. Число їх коливається від 1—2 до багатьох. Деякі види роду евглена (Еиріепа), виробляють велику кількість червоного пігменту (астак-сантину), що відіграє певну роль у регулюванні процесу засвоєння світла хлоропластами.
Основною запасною речовиною евгленових водоростей є полісахарид парамілон, який концентрується навколо піреноїда або утворюється безпосередньо в протоплазмі.
Вічко, або стигма, розташоване на передньому кінці клітини поблизу джгутикового апарату (рис. 6.16).
Ядро велике, нуклеоплазма містить одне або декілька ядерць.
Живлення у евгленових автотрофне, мезо-трофне, гетеротрофне, що обумовлює наявність значного числа безбарвних форм.
Фототрофний тип живлення характерний для більшої частини представників порядку евгленових: евглен (Euglena), трахеломонасів (Trachelomonas), стромбомонасів (Strombo-monas), лепоцинклісів (Lepocinclis) і інших зелених форм. Сапрофітний тип живлення переважає у безбарвних представників цього ж порядку: астазій (Astasia), дистигм (Distigma), меноїдіумів (Menoidium) і інших форм, що не мають хлоропластів.
Для родів перанема (Peranema), гетеронема (Heteronema), ентосифон (Entosiphon) і інших, що об'єднуються в порядок перанемо-вих, характерний голозойний (анімальний) тип живлення: заковтування оформлених частинок органічної речовини або інших дрібніших організмів.
Рис. 6. 16. Евглена зелена (Euglena viridis):
1 — джгутик; 2 — вічко (стигма); 3 — скоротлива вакуоля; 4 — хроматофори; 5 — ядро

Розмноження відбувається шляхом поздовжнього поділу особини на дві.
Евгленові - це переважно прісноводні водорості, досить широко розповсюджені у водоймах з повільною течією. Більшість з них розвивається в поверхневих або придонних шарах води. Деякі види повзають по дну. Як виняток серед евгленових водоростей евтреп-ція (Еиігерйа), яка віддає перевагу морським або солонуватим водоймам. Серед евгленових виявлені і паразитичні форми, що живуть в кишечниках олігохет, нематод, копепод, амфібій, на зябрах риб.
Господарське значення евгленових полягає у здатності деяких з них до міксотрофного живлення.
Ряд видів використовуються як біологічні індикатори забруднення водойм, у медичній практиці як модельні об'єкти для з'ясування дії різних лікувальних препаратів.
Багато видів евглен (Euglena) володіють здатністю до масового розвитку, викликаючи «цвітіння» континентальних вод (рис. 6.17).
Рис. 6. 17. Забарвлені евгленові водорості: 1 - Euglena sanguinea, заповнена гематохромом,
2 - Strobomonas ensifera, 3 - Euglena ehrenbergii, 4 - Trachelomonas bituricensis
У трахеломонасів (Trachelomonas) є своєрідні коричневі «будиночки», стінки яких просочені гідроксидом заліза. Трахеломонаси добре розвиваються у водоймах з гуміфікованою водою, багатою сполуками заліза, які здійснюють пригнічуючий вплив на більшість інших водоростей. Трахеломонаси часто зустрічаються у в малих лісових заболочених водоймах Полісся.
До порядку перанемові (Peranematales) відносяться безбарвні організми, без хлоропластів, які харчуються переважно голозойно. Мешкають у прісних, рідко в солонуватих водах, багатих на органічні речовини.
До порядку евгленоморфові (Euglenomorphales) відносяться паразити, що живуть в кишечниках червів, ракоподібних, амфібій, на зябрах риб. Клітини різноманітної форми, значна кількість живе усередині тіла господаря без джгутиків. Джгутики з'являються лише після вивільнення паразита з тіла господаря.
Порядок включає однойменну родину і понад 100 видів паразитних евгленових. Найвідоміші види родів Euglenamorpha, Hegneria, паразитні форми роду Astasia і ін. Ці водорості часто зустрічаються в поверхневих шарах мулу в різних малих водоймах.
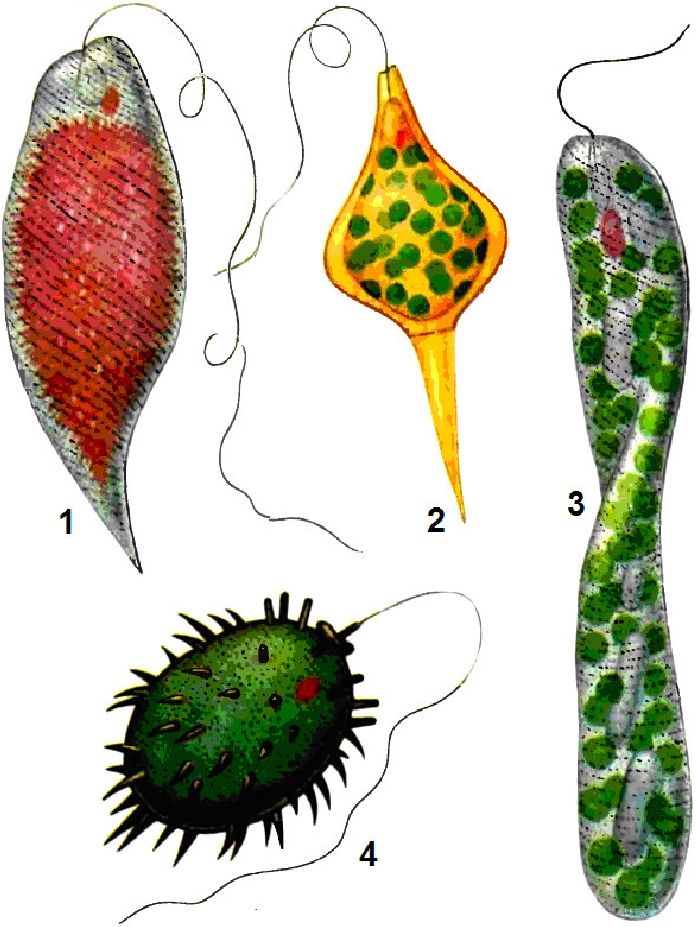
Динофітові водорості, або динофлагеляти, — це переважно одноклітинні водорості з монадним типом структури. Проте є і колоніальні і, зрідка, нитчасті форми. Клітини динофітових двобічно симетричні або асиметричні, переважно округлої, еліпсоїдної, яйцевидної, рідше грушовидної або іншої форми тіла. Деякі види мають форму тіла у вигляді півмісяця або піраміди. Клітини зазвичай забарвлені в темно-бурий, червоний, іноді в жовтий або жовто-зелений колір, є також безбарвні форми.
Розміри коливаються в межах від 6 до 1500 мкм. Серед прісноводних представників найбільших розмірів досягають види роду цераціум (Сегайит) — до 450 мкм. Відомо близько 300 видів динофітових, що мешкають у прісних і морських водах.
Клітинний покрив у найбільш примітивних форм представлений гладким тонким і ніжним перипластом. Переважна більшість динофітових вкриті целюлозною оболонкою, яка складається з пластинок (щитків), які часто утворюють панцир. Він складається з трьох основних частин: верхньої — епітеки, нижньої — гіпотеки і середньої — пояска, що сполучає обидві частини в суцільний панцир (рис. 6.18). Епітека і гіпотека складаються з пластинок і щитків, сполучених у певному порядку швами. Число і розміщення пластинок, а також наявність шипиків на поверхні належать до важливих систематичних ознак.
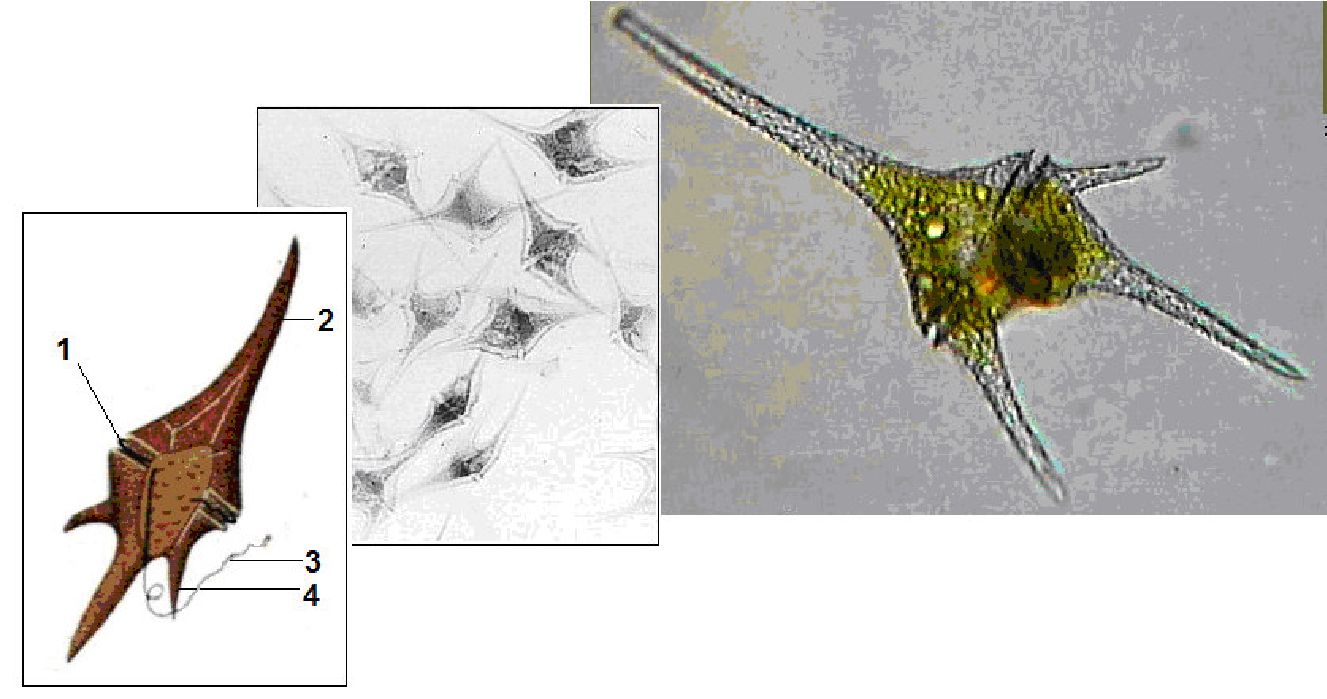
Рис. 6. 18. Цераціум (Сегайит): 1 - поясок, 2 - апікальний виріст, 3 - задній джгутик, 4 — антапікальний виріст
Монадні динофітові мають по два джгутика різної довжини і будови — поперечний і подовжній. Поперечний джгутик має вид хвилястої нитки, виходить з верхньої джгутикової пори і опоясує клітину вліво. Цей джгутик розташований в поперечній борозенці. Подовжній — рульовий — джгутик у вигляді тонкої нитки виходить з нижньої джгутикової пори, і у більшості видів розташований в подовжній борозенці панцира. Він тягнеться уздовж клітини і забезпечує поступальний рух.
Динофітовим властива своєрідна будова ядра, яка називається динокаріоном, в якій хромосоми кріпляться до ядерної мембрани. Хромосоми, на відміну від інших еукаріот, без гістонів і складаються з мікрофібрил, що містять чисту ДНК. Під час мітозу ядерна оболонка не зникає і веретено поділу не утворюється. Це ядро мезока-ріотного типу, яке вважається проміжним між нуклеоїдом прокаріотів і справжнім ядром еукаріотів.
Хлоропластів у клітині багато (рідше декілька або один). Хлоропласти містять хлорофіли а і с, Р- і у-каротини, та ряд ксантофілів. Піреноїди спостерігаються дуже рідко. Вічко (стигма) розташоване за межами хлоропласту, в основі джгутиків.
У прісноводних видів є розсіяні в протопласті вакуолі. Характерною особливістю динофітових є своєрідні вакуолі, які називаються пузулами, що відкриваються вузьким каналом у місці прикріплення джгутиків. Пузула нагадує мішок або трубку, яка глибоко заглиблюється всередину клітини і безпосередньо дотикається з вакуолею з клітинним соком. У деяких представників виявлені трихоцисти — паличкоподібні або булавоподібні безбарвні утворення, радіально розташовані на периферії протопласту.
Продуктами асиміляції динофітових є крохмаль і олія, рідше хризоламінарин, глікоген і ін.
Розмноження у динофітових водоростей здійснюється в основному поділом клітин в рухомому стані або зооспорами і апланоспо-рами. У деяких видів описаний статевий процес, що відбувається шляхом злиття дорослих клітин або джгутикових клітин і апланос-пор. За несприятливих умов у багатьох панцирних видів утворюються товстостінні цисти (Сегайит согпиічт).
Через нез’ясоване таксономічне положення в системі органічного світу, володіючи ознаками як рослин, так і тварин, динофітові привертають увагу ботаніків і зоологів. Цю таксономічну групу роз
глядають або у складі фітопланктону, або як компонент протозоопланктону. Одні і ті ж види (при збереженні їх видових і родових назв) входять до складу двох систем - ботанічної і зоологічної, підкоряючись двом різним номенклатурним кодексам.
Динофітові - це одна з домінуючих груп мікроводоростей у водних екосистемах. Це типові мешканці пелагіалі морів і океанів, хоча зустрічаються і в прісних континентальних водоймах.
Динофітові мають високу різноманітність у морських екосистемах і можуть викликати «червоні припливи», розвиваючись у масових кількостях. Багато видів токсичні і можуть викликати загибель тварин під час масового розвитку. Нейротоксини, які виробляються динофітовими, накопичуються організмами-фільтраторами (молюсками), і загрожують людям, які вживають їх у їжу. Деякі безбарвні форми (фістерія) теж можуть утворювати токсичні «цвітіння».
Є морські динофітові, здатні до біолюмінесценції, які при масовому розвитку викликають блакитне мерехтіння води вночі.
Деякі види є ендосимбіонтами морських тварин, а також відіграють важливу роль у біології коралових рифів.
У наших прісних стоячих водоймах широко розповсюджені представники класу динофіцієвих із родів перидиніум (Peridinium) і цераціум (Ceratium). Динофітові водорості складають істотну частину фітопланктону Чорного моря, літом їх значення істотно зростає. Останніми роками динофітові частіше стають причиною «цвітіння» води, які спостерігаються у північно-західному районі моря (Gymnodinium sanguineum, G. simplex, Prorocentrum cordatum і P. compressum) (рис. 6.19).
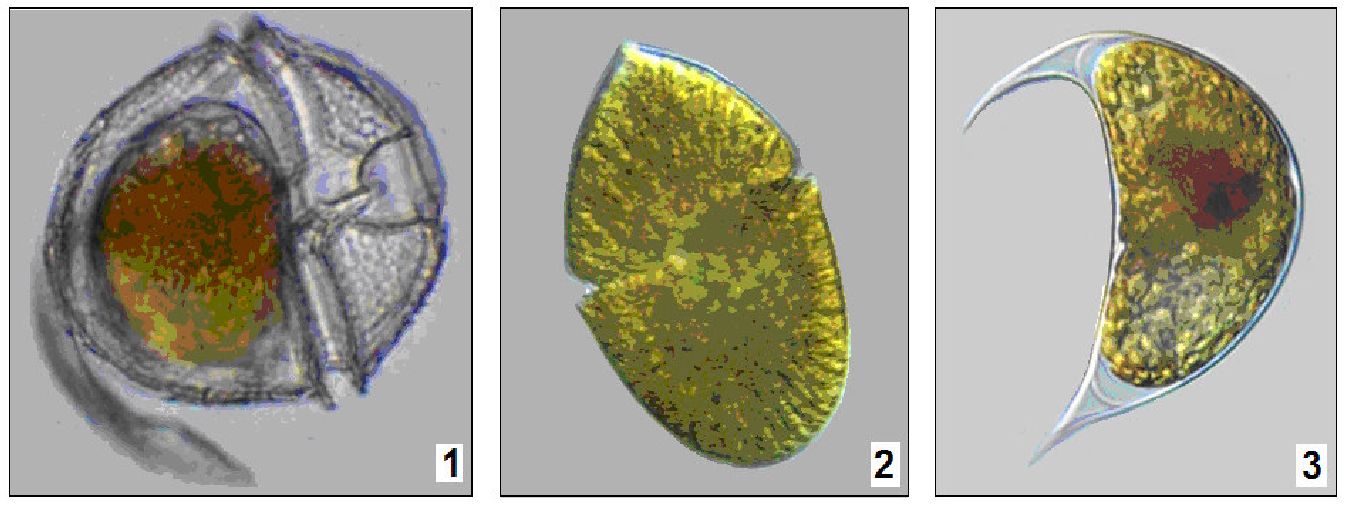
Рис. 6. 19. Динофітові водорості: 1 — Peridinium, 2 — Gymnodinium, 3 — Cystodinium
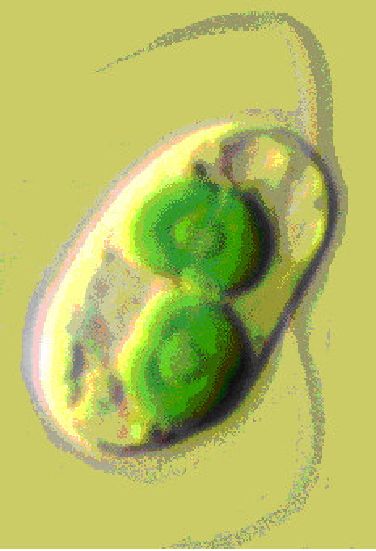
Ночесвітка, ноктилюка (Noctiluca) - це рід панцирних джгутикових з округлим тілом (діаметр 0,6-3 мм). Цитоплазма заповнена жировими включеннями, які при механічних чи хімічних подразненнях, окислюючись, починають світитися (рис. 6.20). Noctiluca miliaris - один з основних організмів, що викликають світіння моря. Ночесвітки утворюють скупчення в поверхневих шарах теплих, рідше бореальних вод, у Чорному і далекосхідних морях.
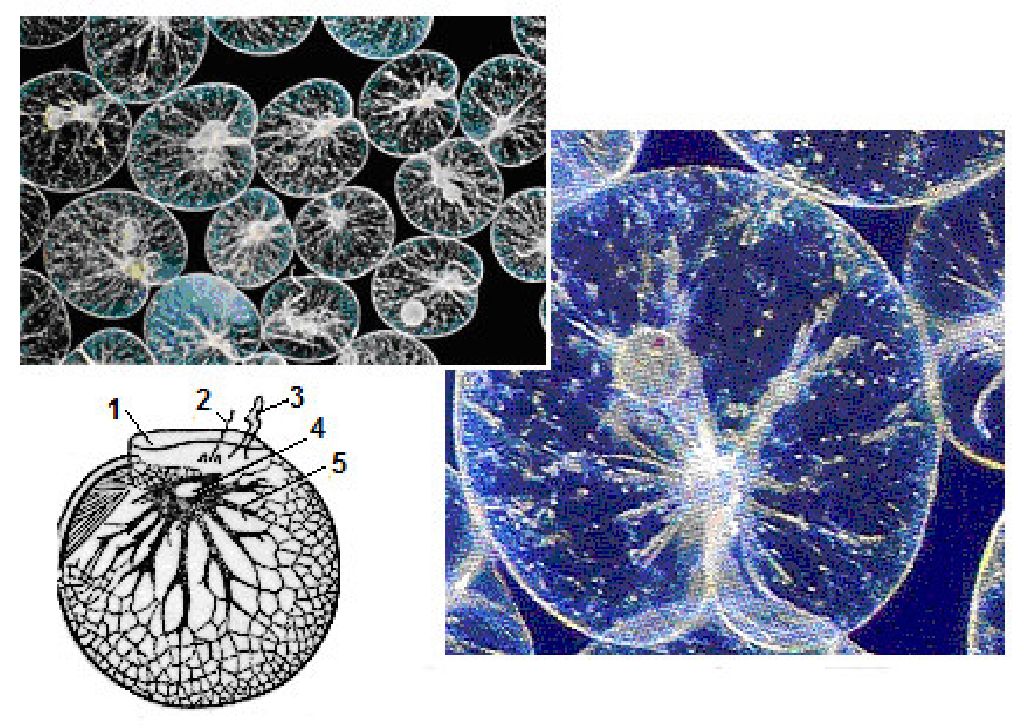
Рис. 6. 20. Ночесвітка (Іосйіиса): 1 — павколоротова западина; 2 — джгутик; 3 — щупальце (товстий джгутик); 4 — ядро; 5 — тяжі цитоплазми
6.4.4. Криптофітові водорості (СгурІорЬуІа)
Криптофітові водорості (криптомонади) є невеликою своєрідною групою прісноводних і солонуватоводних одноклітинних організмів, які переважно мають монадний тип структури. Талом у них завжди одноклітинний, і лише у деяких видів при розмноженні можуть виникати скупчення нерухомих клітин, сполучених слизом. Значно рідше зустрічаються пальмелоїдна і кокоїдна структури.
Відділ об'єднує близько 100 видів, що відносяться до 21 роду. Є ботанічні і зоологічні класифікації криптофітових.
Переважно клітини криптофітових водоростей мають опуклий спинний і увігнутий черевний бік. Розміри клітин від 6 до 50 мкм.
Передній кінець клітини з заглибленням, він криво зрізаний. Глотка мішковидна, по її краю і на поверхні знаходяться трихоцис-ти у вигляді паличок або зерняток, які при подразненні викидають тонкі нитки.
Клітини криптофітових водоростей мають покрив у вигляді щільного перипласту, часто подовжньо або спірально покресленого у зв'язку з наявністю в зовнішньому шарі цитоплазми своєрідних тілець, що розташовані правильними рядами і виділяють слиз.
Два джгутики майже однакової довжини, при русі або направлених вперед, або один — вперед, інший — назад (рис. 6.21).
Хлоропластів один-два, рідше більше; вони забарвлені в різні кольори від синьо-зеленого і оливково-зеленого до жовто-бурого, коричневого і темно-червоного. Іноді хлоропласти відсутні. Продукт асиміляції — крохмаль, у деяких представників олія і хризола-мінарин.
Живлення фотоавтотрофне або сапрофітне, рідко голозойне.
Стигма виявлена не у всіх представників, вона є частиною хлоропласту.
На передньому кінці клітини знаходиться одна або декілька скоротливих вакуолей.
Розмноження у криптофітових відбувається шляхом подовжнього поділу надвоє в рухомому або нерухомому стані.
У циклі розвитку деяких видів утворюються округлі цисти, вкриті товстим слизовими обгортками до 40 мкм у діаметрі.
У криптомонади (Cгyptomonas frigoris) утворюються сферичні цисти з гладенькою, безбарвною, безструктурною оболонкою до 10—15 мкм в діаметрі. Виникають вони в природних умовах на снігу і існують тривалий час. При настанні сприятливих умов вони проростають, при цьому вміст кожної цисти ділиться і утворюється 2 дочірніх клітини, які після розриву оболонки виходять з цисти назовні і вкриваються власними оболонками.
Криптофітові водорості — це типові представники планктону.
1 2
З
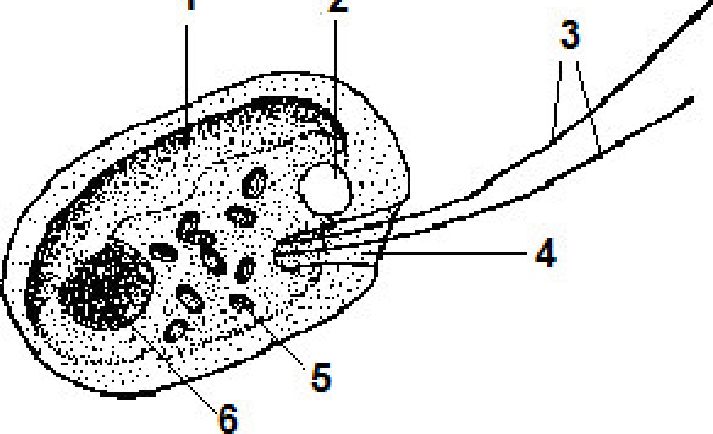
Рис. 6. 21. Будова клітини криптомонад: 1 — хлоропласт,
2 — скоротливі вакуолі,
3 — джгутики, 4 — глотка,
5 — піреноїди, 6 — ядро

Зрідка вони зустрічаються в мулі солоних озер (Cryptomonas salina, С. stigmatica) і в поверхневій плівці мулу прісноводних водойм, серед детриту. Є види, що живуть в болотах з низькими значеннями pH (3,5—4,1), наприклад, Cryptomonas суііпйгіса. Є ліофільні види, поширені у сірководневій зоні озер на глибині від 13 до 25 м (Сгуріаиіах іНіорНіїа, Сгуріотопаз рНазеоІиз).
3
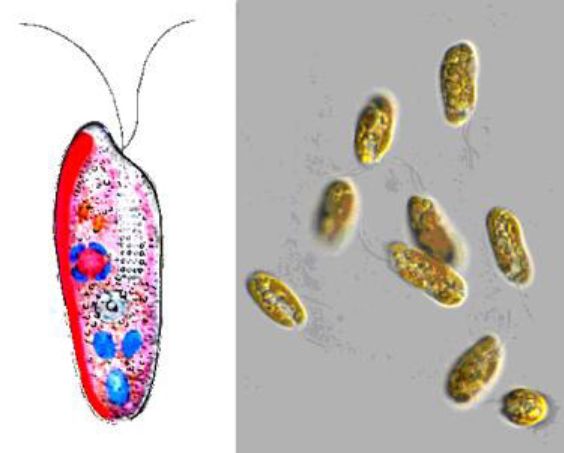


Рис. 6. 22. Криптофітові водорості: 1 - Cyanophora paradoxa, 2 - Chilomonas paramaecium, 3 - Rhodomonas, 4 - Chroomonas
Найцікавіші в екологічному і практичному відношенні види, що живуть у стічних водах, забруднених різними речовинами. Часто вони досягають тут масового розвитку і навіть викликають «цвітіння» води (Cryptomonas ovata, С. caudata, С. compressa і ін.).
6.4.5. Рафідофітові водорості (Raphidophyta)
Рафідофітові водорості є невеликою, чітко відособленою групою мікроскопічних одноклітинних джгутикових організмів, що налічує близько 20 видів. За рядом ознак вони різко відрізняються від інших груп водоростей, у першу чергу це стосується будови джгутиково
го, ядерного і вакуолярного апаратів, структури клітинного покриву, складу пігментів, продуктів асиміляції
Клітини мають різну форму. Довжина від 6-21 мкм у видів роду мономастикс (Monomastix) до 20-100 мкм у гоніостомума ^оп^іютит). Клітини світло-зелені або безбарвні. Передній кінець має заглиблення, з якого виходять два джгутики різної довжини. Один з них при русі направлений вперед, інший, рульовий, тягнеться ззаду.

Рис. 6. 23. Рафідофітові водорості: 1 - СЬайопеІІа шагіпа, 2 - СЬайопеІІа зр., 3 - Оопуозіошиш зр., 4 - УасиоІагіа
Клітини вкриті порівняно тонкою плазмалемою, переважно гладенькою і безбарвною, під якою знаходяться слизисті тільця або голковидні, радіально розташовані трихоцисти, що викидають при подразненні довгі слизисті нитки.
Ядро досить велике, кулясте або еліпсоїдне, з одним або декількома ядерцями, розташоване у внутрішньому шарі цитоплазми. На верхній частині ядра знаходиться напівкругле утворення — супра-нуклеарний аппарат.
Хлоропласти дрібні, переважно дисковидні або еліпсоїдні, переважно численні (рідше 1-2), світло-зелені. Продукт асиміляції — олія або крохмаль і лейкозин.
Верхня частина клітини містить вакуолярний апарат, що складається з однієї великої і кількох дрібних скоротливих вакуолей, які виливають свій вміст у глотку, а потім через вивідний канал назовні.
Розмножуються рафідофітові подовжнім поділом клітини в рухомому стані без втрати джгутиків (Gоniоstоmum seтeт). При поділі дочірні клітини певний час можуть знаходитися в слизистих скупченнях по 2-4, звільняючись після повного зникнення слизу.
Живлення у забарвлених рафідофітових — фотоавтотрофне, у безбарвних представників — сапрофітне і голозойне. Голозойні форми харчуються різними мікроорганізмами, іноді одноклітинними водоростями, навколо яких утворюють тимчасові травні вакуолі.
Рафідофітові водорості найчастіше зустрічаються в планктоні невеликих прісноводних стоячих водойм: ставках, озерах, старицях, в сфагнових болотах, проте, переважно у незначних кількостях, або одиничними екземплярами.
Інтерес до рафідофітових водоростей посилився в другій половині 20-го сторіччя у зв'язку з масовим розвитком в деяких озерах Скандинавії гоніостомума (Gonyostomum semen), що викликає інтенсивне «цвітіння води», ускладнює рекреаційне використання озер і може викликати токсичні ефекти. У країнах Скандинавії роботи з вивчення цього виду були включені в національні програми.
6.4.6. Золотисті водорості (Chrysophyta)
Золотисті, або хризофітові, водорості представлені одноклітинними, колоніальними або багатоклітинними організмами характерного золотисто-жовтого, буро-зеленого, рідше зеленувато-жовтого кольору. Є безбарвні форми. До відділу золотистих водоростей відносяться близько 800 видів.
Морфологічно дуже різноманітні та представлені різними типами структур: амебоїдною, джгутиковою, монадою, пальмелоїдною, кокоїдною, нитчастою і пластинчастою (рис. 6.24). Більшість видів мають монадний тип будови тіла. Монадні форми і стадії мають джгутики (переважно 1-2, рідше 4).
Розміри клітин золотистих водоростей коливаються в межах 2120 мкм.
Клітини в більшості випадків без клітинної оболонки, іноді вкриті лише плазмалемою, часто також дрібними лусочками з кремнезему, що можуть зростатися у панцир, рідше утворюють будиночки (Chrysococcus, Hyalobryon). Будиночки бувають кулясті або яйцевидні, рідше іншої форми. Стінки будиночків складаються з клітковини, просоченої солями кальцію і заліза; останні забарвлюють будиночки в коричневий колір (Eleutheropyxis fulva). У деяких видів будиночки оточені слизом (Chrysocrinus hydra) або зернистими утвореннями (Heliochrysis eradians). Стінки будиночків можуть бути просочені кремнієм.
Рис. 6. 24. Основні типи структури тіла золотистих водоростей: 1 - амебоїдна (Brehmiella chrysohydra), 2 - амебоїдна (Chrysarachnion insidians), 3 - монадна (Synochromonas pallida), 4 - кокоїдна (Chrysobotrys parvula), 5 - пальмелоїдна (Hydrurus foctidus), 6 - різнонитчаста (Phacothamnion borzianum), 7 - пластинчаста (Phacodermatium rivulare)
Будиночки прикріпляються до субстрату підошвою, товстою слизистою ніжкою або тонкою стеблинкою.
Переважна більшість золотистих водоростей мають джгутики, а деякі представники утворюють випинання у формі ризоподіїв, псе-вдоподіїв і аксоподіїв. Якщо джгутик один, як, наприклад у хрому-ліни, він має перисту будову, якщо два, то один з них зазвичай перистий, інший гладенький (Ochromonas, Synura). У деяких родів (Chrysochromulina) обидва джгутики гладенькі і рівної довжини.
Під час поділу клітин або перед утворенням цист джгутики можуть або втягуватися, або скидатися. Втягування характерне для представників, що не мають щільних покривів.
Ядро звичайно одне, розміри його незначні.
Хлоропластів у клітині 1-2, рідше більше. У хлоропластах знаходяться піреноїди. Стигма є частиною хлоропласту, розташована на його передньому кінці.
Вакуолярний апарат складається з пульсуючих (скоротливих) або непульсуючих вакуолей.
Продукти асиміляції золотистих водоростей — це олії і хризола-мінарин, у деяких волютин і глікоген. У деяких видів роду Synuгa у клітині накопичується гематохром.
Розмноження нестатеве (вегетативне або за допомогою спеціалізованих клітин — амебоїдів, зооспор і апланоспор) і статеве (рідко). Вегетативне розмноження у одноклітинних форм здійснюється шляхом поділу клітини надвоє. Багатоклітинні форми можуть розмножуватися шляхом фрагментації тіла на окремі ділянки, здатні до подальшого розвитку. До вегетативного розмноження відноситься також брунькування.
Нестатеве розмноження за допомогою амебоїдів відбувається, наприклад, у представників, що мають будиночки. Амебоїди дуже дрібні, з короткими тупими псевдоподіями, виповзають з отвору будиночка і утворюють потім свій власний.
Зооспори можуть виникати в звичайних вегетативних клітинах або в особливих органах — зооспорангіях.
Статевий процес буває рідко. Характерною особливістю циклу розвитку золотистих є утворення цист різноманітної форми з целюлозною оболонкою, що пізніше просочується оксидом кремнію. Утворюються цисти у різний час року, зазвичай літом і восени.
До порядку коколітові (Соссоїіійа^) відносяться одноклітинні монадні переважно морські форми. Клітини вкриті слизистим обгортками, на поверхні і усередині яких відкладаються своєрідні вапнякові тільця — коколіти. Слизиста оболонка з часом перетворюється на панцир. Клітини зазвичай з двома хлоропластами і двома джгутиками.
Золотисті водорості поширені по всій Земній кулі, у водоймах різних типів. Переважна більшість хризофітових мешкають у прісних водах, і лише незначна їх кількість зустрічаються в морі. Більшість живуть у чистій воді, деякі зустрічаються у водоймах, забруднених різними органічними речовинами. Типовим місцем поширення золотистих водоростей є сфагнові болота з кислою реакцією води.
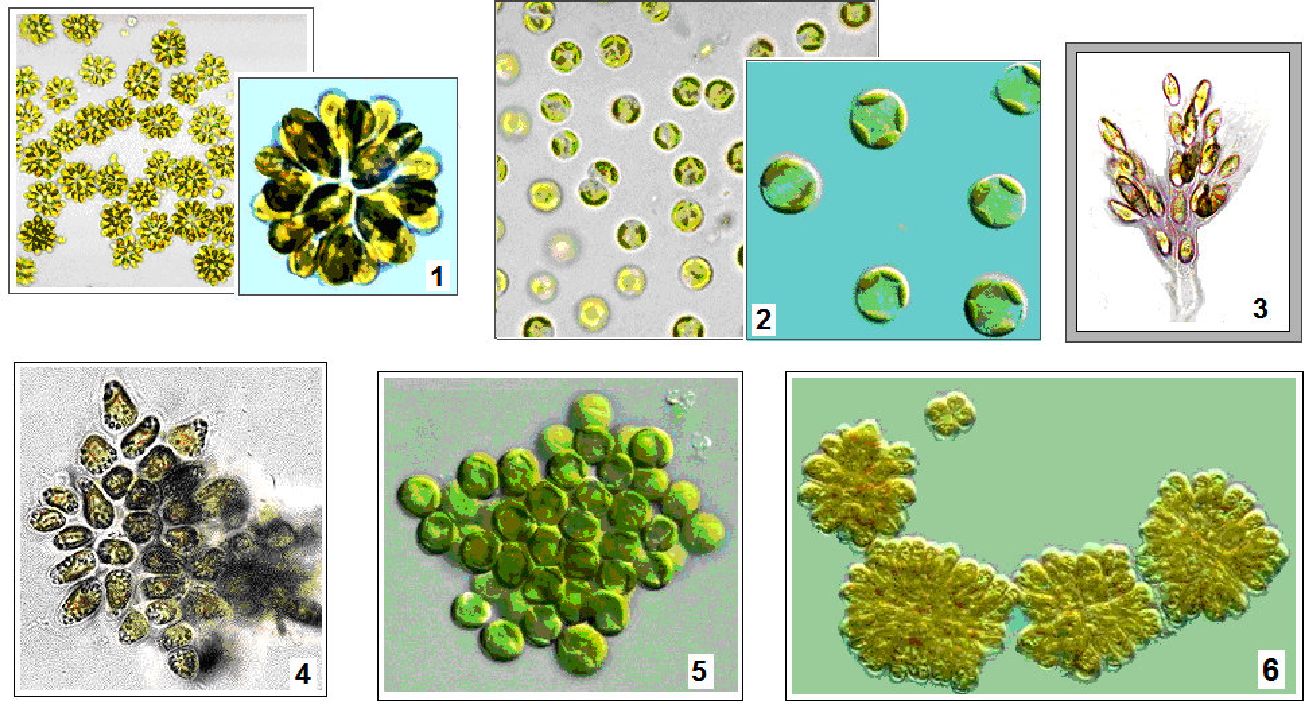
Рис. 6. 25. Золотисті водорості: 1 - Synura, 2 - Tetrasporopsis, 3 - Dmobryon, 4 - NaegeИeИa, 5 - Chrysocapsa, 6 - Phaeoplaca thaИosa
Деякі види з родів маломонас (Mallomonas), синура (Synura), ди-нобріон (Dinobryon), розвиваючись у масових кількостях, можуть викликати «цвітіння» води, додаючи їй запах риб'ячого жиру і погіршуючи її якість. Особливо важкі наслідки для рибних господарств має «цвітіння» ставів, що викликається масовим розвитком токсичного виду примнезіуму маленького (Prymnesium рагяит), що належить до класу иримнезіофіцієвих (Prymnesiophyceae). Цей вид широко розповсюджений у рівнинних і гірських озерах Західної Європи. Токсини, які виділяє цей вид, отруйні для риб і мало впливають на людей. Виявлений в Азовському морі.
Золотисті водорості беруть активну участь у накопиченні органічних речовин, мулу (сапропелю), який має велике практичне значення для народного господарства. Певне значення мають у ланцюзі живлення риб.
Викопні рештки золотистих водоростей відіграли певну роль в утворенні покладів вапняків і крейди, формуванні осадочних порід.
6.4.7. Діатомові водорості (Bacillariophyta, або Біаіютеае)
Діатомові водорості, діатомеї - це особлива група одноклітинних організмів, клітина яких зовні вкрита твердою оболонкою, що складається з прозорого панцира із кремнезему.
Діатомові водорості мікроскопічні (0,75-1500 мкм), однокліти
нні, одиночні або колоніальні форми; серед останніх зустрічаються види, що живуть в слизистих трубках і створюють бурі кущі заввишки до 20 см. Є колонії різного типу: ланцюжки, нитки, стрічки, зірочки, кущики або слизисті плівки.
Панцир складається з двох частин — епітеки і гіпотеки, що налягають одна на одну як кришка на коробку. Зовнішня частинка (епі-тека) більша, а внутрішня (гіпотека) менша. Кожна з них має стулку і обідковий поясок (рис. 6.26). Поверхня стулок плоска або випукла. Оболонки панцира пронизано дрібними отворами, що забезпечують обмін речовин між протопластом і навколишнім середовищем.
Форма панцира залежить від форми стулок і їх висоти. Завдяки симетричності будови панцира через нього можна провести подовжню, поперечну і центральну осі симетрії. Якщо через панцир можна провести всі три площини симетрії, то він повністю симетричний, дві — бісиметричний, одну — моносиметричний. Зустрічаються асиметричні панцирі, через які не можна провести жодної площини симетрії.
Панцир пронизують отвори — ореоли (рис. 6.27), затягнені зверху або зсередини перфорованою плівкою (велумом). Вони становлять 10-75% поверхні стулки. Частини отворів без стулок утворюють поля — центральне, осьове і бічне.
Потовщення, що виступають над зовнішньою або внутрішньою поверхнею стулки називаються ребрами, вони забезпечують міцність панцира. У багатьох діатомових водоростей на зовнішній поверхні панцира утворюються виступи, щетинки, шипики, які збільшують його поверхню і служать для з'єднання клітин в колонію.
За будовою панцира діатомові розділяють на два класи: пенатні (Реппаїорйуееае) з двобічносиметричним панциром і центричні (Сепїгорйуееае) з радіально-симетричним панцирем.
Велике біологічне значення має слиз, що виділяється діатомовими водоростями, який не лише сприяє утворенню колоній і прикріпленню водоростей до субстрату, але і захищає їх від висихання та дії інших несприятливих умов середовища.
Багато пенатних водоростей мають шов, який складається з одні-єї-двох щілин, що пронизують оболонку стулки від кінців до центра. Шов забезпечує взаємодію протопласту із зовнішнім середовищем і здатність до руху. Наявність шва — це прогресивна ознака, характерна для еволюційно молодих видів.

Рис. 6. 26. Пенатна водорість пінуля-рія (Pinnularia sp.): 1 — вузол, 2 — шов, 3 — епітека, 4 — гіпотека, — пояски
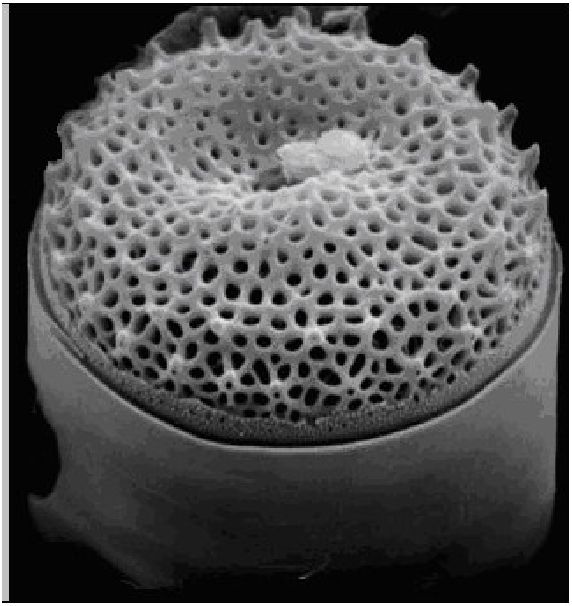
Рис. 6. 27. Ореоли на панцирі таласіозі-ри (Thalassiosira sp.)
Діатомові мають виключно кокоїдний тип структури тіла. Розмір клітин від 4 до 2000 мкм. У відділі, за даними різних авторів, налічується 12—25 тис. видів.
Клітини живуть поодинці або об'єднані в колонії різного типу. (рис. 6.28).
Ядро розміщене в центрі клітини в цитоплазматичному містку чи в пристінному шарі цитоплазми.
Хлоропласти діатомових водоростей різноманітні за формою, розмірами і положенням у клітині. У більшості діатомових хлоропласти дрібні, численні, у вигляді зерен або дисків. Лише у високо-організованих пенатних хлоропласти великі, один чи два в клітині.
Хлоропласти забарвлені в жовто-бурий колір з різними відтінками. Забарвлення зумовлено пігментами: хлорофілами aie, Р-каротином, s-каротином, рядом ксантинів. Пігментна система діатомових водоростей забезпечує фотосинтез на глибині до 50 м завдяки додатковим пігментам з групи фукоксантинів, які передають поглинену ними енергію хлорофілу а. Після загибелі клітини бурі пігменти розчиняються у воді і зелений хлорофіл стає яскраво помітним. У гетеротрофних безбарвних діатомових водоростей (Nitzschia) пігменти відсутні.
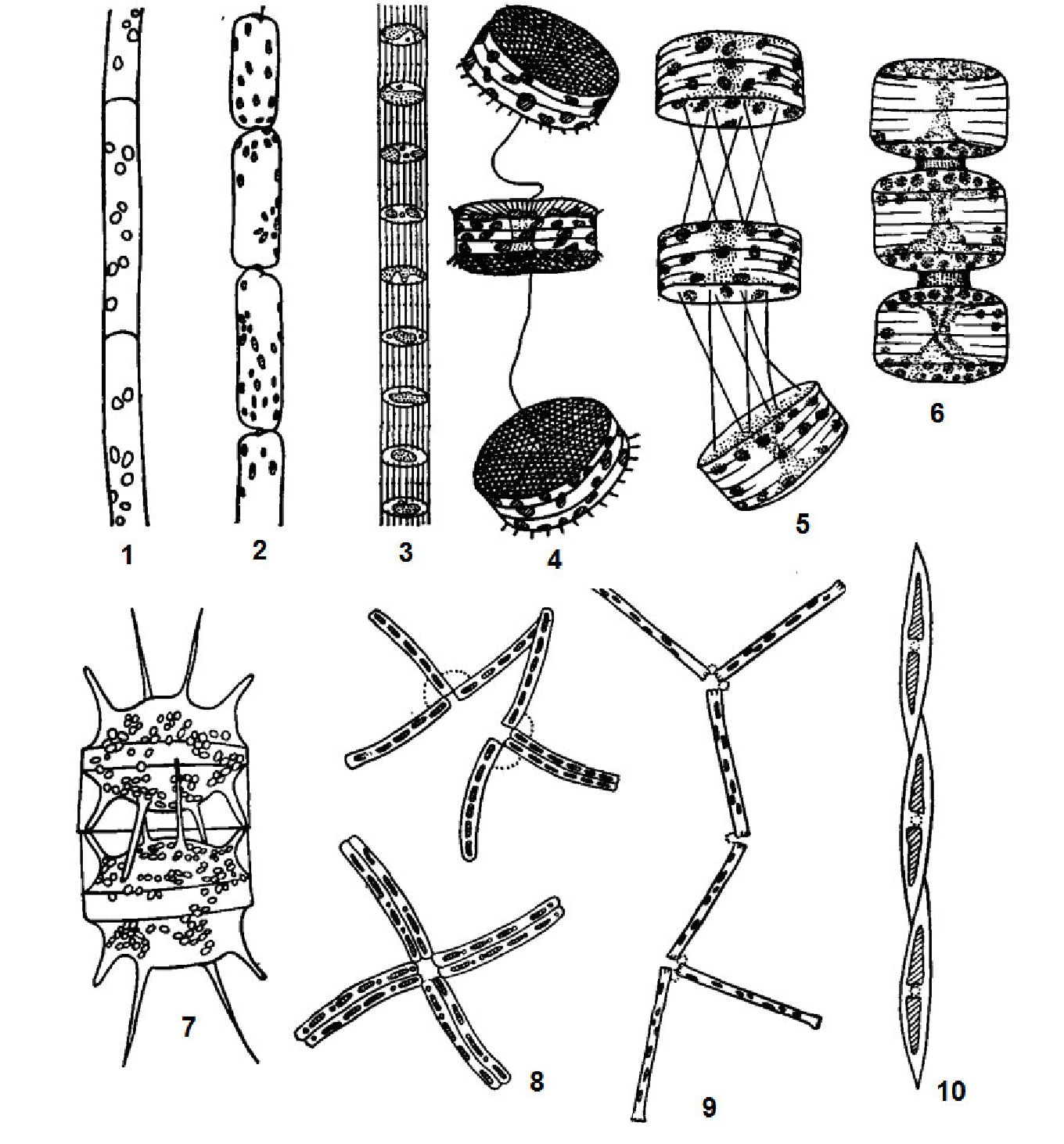
Рис. 6. 28. Типи колоній діатомових
1 - ниткоподібна колонія (йерівсуііпйгш ііапісш); 2-3 - ланцюжкові колонії (Rhizosolenia fragilissima, Sceletonema costatum); 4 - гнучкий ланцюжок (Thalassiosiгa excentrica); 5 - гнучкий ланцюжок (Thаlаssіosіrа роїусйогйа); 6 -прямий ланцюжок (Pоrоsіrа glaciatus); 7 - прямий ланцюжок, утворений за допомогою виростів (Віййиірйіа mоbіlіеnsіs); 8 - зигзагоподібний ланцюжок (Thalassionema nitzchioides); 9 - зигзагоподібний ланцюжок фіаіота еіопраіют); 10 - нитковидна колонія (І"^сйіа sеrіаtа)
У периферичній частині клітини знаходиться силікалема — складна система цистерн і трубочок, у яких виявлені елементи кремнієвого панцирю. Зовнішня мембрана цистерн силікалеми тісно
прилягає до плазмалеми. Функції силікалеми - формування панцирю і утворення нової плазмалеми.
Основний спосіб розмноження діатомових водоростей — вегетативний поділ клітини надвоє. Найінтенсивніше розмноження діатомових спостерігається навесні і восени. Перед поділом клітини у діатомей починають нагромаджувати в протопласті краплинки олії, завдяки чому він значно збільшується в об'ємі, епітека і гіпотека панцира розходяться. Протопласт ділиться на дві рівні частини. Ядро ділиться мітозом, утворюючи два дочірніх.
Після закінчення поділу кожна з дочірніх клітин одержує від материнської лише одну стулку панцира і добудовує обов'язково внутрішню, тобто гіпотеку. Після багаторазових поділів спостерігається поступове зменшення розмірів клітин водоростей. У деяких видів вони зменшуються в 3 рази у порівнянні з первинними. Відновлення первинних розмірів клітин відбувається під час проростання спор і в результаті статевого процесу.
Перед початком статевого процесу дві клітини зближуються, покриваються загальним слизистим чохлом і розсовують стулки. Після запліднення утворюється зигота, яка без стадії спокою починає рости, збільшується в розмірах і перетворюється на ауксоспору («спору, що росте»). Клітина, що утворюється з ауксоспори, називається ініціальною і більша за вихідну клітину.
У діатомових виявлено декілька типів статевого процессу: ізога-мний, анізогамний (гетерогамний), оогамний, автогамний.
Всі діатомові водорості диплоїдні організми, а гаплоїдна фаза ядра у них спостерігається тільки перед злиттям ядер у ауксоспорі.
У несприятливих умовах діатомові водорості переходять у стан спокою. Тоді протопласт пересувається до одного з кінців клітини і, унаслідок втрати клітинного соку, сильно стискається. Життєдіяльність цих клітин поновлюється при настанні сприятливих умов. Деякі прісноводні планктонні види в зимовий період занурюються на дно водойми, де перебувають в стані спокою або зниженої життєдіяльності до початку нового вегетаційного періоду.
Діатомові водорості широко розповсюджені і населяють різні біотопи: прісні і солоні, стоячі і проточні водойми, вологі скелі, ґрунт і навіть орні землі.
У водних екосистемах діатомові — цілорічні домінанти. У відкритій частині океану діатомові можуть розповсюджуватися до гли
бини 80-350 м, де утворюють глибоководний «тіньовий планктон». Це обумовлено здатністю деяких видів переходити на гетеротрофне живлення при великій кількості органічних речовин у воді. У планктоні прісних водойм переважають пенатні діатомові; глибина їх розповсюдження залежить від прозорості води.
У бентосних фітоценозах теж переважають пенатні (Іаяісиїа, Campylodiscus, Pinnularia, Gyrosіgmа). Переважно вони мешкають на глибині не більше 50 м.
Діатомові водорості — основна ланка трофічних ланцюгів водних екосистем. Вони відіграють основну роль у продуктивності водойм. У 100 г органічної речовини діатомових міститься 40% білків, 30% вуглеводів, 30% ліпідів. Діатомовими водоростями харчуються різні тварини, зоопланктонні організми — гіллястовусі і веслоногі ракоподібні, які складають найважливішу частину в живленні промислових риб.
Діатомові водорості здатні інтенсивно ділитися, даючи до чотирьох поколінь за добу, що нерідко викликає «цвітіння» води, появу неприємних запахів у питній воді ^іерйапо^сш йапі^сйіі, CydoteПa meneghiniana).
Щорічно поглинаючи зі Світового океану близько 3 млрд. т кремнію, діатомові водорості відіграють основну роль у його кругообігу. Викопні діатомові створили могутні відкладення, так звані діатоміти, або трепели, що мають промислове значення.
Сучасна класифікація діатомових водоростей заснована на морфології і структурі панцира. Виділяють два класи діатомових: центричні і пенатні. Структурні елементи на стулках панцира у центричних діатомових мають радіальне і тангентальне розміщення, у пенатних — двобічне або поперечне.
Клас центричні діатомеї (СеПторЬусеае)
Клітини поодинокі або сполучені в ниткоподібні або ланцюжкові колонії. Хроматофори зазвичай у вигляді дрібних численних зерен, рідше одна або декілька пластинок. Форма панцира різна: циліндрична, дисковидна, лінзовидна, куляста, еліпсовидна, діжковидна, рідше призматична, зі вставними обідками різної форми (рис. 6.29).
Стулки панцира мають радіальну будову і завжди без шва. Стулки округлі, еліптичні, трикутні і ін. По краю стулки розміщуються вирости або шпильки, за допомогою яких клітини з'єднуються в колонії, крім того є роги, щетинки та інші елементи структури.

Рис. 6. 29. Центричні діатомові: 1 - CydoteИa oceИata; 2 - CydoteUa (а - вигляд зверху, б - вигляд збоку); 3 - Arachnoidiscus єЬгєпЬє^іі; 4 - Melosira varians; 5 - Thalassюsira; 6 - Stephanodiscus
Центричні діатомові - це переважно морські планктонні організми. У прісних водоймах зустрічаються рідше.
У прісних і солонуватих водоймах часто зустрічаються види: та-ласіозіра (Thalassiosira Іасші^), циклотелла (Сусіоїеііа осеїіаіа), мелозіра (Mе1оsirа яагіащ), ризосоленія (Rhizosolenia longiseta).
Клас пенатні діатомеї (РеппяІорЬуееае)
Клітини поодинокі чи сполучені в колонії різного типу. Панцир симетричний по поздовжній осі, часто з вставними обідками. Стулки двобічно симетричні - лінійні, ланцетні, еліптичні (рис. 6.30).
Пенатні водорості — прісноводні і морські форми, що живуть в бентосі і лише поодинокі види планктонні.
Клас пенатних діатомей сформувався пізніше від центричних. Він поділяється на чотири порядки: безшовні (Агарйа^), одношов-ні (Мопогарйа^), двошовні фігарйа^) і каналошовні
(Aulonoraphales).
Діатомеї — порівняно молода група водоростей, їх викопні рештки знаходять починаючи від ранньокрейдяного періоду до сучасних відкладів. Перші викопні представники були морськими і належали до центричних діатомей. У крейдяному періоді з'явилися і пе-
рші види пенатного типу.

Рис. 6. 30. Пенатні діатомові: Tabellaria sp., 2 - Navicula radiosa, 3 - Pinnularia viridis, 4 - Gyrosigma sp., 5 - Rhopalodia gibba, 6 - Cocconeis sp. на кладофорі, 7 -Surirella sp., 8 - Gomphonema acuminatum, 9 - Cymbella sp., 10 - Cymatopleura solea, 11 - Eunotia serra, 12 - Amphora ovalis, 13 - Nitzschia sp., 14 - Asterionella formosa, 15 - Fragilaria sp., 16 - Meridion circula, 17 - Cymbella stuxbergi
Діатомеї є кормом для бактерій і найпростіших, якими в свою чергу живляться безхребетні тварини, риби та їх молодь.
До відділу відносяться близько 600 видів переважно мікроскопічних, рідше макроскопічних водоростей світло- або темно-жовто-зеленого, рідше зеленого кольору, іноді безбарвних. Розміри їх коливаються від 0,5—1,5 мкм (Chloridella glaciales) до десятків сантиметрів у довжину (нитки Tribonema, Vaucheria). Це одноклітинні, колоніальні, багатоклітинні і неклітинні організми, які активно рухаються або нерухомі, прикріплені і вільноживучі.
Спостерігаються майже всі типи структури вегетативного тіла: кокоїдний, монадний, гемімонадний, сифональний, сифонокладаль-ний, нитчастий, різнонитчастий, псевдопаренхіматозний, паренхіматозний. Більшість мають кокоїдний тип структури (рис. 6.31).

Рис. 6. 31. Жовтозелені водорості: 1 - 3 - Воїгубіиш granulatum (1 - обростання на грунті при малому збільшення, 2 - окрема особина при великому збільшенні, 3 -утворення ризоцист); 4 - МізсИососсш сопїегуісоїа; 5 - СШогоШесіиш сгаззіарех, клітина і вихід із неї зооспор; 6 - Регопіеііа сигуірез
У видів із амебоїдною, монадною і частково гемімонадною структурою вегетативного тіла клітини вкриті лише плазмалемою; вони легко змінюють свою форму, утворюючи псевдоподії.
У переважної більшості жовтозелених водоростей клітини мають міцну клітинну оболонку, цілісну або двостулкову. Клітинна оболонка пектинова, іноді з домішкою целюлози або геміцелюлози (ТгіЬопета), у вошерії (Уаисйегіа) — целюлозна. На поверхні оболонки може відкладатися вапно, часто вона насичена сполуками заліза, що забарвлюють її в жовтуватий, червонуватий, іржаво-червоний, бурий або коричневий тони. Ще частіше в клітинних оболонках жовтозелених водоростей виявляється кремнезем, що додає їй особливих твердості, крихкості і блиску.
Іноді клітини повністю вкриті слизом, що об'єднує їх в колонії (ОІоеоЬоїгуйасеае).
Хлоропласти різноманітні за формою і будовою. Число їх коливається від одного до декількох сотень в одній. Комплекс фотосинтетичних пігментів включає хлорофіли а і с, хлорофіл Ь відсутній. Є кілька каротиноїдів і ксантофілів. Піреноїди спостерігаються рідко. Запасними поживними речовинами є краплини олії, хризоламінарин та білкові кристали. У рухомих водоростей є стигма червоного, коричневого або оранжево-червоного кольору.
Монадні форми і стадії (зооспори, сперматозоїди) мають по два джгутики різної будови і довжини.
Прісноводні види, як правило, мають скоротливі вакуолі; лише у морських і солонуватоводних видів вони відсутні.
Вегетативне розмноження здійснюється поділом клітин або розпадом колоній і багатоклітинних форм на окремі частини. Найшир-ше у жовтозелених водоростей представлене нестатеве розмноження за допомогою спеціалізованих клітин: амебоїдів, зооспор і автоспор. Статевий процес (оогамного типу) з достовірністю відомий лише у видів роду вошерія (Уаисйегіа). Статеві органи — антеридії і оогонії — утворюються або на одній (однодомні), або на різних (дводомні) рослинах (рис. 6.32). Дводжгутикові антерозоїди, що утворюються в антеридії, проникаючи в оогоній, запліднюють яйцеклітину. Зигота, або ооспора, покривається товстою багатошаровою оболонкою і після періоду спокою проростає в нову рослину.
Жовтозелені поширені у воді, ґрунті і в повітрі на всіх континентах земної кулі.
Рис. 6. 32. Жовтозелені водорості: а - Botrydiopsis агйіга, б - Уаисйегіа (1 - антеридій, 2 - оогонії), 3 - ТгіЬопеша
Більшість видів жовтозелених — це прісноводні організми. Вони зустрічаються у різних водоймах, проте віддають перевагу чистим водам стоячих водойм з кислою реакцією середовища (сфагнові і торфові болота, канави, ями і калюжі на торф'яниках).
Більшість видів мешкають в товщі води або входять до складу перифітону. Бентосних і нейстонних форм небагато. Зрідка жовто-зелені водорості можуть викликати «цвітіння» води евтрофних водойм (Воітуйіорзіз агйіга).
Жовтозелені багато представлені в грунті, беруть участь в створенні грунтової родючості, при масовому розвитку можуть навіть викликати «цвітіння» поверхні грунту.
Жовтозелені водорості беруть участь в утворенні первинної продукції водойм, відіграють значну роль у живленні гідробіонтів, самоочищенні забруднених вод.
Найпоширеніший порядок ботридієвих (Воїгуйіаіез) з двома родинами Воітуйіасеае (1 рід) і Уаисйегіасеае (1 рід). Представники порядку широко розповсюджені по всій планеті в морях, солонуватих і прісних водах, а також на вологому грунті. Це неклітинні водорості сифональної структури. Макроскопічні таломи подібні до бульбашки (Воітуйіит) або нитки (Уаисйегіа), прикріплені або не-прикріплені.
Червоні водорості, або багрянки, - це багато-, одноклітинні і колоніальні організми від рожевого до темно-червоного, майже чорного кольору. Є близько 4 тис. видів, що відносяться до понад 600 родів. Поширені переважно у теплих морях і океанах. У прісних і солонуватих континентальних водоймах, а також в аерофітних умовах мешкає не більше 50 видів.
Морські багрянки - це переважно багатоклітинні великі рослини, зі складною будовою, які досягають у довжину від кількох сантиметрів до метра. Тіло має складну будову: розчленоване, нагадує розгалужені кущики або стрічковидні пластинки.
Серед морських багрянок є багато мікроскопічних форм, але лише найпримітивніші з них мають одноклітинні або колоніальні та-ломи. У високоорганізованих представників слань має складну тканинну будову. Периферійні шари слані складаються з дрібних корових клітин, багатих на хлоропласти. Внутрішні клітини більші за розміром, не мають хлоропластів і виконують різні функції.
Багатоклітинні таломи багрянок розгалужуються моноподіально, симподіально, дихотомічно. До субстрату водорості прикріплюються ризоїдами на нижньому боці талому, а також виростами-присосками або дископодібною підошвою.
Клітини червоних водоростей різноманітні за формою, з цілісною двошаровою оболонкою. Внутрішній шар оболонки складається з целюлози, зовнішній — з пектинових речовин. Серед пектинових речовин є колоїдні речовини — агар, карагінан, агароїди. Вони здатні розчинятися в киплячій воді з утворенням колоїдних розчинів. До групи пектинових речовин відносяться також колоїдні речовини, які містяться в клітинних оболонках і міжклітинниках багатьох морських багрянок (Рйуііорйога).
У морських високоорганізованих форм клітини багатоядерні, а у примітивних форм і репродуктивних клітин (спермаціїв, карпо- і тетраспор) одноядерні. Ядро маленьке, з ядерцем.
У цитоплазмі є один-два хлоропласти, у найпримітивніших форм з піреноїдом. Форма хлоропластів різна і не завжди постійна; вона може бути різною навіть в різних клітинах одного організму і змінюється з віком організму.
Червоні водорості мають складний набір пігментів. Поряд з хлорофілами а і б, а- і Р-каротинами і кількома ксантофілами, хлоро-
пласти червоних водоростей містять специфічні для даної групи рослин додаткові пігменти: червоний фікоеритрин і синій фікоціа-нін. Ці пігменти виявлені також у синьозелених і криптофітових водоростей. Різне кількісне співвідношення фікоеритрину і фікоціа-ніну визначає колір червоних водоростей. У прісноводних водоростей фікоеритрину мало або він відсутній, тому вони забарвлені переважно в сірий, темно-зелений, синьо-зелений або зелений колір.
Запасним полісахаридом є багрянковий крохмаль, який від йоду не синіє, а спочатку стає жовто-бурим, а потім червоним.
Більшість червоних водоростей - дводомні рослини, які мають статеві органи складної будови. Спорофіт і гаметофіт за будовою подібні. Розмножуються нестатевим і статевим шляхом.
Вегетативне розмноження одноклітинних і колоніальних форм відбувається поділом клітини. У багатоклітинних водоростей утворюються додаткові пагони, що перетворюються у нові рослини. Нестатеве розмноження у низькоорганізованих водоростей здійснюється моноспорами, тобто спорами, які утворюються по одній у клітині. Для більш високоорганізованих характерне нестатеве розмноження тетраспорами, які утворюються по чотири в клітині, що називається тетраспорангієм. І моноспори, і тетраспори без оболонки, без джгутиків. Потрапляючи у водне середовище, вони вкриваються оболонкою і проростають.
Моноспори і тетраспори проростають, як правило, в гаметофіти, на яких утворюються органи статевого розмноження, які мають складну будову.
Статевий процес оогамний. Лише одноклітинні і колоніальні багрянки статево не розмножуються. Гамети не мають джгутиків. Чоловічі гамети - спермації - утворюються в клітинах, які функціонують як чоловічі органи розмноження і називаються сперматангі-ями, або антеридіями. При заплідненні спермації викидаються назовні і пасивно переносяться течією води, а жіночі гамети - оогонії - залишаються на гаметофіті у жіночому статевому органі, який називається карпогоном і утворюється на кінцях бічних гілочок. Карпогон - це особлива клітина без хлоропласта. У бангієвих водоростей карпогон являє собою мало змінену вегетативну клітину, яка відрізняється від решти більшими розмірами і інтенсивнішим забарвленням. У флоридових нижня частина карпогону розширена, містить ядро, а верхня частина вузька, утворює трубчастий виріст —трихогіну, яка служить для вловлювання спермаціїв.
При заплідненні спермації прикріпляються до вершини трихогі-ни, оболонки їх в місці зіткнення розчиняються, і ядро спермація перетікає в порожнину трихогіни. Пересуваючись по ній, воно потрапляє в черевну частину карпогону і там зливається з його ядром. Після запліднення карпогон відділяється від трихогіни поперечною перегородкою, трихогіна відмирає.
Зигота без періоду спокою починає ділитися і утворює особливі спори — карпоспори, із яких розвиваються спорофіти (рис. 6.33).
У бангієвих після запліднення карпогон ділиться на 4-32 карпоспори, які у вигляді голих клітин виходять у воду. У флоридових у розвитку зиготи крім карпогону беруть участь інші спеціальні клітини.
Рис. 6. 33. Порфіра (РогрЬуга ер.) із пластинчастою структурою талому (1 -
загальний вигляд; 2 - вигляд зверху, помітні карпоспори; 3 - 5 поперечний зріз пластинки: 3 - з карпогонами, 4 - з сперматангіями, 5 - з карпоспорами)
З диплоїдних карпоспор утворюється тетраспорофіт, який розмножується лише нестатево - тетраспорами. Тетраспори утворюються редукційним поділом. Вони гаплоїдні і при проростанні утворюють статеву генерацію — гаметофіт.
Таким чином, у більшості червоних водоростей розвиток проходить такі форми: гаметофіт, карпоспорофіт, тетраспорофіт. У цьому циклі є вільноживучі форми розвитку — тетраспорофіти і гаметофіти. Редукційний поділ відбувається при утворенні тетраспор, у зв'язку з чим тетраспори і гаметофіти, що утворюються з них, є гаплої-
дною фазою, а зигота, карпоспори і тетраспорофіт — диплоїдною фазою.
Червоні водорості широко розповсюджені у всіх морях, але у напрямку високих широт загальна кількість видів багрянок поступово зменшується і різко падає в Арктичному басейні. Близько 2% видів багрянок зустрічається у прісній воді. Таломи їх прикріплюються до підводних субстратів — каменів, скель, водних рослин. Завдяки наявності червоних пігментів, які здатні засвоювати навіть незначну кількість світла у спектральному діапазоні, який недоступний для інших водоростей, багрянки можуть рости на глибині 100-200 м.
Серед червоних водоростей є епіфіти, ендофіти і паразити. Ен-дофітизм буває частковим або повним. У другому випадку талом ендофіта, який живиться автотрофно за рахунок фотосинтезу, повністю занурений в тканини водорості-господаря, а на поверхню виносяться тільки нитки з органами розмноження. Багато багрянок ведуть паразитичний або напівпаразитичний спосіб життя. їх таломи значно редуковані і виступають над поверхнею водорості-господаря у вигляді кульок або лопатеподібних утворень.
Господарське значення морських багрянок визначається присутністю в оболонках деяких водоростей великої кількості фікоколої-дів — слизистих речовин з групи полісахаридів. Найбільш відомим промисловим продуктом є агар, що одержується з видів родів Gelidium, Gracilaria, Ahnfeltia і інших агарофітів. Агар широко застосовують в медицині, мікробіології, харчовій, целюлозно-паперовій і текстильній промисловості. З інших фікоколоїдів істотне значення мають карагенін і агароїд. Карагенін одержують в основному з видів роду хондрус (Chondrus), агароїд — з чорноморської філофори (Phyllophora nervosa). Морські червоні водорості разом з іншими водоростями використовують також для виготовлення водоростевої муки, що йде на корм домашнім тваринам і на добриво. Ряд морських багрянок їстивні. Перше місце серед їстівних багрянок займають види родів Rhodymenia і Porphyra. У Японії розвинене промислове культивування порфіри в природних умовах. Аквакультура деяких червоних водоростей широко практикується в США, Канаді, Франції, Норвегії, Данії, Великобританії і інших країнах. З багатьох червоних водоростей отримують йод.
Відділ червоних водоростей нараховує близько 4000 видів і 600 родів. Червоні водорості діляться на два класи: бангієві і флоридові.
Клас бангієві (ВапріорЬусеае) включає одноклітинні, колоніальні, а також багатоклітинні водорості трихальної, гетеротрихальної і пластинчасто-паренхіматозної структури (рис. 6.34). Вегетативне розмноження поділом клітини на дві або більше дочірніх клітин. Нестатеве розмноження відоме тільки у високоорганізованих представників. Гамети формуються в результаті прямого перетворення вегетативних клітин, жіночі по одній, чоловічі — багато в кожній клітині. Зигота ділиться і вся перетворюється на карпоспори. Моно-спори і карпоспори голі, здатні до амебоїдного руху.
Більшість представників класу, особливо серед примітивних, — прісноводні і наземні мешканці. Морські форми поширені в прибережній зоні літоралі всіх морів. Є близько 20 родів і 70 видів.
Рис. 6.34. Бангієві водорості: 1 - Porphyridшm; 2 - Chroothece; 4 - однорядні і багаторяд-ні нитчасті колонії Goшotrichum
У країнах Східної Азії види порфіри (Porphyra) вживають у їжу.
До порядку компсопогонових (Compsopogonales) відноситься рід компсопогон (Compsopogon), представники якого можуть утворювати обростання - «чорну бороду» на листках акваріумних рослин. У компсопогону талом кущикоподібний, він прикріплюється до субстрату підошвою, а також ризоїдними виростами (рис. 6.35).
Клас флоридові (Florideophyceae) об’єднує багатоклітинні водорості найрізноманітнішої форми, переважно складної анатомічної будови. Слань їх представлена системою розгалужених ниток. Ріст апікальний за рахунок верхівкової клітини. У деяких форм талом рясно просочений вапном.
Клітини одноядерні, рідше багатоядерні, з одним або декількома хлоропластами, найчастіше без піреноїдів.
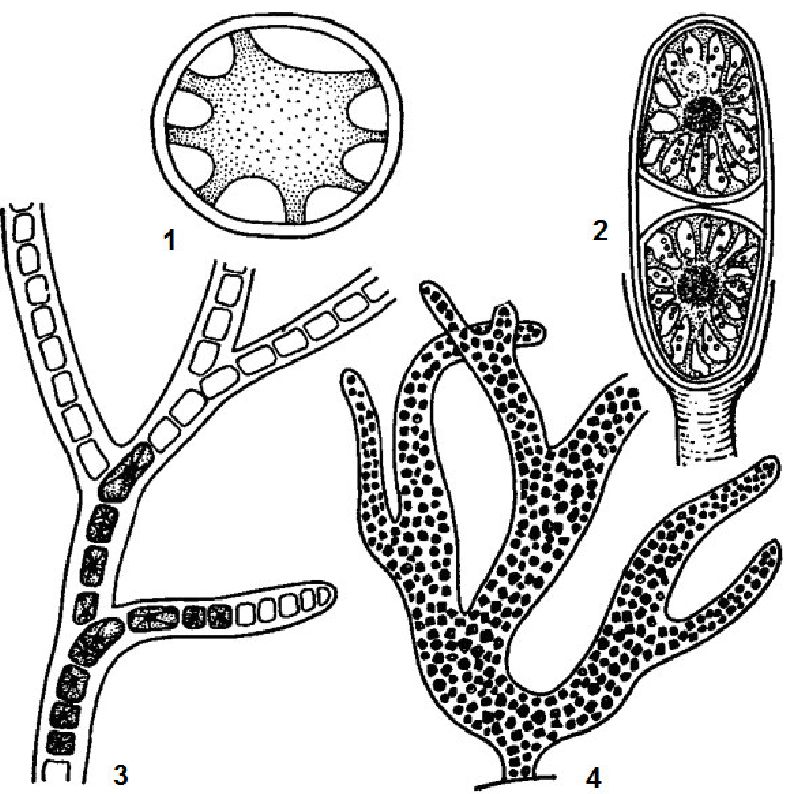
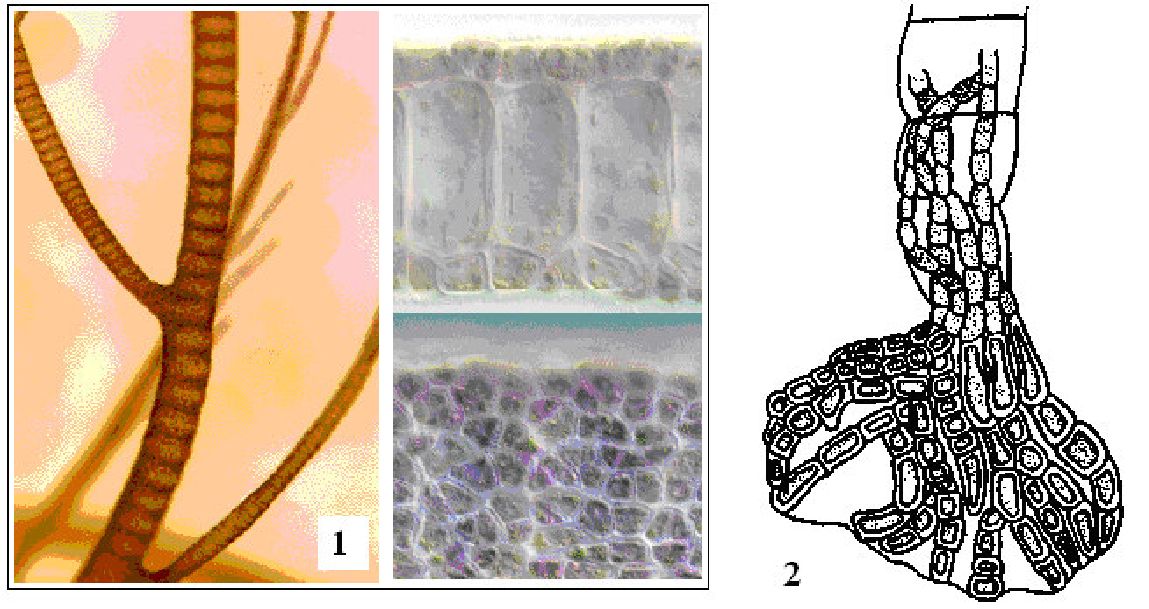
Рис. 6. 35. Компсопогон (Сошръоророи сптуЬвыъ). 1 - фрагмент талому (справа - мікроскопічна будова тканин); 2 - базальна частина талому з ризоїдами, які утворюють підошву
Переважна більшість флоридових — типові морські мешканці, широко розповсюджені у всьому Світовому океані.
До порядку гігартинові належать водорості різноманітних розмірів і форми, талом нитчастий, плоский, пластинчастий або шкірковидний, простий, розгалужений або розітнутий на лопаті, складної анатомічної будови (рис. 6.36).
У циклі розвитку відбувається чергування гаметофіта і спорофіта, схожих за зовнішньою і внутрішньою будовою.
Гігартинові — надзвичайно різноманітна група морських водоростей. Найвідомішіродини: Рйуііорйо-
гасеае, Gigartinaceae,
Огаеііагіасеае, Ийойо-рйуііійасеае і ін.

Рис. 6.36. Гігартинові водорості: 1 - філофора (Phyllophora crispa), 2 - гігартина (Gigartina sp.), 3 -хондрус кучерявий (Chondrus crispus), 4 - грацілярія (Gracilaria sp.)
Порядок родименієві ^койутепіаЄ). Талом від циліндричного до пластинчастого, простий або розгалужений, складної анатомічної будови, багатоосьової типу (рис. 6.37). У циклі розвитку відбувається чергування гаметофіта і спорофіта, схожих за зовнішньою і внутрішньою будовою. Поширені по всіх морях від тропіків до Північного Льодовитого океану; найбагатше представлені в теплих морях. Порядок представлений двома родинами: Сйатріасеае і КНосІутепіасеае.
До відділу бурих водоростей відносяться переважно морські макроскопічні водорості, загальною зовнішньою ознакою яких служить жовтувато-буре забарвлення. Бурі водорості — багатоклітинні рослини, великі, складно розчленовані, прикріплені до субстрату. Відомо близько 1500 видів бурих водоростей, що відносяться до 240 родів. У прісних, переважно холодних проточних водах помірних широт виявлено лише п’ять видів бурих водоростей.
Розміри таломів від кількох десятків мікрометрів до 30-50 м (у деяких видів родів Laminaria, Macrocystis, Sargassum). Слань має різну форму (нитчаста, куляста, мішкоподібна, пластинчаста, кущоподібна і ін.). Слані деяких представників мають повітряні пухирі пневматофори, які утримують гілочки у вертикальному положенні. Усі бурі водорості прикріплюються до грунту чи інших водних рослин за допомогою ризоїдів, базальних дисків.
У високоорганізованих бурих водоростей (Laminaria, Fucus, Sargassum) таломи диференційовані і нагадують квіткові рослини. У них є стебло-, листко- і коренеподібна частини, деякі представники мають повітряні міхурі, що утримують гілки у вертикальному положенні. Слань має примітивні тканини: асиміляційну, запасаючу, механічну і провідну.
Галуження бурих водоростей моноподіальне. Бічні гілочки чер
*ис. 6.37. Родименієві во-іорості: 1 - Rhodymenia iacifica, 2 - R. palmata
6.4.10. Бурі водорості (Phaeophyta)
гові розсіяні або супротивні. При швидкому доростанні їх до розмірів головної нитки виникає дихотомоподібне галуження.
Клітини бурих водоростей одноядерні. Оболонка їх складається з внутрішнього целюлозного шару і зовнішнього пектинового. Целюлоза бурих водоростей за своїми властивостями відрізняється від целюлози квіткових рослин, і тому її іноді називають альгульозою.
Сусідні клітини сполучаються за допомогою пор і плазмодесм.
Хлоропластів декілька, вони маленькі, дископодібні, стрічкоподібні, пластинчасті з дуже дрібними піреноїдами. До їх складу входять пігменти: хлорофіли а і с, каротини і ксантофіли, особливо фу-коксантин інтенсивно бурого кольору. Різні співвідношення цих пігментів обумовлюють забарвлення бурих водоростей від оливково-жовтуватого до темно-бурого, майже чорного кольору.
Клітини бурих водоростей мають одну велику або декілька дрібних вакуолей. Крім того, зустрічаються фізоди — це дуже дрібні вакуолі (до 4 мкм в діаметрі) із таніноподібною речовиною.
Запасною поживною речовиною є полісахарид ламінарин, шестиатомний спирт маніт і в невеликих кількостях олія.
Розмножуються бурі водорості вегетативно, нестатевим і статевим способами.
Вегетативне розмноження відбувається при випадковому відокремленні гілок від слані. Спеціальні бруньки вегетативного розмноження є лише у видів роду сфацелярія ^рйасеїагіа).
Нестатеве розмноження здійснюється зооспорами, нерухливими тетраспорами і моноспорами. Зооспори і гамети утворюються у одно- і багатогніздих вмістищах (рис. 6.38).
Статевий процес ізо-, гетеро- і оогамний. Гамети зазвичай утворюються в багатогніздних гаметангіях, по одній в кожній камері.
Більшість бурих водоростей мають самостійні форми розвитку — спорофіт і гаметофіт або гаметоспорофіт (рис. 6.39). Гаметофіт і спорофіт можуть мати як однакову будову і розміри, так і різну, тобто у бурих водоростей існує ізоморфна і гетероморфна зміна форм розвитку. Найчастіше більші розміри і складнішу будову має спорофіт.
До бурих водоростей, що не мають зміни форм розвитку, а лише зміну ядерних фаз, відносяться представники родин фукусових Цисасеае), цнстозейровнх (Cystоsеirасеае) і саргасових
(Ба^а88аееае).
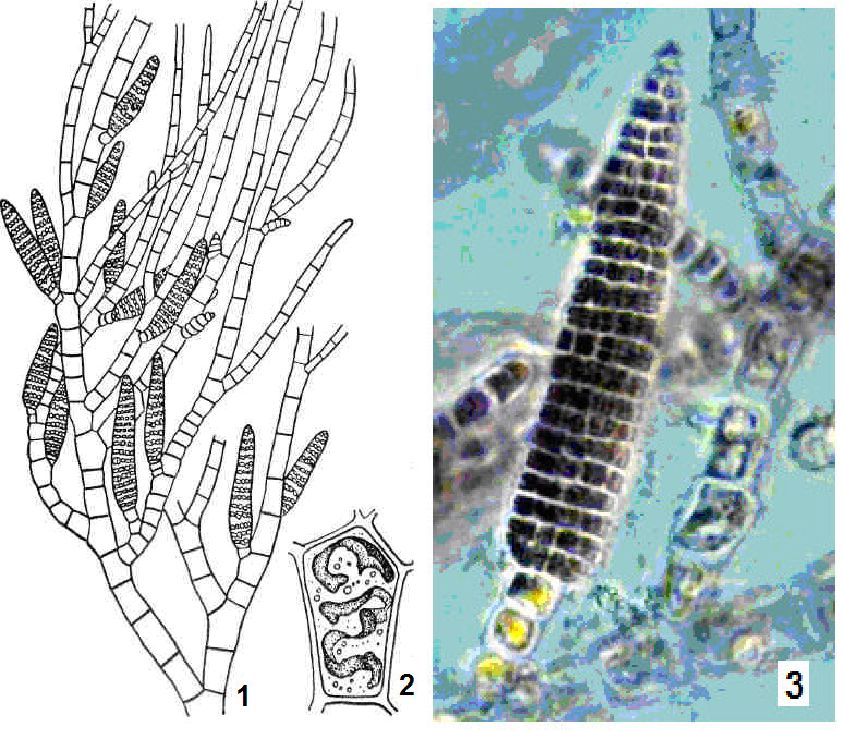
Рис. 6.38. ЕСосагрш 8ІІісиІ08и8: 1 - фрагмент талому з багато гніздними спорангіями; 2 — клітина із стрічкоподібними хлоропластами; 3 - спорангій (мікрофотографія)
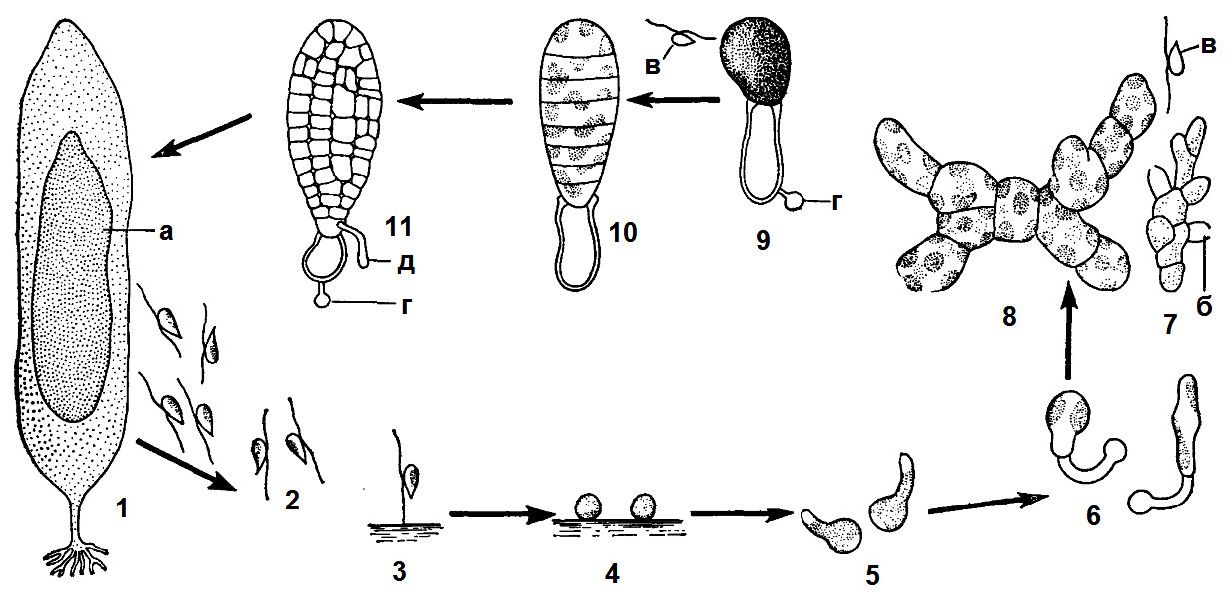
Рис. 6.39. Цикл розвитку ламінарієвих водоростей
1 - талом зі спорами: а - пляма спорангіїв; 2 - плаваючі зооспори; 3 - зооспора прикріпилася переднім джгутиком до грунту; 4 - ембріоспори; 5 - ембріоспори з проростковою трубкою; 6 - утворення перших клітин гаметофітів (зліва - жіночий, справа - чоловічий); 7 - чоловічий гаметофіт: б - антеридій, в - антерозоїди; 8 -багатоклітинний жіночий гаметофіт; 9 - одноклітинний жіночий гаметофіт після виходу яйцеклітини з оогонію: г - оболонка ембріоспори; 10-11 - початок розвитку слані спорофіта: д - первинний ризоїд
Нормальне відтворення їх можливе лише статевим шляхом. Статевий процес — типова оогамія. Статеві органи розвиваються в концептакулах. Із стінки концептакула виростають довгі волоски — парафізи, що заповнюють майже всю його порожнину (рис. 6.40). Особливо довгі волоски розвиваються в жіночих концептакулах, де вони у вигляді пучка видаються з отвору концептакула. Серед цих волосків розвиваються оогонії і антеридії.
Майже усі бурі водорості винятково морські рослини. Найбільші за розмірами їх форми (ламінарієві, деякі фукусові) зустрічаються у морях приполярної і полярної зон. Тут для них сприятливі помірна і низька температура води і підвищений у порівнянні з водами тропічних морів вміст біогенних речовин. Селяться вони переважно на скелях і камінні на глибині 6-15 м.
Рис. 6.40. Спорангії (а) і парафізи
(б) ламінарієвих водоростей
Серед бурих водоростей є види з ефемерними, однорічними і багаторічними таломами. На тривалість існування таломів великий вплив мають екологічні умови. Багаторічні таломи бурих водоростей бувають декількох типів. У одних водоростей талом багаторічний, щороку відмирають тільки пагони, на яких розвивалися органи розмноження ^иса^), у інших ^атіпагіа^) багаторічними є стовбур і органи прикріплення, пластинчаста частина — однорічна.
У деяких тропічних видів саргасових водоростей багаторічним буває лише диск, що служить для прикріплення талому.
У розвитку бурих водоростей спостерігається сезонність. Так, у помірних субарктичних поясах вони найбільше відростають влітку, хоч посилене збільшення їх сланей починається вже наприкінці зими, коли температура води близько 0°С. У тропічних морях масовий розвиток бурих водоростей спостерігається взимку.
Найскладніша будова у представників порядку ламінарієвих.
Роль у природі бурих водоростей надзвичайно велика. Вони є одним з основних джерел органічної речовини в прибережній зоні, особливо в морях помірних і приполярних широт, де їх біомаса може досягати десятків кілограмів на 1 м2. Крім того, зарості бурих водоростей служать місцем розмноження, укриття і живлення бага
тьох прибережних тварин; вони створюють також умови для поселення мікроскопічних і макроскопічних водоростей інших систематичних груп.
Велике і господарське значення бурих водоростей, особливо як сировина для отримання різних речовин (наприклад, альгінатів — солей альгінової кислоти, зокрема альгінату натрію). Цю речовину широко використовують для стабілізації різноманітних розчинів і суспензій. Додавання невеликої кількості альгінату натрію підвищує якість харчових продуктів (консервів, морозива, фруктових соків і т. д.), різноманітних фарб і клеїв. Вони входять до складу змащувальних матеріалів для машин, розчинних хірургічних ниток, мазей і паст у фармацевтичній і парфумерній промисловості.
Бурі водорості використовують також як сировину для отримання маніту, який використовується для виготовлення таблеток і дієтичних продуктів харчування, у виробництві синтетичних смол, фарб, паперу, вибухових речовин, для дублення шкіри, при проведенні хірургічних операцій.
Здавна у медицині бурі водорості використовують для виготовлення замінників крові, препаратів, що запобігають згортанню крові, для виведення радіоактивних речовин з організму.
Бурі водорості, особливо ламінарієві, споживають у їжу. Вони багаті на мікроелементи, і борошно, виготовлене з них, використовується як домішка до корму сільськогосподарським тваринам. У свіжому і переробленому вигляді їх застосовують як добрива.
Інтенсивне використання дикорослих морських макрофітів, зокрема бурих водоростей, призвело до вичерпання їх природних запасів і поставило людство перед необхідністю їх штучного вирощування. Тому в деяких морських країнах у останні 50 років значного розвитку одержала аквакультура водоростей (у Норвегії, Великобританії, Франції, США і ін.). Штучно вирощують види родів Laminaria і Macrocystis.
Відомо близько 1500 видів бурих водоростей. Вони діляться на два класи: феозооспорові і циклоспорові.
Клас феозооспорові (Phaeoгoosporophyceae) об'єднує 11 порядків, найважливішим з яких є ламінарієві.
Порядок ламінарієві (Laminariales). Це найбільші за розміром водорості. Спорофіти ламінарієвих — це рослини висотою до 20-50 м. Талом має паренхіматозну структуру, кілька листкових пласти
нок, розташованих на простому або розгалуженому стовбурі, прикріплюється до ґрунту диском чи ризоїдами (рис. 6.41). У місці переходу листкової пластинки у стовбур знаходиться інтеркалярна меристема, за рахунок якої наростає стовбур і пластинка. Швидкість росту слані 10-14 см на добу. Листкова пластинка у багаторічних представників щорічно відмирає, а зимує лише стовбур з ризоїдами.
У стовбурах багаторічних ламінарій помітні річні кільця.
Рис. 6.41. Морські бурі водорості порядку ламінарієвих:
1 — Macrocystis; 2 — Lessonia; 3 — Postelsia
Спорофіти ламінарієвих утворюють одноклітинні спорангії, що розвиваються серед одноклітинних булавоподібних парафіз.
Гаметофіти багатоклітинні, у вигляді розгалужених ниток. Чоловічі гаметофіти утворюють на окремих клітинах вирости, які розвиваються у антеридії, що формують по одному сперматозоїду. На оогоній може перетворитися будь-яка клітина жіночого гаметофіту. Крізь отвір на верхівці оогонію яйцеклітина виходить назовні, але залишається прикріпленою до краю отвору. Статевий процес — оогамія. Із зиготи розвивається новий макроскопічний спорофіт.
До роду макроцистіс належить понад 20 видів водоростей, поширених переважно в холодних морях, у тому числі гігантська водорість макроцистіс (Macrocystis) до 60 м завдовжки, що росте на узбережжі Тихого океану на глибині 10-20 м.
Рід ламінарія (Laminaria) налічує 30 видів, більшість з яких поширена в північній півкулі; більше всього видів ламінарій у Ти
хому океані. Талом до 22 м завдовжки, розчленований на листкови-дну пластинку, циліндричний черешок і ризоїди, якими кріпиться до субстрату (рис. 6.42). Пластинка у цих водоростей рівна або зморшкувата, без волосків, ціла або розітнута, без отворів, іноді з двома подовжніми рядами вм'ятин і опуклостей або з двома подовжніми складками. Найбільше промислове значення мають ламінарії цукриста ^. saccharina), пальчаста ^. йірйаїа), північна ^. йурегЬогеа), японська (Ь. japonica), вузька (Ь. аг^шШа).

Рис. 6.42. Ламінарія пальчаста (Laminaria digitata)
Клас циклоспорові (Cyclosporophyceae) об'єднує морські водорості. Талом циклоспорових великий (1-12 м), складається з стовбура, підошви і листоподібних утворень. Росте верхівковою клітиною.
Циклоспорові не мають чергування поколінь, а лише змінюють ядерні фази. Нестатевого розмноження немає. Статевий процес — оогамія. Мейоз відбувається при утворенні гамет. З настанням періоду розмноження на кінцевих розгалуженнях талому в особливих заглибленнях — скафідіях — утворюються гаметангії. Скафідії у більшості роздільностатеві. Всередині одних розвиваються антеридії, всередині інших — оогонії. При дозріванні статеві клітини виходять із скафідія і запліднюються у воді. Зигота вкривається оболонкою і без періоду спокою проростає у нову рослину — спорофіт.
Циклоспорові ростуть у всіх морях від Арктики до Антарктики. Усі вони мають промислове значення. Найпоширенішим є рід фукус (Fucus), види якого використовують як добрива, на корм худобі, для виробництва борошна, альгінатів.
Клас поділяють на 3 порядки, найважливіший з яких порядок фукусові (Fucales). Талом досить великий, різноманітної форми,
складної морфологічної і анатомічної будови, розчленовується на основну, циліндричну або пластинчасто-стрічкоподібну частину, зазвичай короткий стовбур і конусовидну або дисковидну підошву. Основна частина талому найчастіше дуже розгалужена або розчленована, шкіряста або м'ясиста.
Органи розмноження розвиваються в концептакулах, розкиданих по всьому талому або сконцентрованих в певних місцях, — у реце-птакулах. Мешкають у всіх морях від Арктики ка Антарктики, крім Каспійського і Аральського.
Саргасоеі водорості (Sargassaceae) мають кущевидну слань довжиною близько 1-2 м (іноді до 10 м). Основа і стовбур часто багаторічні, гілочки однорічні, ростуть з пазух листовидних пластинок. Є понад 150 видів, які ростуть на дні біля берегів теплих морів. У західній частині Атлантики відомі величезні плаваючі скупчення саргасових водоростей, що колись відірвалися від субстрату і розмножувалися вегетативно. Ця акваторія відома під назвою Саргасове море. Саргасові водорості використовують для отримання альгінатів; деякі види в Південно-східній Азії вживають в їжу.
З бурих водоростей у Чорному морі найчастіше зустрічається цистозейра бородата (Cystoseiгa ЬагЬаїа). Її слань має вигляд великих, сильно розгалужених кущиків, рясно вкритих гілочками, схожими на листя папороті. У вертикальному положенні цистозейру підтримують повітряні бульбашки, що розташовані на верхніх частинах гілочок і виконують роль поплавків. Мешкає вона на каменях і скелях, до яких дуже щільно прикріплюється виростом на нижній частині стовбура — підошвою. Під час штормів частина особин відривається і викидається на піщані береги, де її можна спостерігати практично цілий рік.


2 З
Рис. 6.43. Фукусові: 1 - фукус (Fucus sp.), 2 - саргаса плаваюча (Sargassum natans), 3 - цистозейра бородата (Cystoseira barbata)
Це найчисленніша група водоростей, що об’єднує понад 20 тис. видів, яким характерне зелене забарвлення, зумовлене наявністю в хлоропластах хлорофілів а і в, а-, [3-, у-, ^-каротинів і близько десяти різних ксантофілів. Пігментний склад зелених водоростей подібний до вищих рослин.
Зелені водорості бувають одноклітинні, колоніальні, ценобіальні та багатоклітинні. їм властиві майже всі типи морфологічної структури водоростей, крім амебоїдної і харофітної, — монадна, кокоїд-на, пальмелоїдна, нитчаста, пластинчаста, сифональна. Зелені водорості мають розміри від 1-2 мкм у одноклітинних форм до кількох десятків сантиметрів.
Монадні форми і стадії мають різну кількість джгутиків: від одного у пединомонади (Pedinomonas) до багатьох (монадні стадії едогонію і ін.).
У більшості зелених водоростей утворюються клітинні оболонки, що зберігають більш менш постійну форму КЛІТИН.
В оболонці багатьох зелених водоростей є два шари — внутрішній - целюлозний і зовнішній - пектиновий. Проте у деяких водоростей оболонка містить більшу кількість шарів, наприклад, оболонка гематокока (Наетаіюсоссш), за даними електронної мікроскопії, складається з чотирьох шарів (рис. 6.44).
Рис. 6.44. Гематокок (Haematococcus)
Оболонки можуть бути інкрустовані солями заліза або кальцію.
У видів роду едогоній (Оейоропіит) на поверхні оболонки виділяється додатковий твердий покрив (кутикула), що складається з кутину.
Клітини примітивних зелених водоростей не мають відокремленої від цитоплазми оболонки, вони оточені лише плазмалемою.
Обмін клітин з навколишнім середовищем відбувається через пори, що утворюються унаслідок неповного змикання оболонок при цитокінезі (первинні) або під впливом ферментів (вторинні пори). У деяких колоніальних і багатоклітинних зелених водоростей через пори зв'язок між сусідніми клітинами здійснюється за допомогою плазмодесм, у інших — через пори виділяється слиз, який сприяє утворенню колоній.
Клітини зелених водоростей містять від одного (Chlamydomonas) до кількох сотень (Cladophora) ядер. Хлоропласт у клітині один або їх багато. Часто в хлоропластах є піреноїди.
У хлоропластах знаходяться фотоасиміляційні пігменти, що обумовлюють зелене забарвлення водоростей. Проте у деяких видів, що мешкають в екстремальних умовах (на снігу, в ропі гіперга-линних водойм) спостерігається інтенсивне утворення каротину, що маскує зелений колір хлорофілів і що обумовлює оранжево-червоне забарвлення клітин (Chlamydomonas nivalis, Dunaliella salina).
Запасною поживною речовиною у більшості зелених водоростей, як і у вищих рослин, є крохмаль, рідше олія.
Розмноження вегетативне, нестатеве, статеве, за допомогою вегетативних і спеціалізованих клітин. Вегетативне розмноження у одноклітинних водоростей відбувається поділом клітини надвоє, у колоніальних і багатоклітинних — фрагментацією талому. Ценобіа-льні (Volvox, Hydrodictyon, Scenedesmus) утворюють дочірні ценобії усередині клітин материнського ценобію. Багато зелених водоростей розмножуються за допомогою акінет.
Нестатеве розмноження здійснюється за допомогою спор. Спори бувають рухомими (зооспори) і нерухомими (апланоспори).
Статеве розмноження представлено різноманітними типами і формами: хологамія, ізогамія, гетерогамія, оогамія, кон'югація. Зигота проростає після мейотичного поділу, тому більшість зелених водоростей у вегетативному стані гаплоїдні. Диплоїдною є лише зигота. Деякі водорості характеризуються зміною поколінь.
Зелені водорості широко розповсюджені у воді, ґрунті, в наземних екотопах на різних географічних широтах, у всіх морях і на всіх континентах земної кулі. Серед них є планктонні, перифітонні і бе-нтосні організми. Є епіфіти, паразити і симбіонти. Усі зелені водорості, крім вольвоксових, у вегетативному стані нерухливі.
Масовий розвиток мікроскопічних зелених водоростей може викликати зелене або червоне «цвітіння» води, ґрунту, снігу тощо.
Відділ поділяється на три класи: справжні зелені водорості, сифонові і кон'югати.
Клас справжні зелені водорості (Chlorophyceae). Це різноманітні зелені водорості монадної, гемімонадної, кокоїдної, сарциноїд-
ної, нитчастої, різнонитчастої, паренхіматозної, псевдопаренхімато-зної, сифональної і сифонокладальної структури, одно-, багатоклітинні або неклітинні, одиночн або в угрупованнях різного типу, мі-кро- і макроскопічні, прикріплені і неприкріплені. Нестатеве розмноження вегетативне або за допомогою спор різної будови, включаючи зооспори. Статевий процес холо- або мерогамія; остання представлена ізо-, гетеро-або оогамією. Статеві органи одноклітинні, у драпарнальдієвих (Drapamaldiella) - багатогніздні.
Порядок вольвокальні (Volvocales). До цього порядку належать одноклітинні, колоніальні, ценобіальні організми, рухливі у вегетативному стані. Є понад 1000 видів. Клітини дво-, чотириджгутикові, одноядерні, вкриті оболонкою, рідко голі.
Хлоропласти мають різну форму, у більшості є піреноїди.
З пластидами пов'язане світлочутливе вічко (стигма) різноманітної форми, зазвичай яскраво-червоного кольору.
Характерним органоїдом всіх прісноводних вольвоксових є скоротливі (пульсівні) вакуолі.
Від переднього кінця клітини відходять джгутики.
Колонії водоростей утворюються як за допомогою слизу, так і без нього. Колоніальні вольвоксові утворюють ряд, що поступово ускладнюється, від простих колоній, що випадково виникають у процесі поділу клітин (Dangeardinella), до ценобіїв, як у пандорини (Pandorina) з генетично обумовленим числом клітин, і до великих ценобіїв вольвоксу (Volvox), що включають десятки тисяч сполучених плазмодесмами клітин, що реагують як цілісний організм, в якому спостерігається складна диференціація клітин на вегетативні і генеративні (рис. 6.45)
Рис. 6.45. Невелика ділянка колонії Volvox aureus (схематизовано):
1 — вегетативна особина колонії; 2 — цитоплазматичний місток; 3 — більша особина, в результаті розвитку якої утворюються нові дочірні колонії
У видів вольвоксу (Уоіяох), крім того, спостерігається диференціація особин на нестатеві і статеві, останні можуть бути двостатевими або роздільностатевими. Складно побудовані колонії вольвоксу іноді розглядають як багатоклітинні організми. Проте від багато клітинних рослин вольвокс відрізняється нездатністю до вегетативного поділу клітин і необмеженого росту протягом вегетативної фази циклу розвитку.
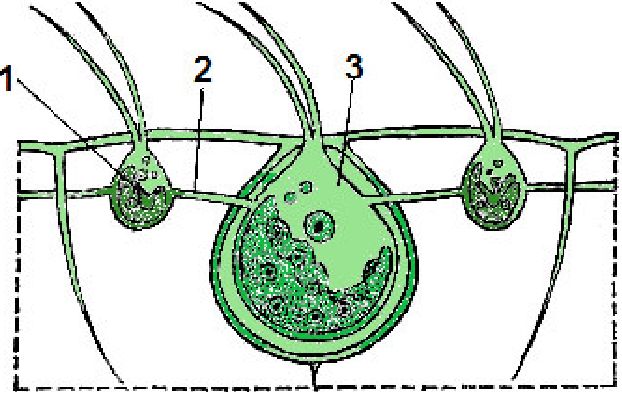
Вегетативне розмноження відбувається поділом клітини, статеве — способами хологамії, ізогамії, гетерогамії, оогамії, нестатеве — зооспорами. З зиготи найчастіше утворюються чотири зооспори, які дають початок новим особинам.
Переважна більшість вольвоксових харчується фотоавтотрофно, рідше сапротрофно. Вони поширені переважно у прісних водах, є мешканцями мілких, стоячих водойм, забруднених органічними речовинами (калюжі, канави, болота, відстійники, ставки), де розвиваються часто у величезній кількості, забарвлюючи воду в зелений колір. Деякі види викликають червоне «цвітіння» солоних водойм і снігу. У морях представлені одиничні види.
Вольвоксові беруть активну участь у самоочищенні забруднених вод, є важливою ланкою в трофічному ланцюзі біоценозів. Це перспективні об'єкти біотехнології як продуценти гліцерину, Р-каротину, аскорбінової кислоти тощо.
Найкраще вивченими є три підпорядки вольвоксових: полібле-фаридові, хламідомонад ові, вольвоксові.
Підпорядок поліблефаридові (РоІуЬІеркаСНаЄ) включає одноклітинні водорості, які не мають сформованої клітинної оболонки і вкриті тонким перипластом.
Найвідоміша з цього порядку дуналіела солоноводна поширена на півдні України у водоймах з різним рівнем солоності — від майже прісних до перенасичених. У процесі життєдіяльності в клітинах дуналіели нагромаджуються каротиноїди, що надають їм рожево-жовтого або цегляно-червоного забарвлення, чим пояснюється червоне «цвітіння» водойм. Ця водорість є джерелом одержання каротину (провітаміну А).
До підпорядку хламідомонадові (ССІатуНотопаНаЄ) відноситься рід хламідомонада (Chlаmydomonаs), що включає близько 500 видів. Це одноклітинні дводжгутикові водорості з м'якою пек-тиново-геміцелюлозною оболонкою, що спереду утворює помітний вигін — носик, від якого відходять два джгутики рівної довжини. У цитоплазмі знаходиться чашоподібний хлоропласт з піреноїдом і добре помітним червоним вічком (стигмою) у верхній частині, ядро і дві пульсівні вакуолі (рис. 6.46).

Види роду хламідомонада (Chlamydomonas) селяться в калюжах і невеликих водоймах, забруднених органічними відходами, спричиняють їх зелене «цвітіння». В Україні поширені хламідомонади Рейнхарда (Сй. геіпсйагйіі), пронизана (Сй. рейша) та ін.
Рис. 6.46. Будова хламідомонади:
1 - хроматофор; 2 - цитоплазма; 3 - вічко; 4 - джгутики; 5 - вакуолі; 6 - ядро; 7 - піреноїди
Підпорядок вольвоксові (Volvocales) об'єднує понад 1000 видів одноклітинних або колоніальних, зокрема ценобіальних, переважно мікроскопічних зелених водоростей, представлених індивідами мо-надної організації, здатних активно пересуватися протягом вегетативної фази життєвого циклу за допомогою джгутиків.
Лише деякі колоніальні вольвоксові можуть досягати макроскопічних розмірів, наприклад, колонії Volvox до 2 мм в діаметрі.
Клітинна оболонка переважно багатошарова, складається з глікоп-ротеїну; целюлоза не виявлена. Відкладення солей кальцію, заліза або марганцю додає оболонці твердість, перетворюючи її на своєрідний будиночок (Coccomonas).
Серед ценобіальних форм найпоширеніші роди гоніум (Gonium), евдорина (Eudorina) і пандорина (Pandorina) (рис. 6.47). Ценобієм називається багатоклітинне тіло, утворенне клітинами однієї генерації, з'єднаних між собою в певному порядку. При розмноженні кожна клітина ценобію утворює самостійний дочірний ценобій.
У гоніуму ценобій пластинчастий, кулястої, чотирикутної форми. У водоймах України поширений гоніум грудний (G. pectorale). Евдорина має еліпсоїдний ценобій. У водоймах України поширений вид евдорина елегантна (Е. elegans).
У пандорини (Pandorina) ценобій кулястий. В Україні поширений планктонний вид пандорина тутова (P. morum).
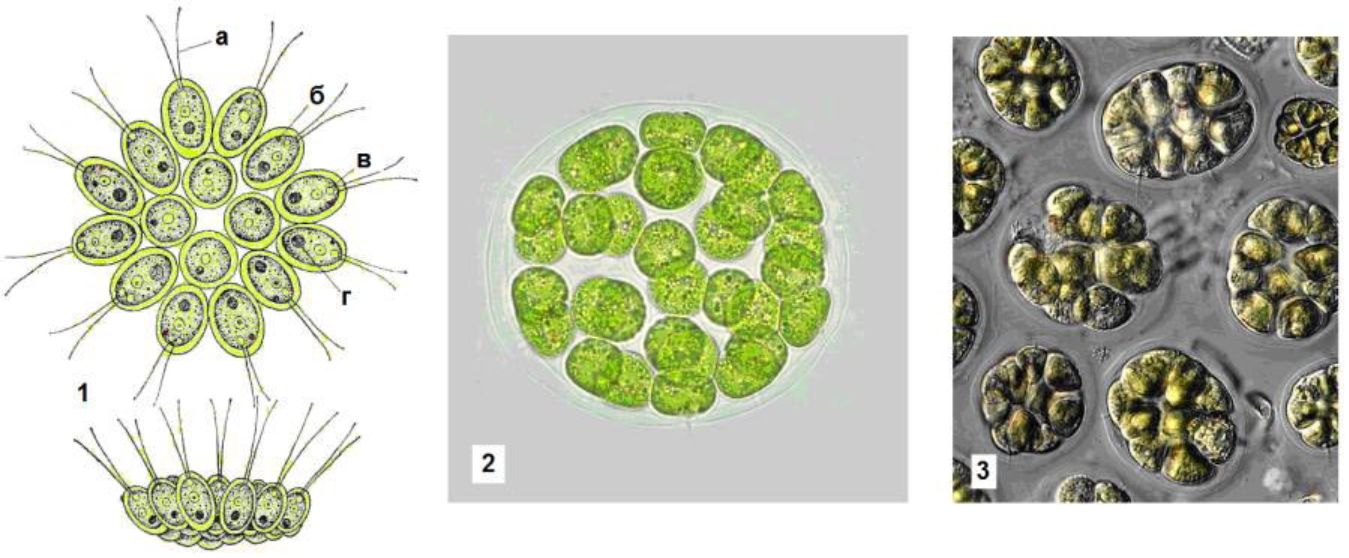
Рис. 6.47. Ценобіальні вольвоксові: 1 - Gonium pectorale (зверху — вигляд колонії зверху: а — джгутики; б — вічко (стигма); в — скоротлива вакуоля; г — ядро; знизу — вигляд колонії збоку); 2 - Eudonna; 3 - Pandonna moram
Крім нестатевого розмноження, у ценобіальних форм наприкінці вегетації здійснюється статевий процес — ізогамія у гоніумів, гетерогамія у пандорин і оогамія в евдорин.
До колоніальних вольвоксових форм належить рід вольвокс (Volvox). Колонії вольвоксу кулясті, до 2 мм у діаметрі, в периферійному шарі яких розташовано до 20 і навіть 50 тис. хламідомона-доподібних клітин, що зрослися між собою бічними ослизненими стінками. В межах колонії спостерігається диференціація і спеціалізація клітин. Між вегетативними клітинами розміщуються більші за розмірами репродуктивні.
Рис. 6.48. Розмноження вольвоксових (всередині материнського ценобію фор муються дочірні)
Найпоширеніший у прісноводних водоймах вольвокс кулястий (V. рІоЬаїог). Вольвоксові дали початок протококовим водоростям.
Порядок хлорококових (СЬІогососса^). Більшість хлорококо-вих водоростей має талом кокоїдної структури. Це одноклітинні, ценобіальні, зрідка колоніальні і навіть багатоклітинні (нитчасті і
пластинчасті) водорості (рис. 6.49). Клітини хлорококових за будовою подібні до вольвоксових, але не мають джгутиків і нерухливі у вегетативному стані. Рухливі лише зооспори і гамети. Клітина має пектинову, целюлозну, часто ослизнену на поверхні оболонку, одноядерна, рідше, багатоядерна, хлоропласт чашоподібний з піреної-дом. Є вакуоля. Пульсівні вакуолі є лише у нижчих представників і в зооспорах. Запасні поживні речовини — крохмаль, рідше олія.


Рис. 6.49. Хлорококові: 1 - Scenedesmus, 2 - KirchnerieИa, 3 - Desmodesmus, 4 - Chlorococcum
У прісних водоймах хлорококові утворюють планктон, бентос на підсихаючому грунті, корі дерев. Зазвичай планктонні види мешкають на глибині 0,5—1 м. Найсприятливіші для розвитку хлорококових водоростей стави, особливо рибницькі, де вони є однією з домінуючих груп. Найчастіше в рибоводних ставах спостерігається одночасна вегетація декількох видів, що створюють домінуючий ценоз конкретної водойми. «Цвітіння» води можуть викликати види родів Scenedesmus, Pediastrum, CrucigenieПa та інші.
Найпоширенішими родами серед протококових є представники родів хлорокок, хлорела, протокок, гідродикціон.
Хлорокок зустрічається в прісних водоймах, на грунті, корі дерев, в таломі лишайників. Клітини кулясті, поодинокі
або зібрані купками, вкриті целюлозною оболонкою, одноядерні. Хлоропласт глибокочашоподібний з піреноїдом. Розмноження здійснюється тільки зооспорами.
Хлорела (Chlorella) — одноклітинна водорість, поширена в тих же місцях, що і хлорокок. Клітини зелені, кулястої форми, 15 мкм у діаметрі. Хлоропласт чашоподібний з одним піреноїдом. Ддро одне. Розмножується автоспорами, які утворюються по 4-8 в одній клітині. Хлорела невибаглива до умов життя і швидко розмножується, завдяки чому є об'єктом масового культивування для практичного використання в різних галузях. Хлорелу широко культивують як харчову і кормову культуру. З одного гектара водної поверхні можна зібрати до 30 т сухої органічної речовини. У біомасі хлорели виявлено більше десяти різних вітамінів.
Протокок (Protococcus) — одноклітинна водорість, що живе на корі дерев, а також у грунті. Розмножується прямим поділом клітини, який відбувається у двох взаємно перпендикулярних напрямах, завдяки чому може утворювати пластинчасті, різнонитчасті таломи.
Водяна сіточка (Hydrodictyon) зустрічається в тихих забруднених затонах річок, ставах, ямах. Ця водорість має вигляд сітчастого мішка, що досягає розмірів до 1 м (рис. 6.50). Стінка мішка утворена довгими (до 1,5 см) циліндричними клітинами, з'єднаними
кінцями в п'яти-, шестикутники.
Порядок улотриксові (Ulothrichales) включає близько 460 видів водоростей, що належать до 70 родів. Це переважно багатоклітинні водорості нитчастої, різнонитчастої або пластинчастої структури, часто макроскопічні, до декількох дециметрів у довжину, забарвлені переважно в зелений колір різних відтінків.
Таломи живуть вільно або прикріплюються до субстрату за допомогою базальної клітини, ризоїдів або підошви.
Рис. 6.50. Водяна сіточка (Hydrodictyon)

Переважна більшість улотриксових мешкає в прісних проточних і стоячих водах, значно менше видів зустрічається в лужних, вапняних і кислих водах, а також у солонуватих водоймах і морях. Улот-риксові водорості переважно прикріплені, зрідка пасивно плавають у товщі води. Переважна більшість улотриксових епіфіти, деякі — ендофіти, що зберігають самостійне автотрофне живлення.
Своєрідну екологічну групу складають ендолітофітні організми, які, проникаючи у вапняний субстрат, розрихлюють його, і він легко піддається дії фізичних і хімічних чинників.
Нарівні з багатьма іншими групами водоростей улотриксові відіграють певну роль в процесах природного самоочищення вод. Деякі види, які чутливо реагують на гідрохімічні зміни середовища, використовують як біологічні індикатори для швидкого визначення ступеня забрудненості води.
Підпорядок улотриксові (Ulothrichales). Мають слань у вигляді однорядної нерозгалуженої нитки, що прикріплюється базальною клітиною до субстрату. Центральним тут є рід улотрикс (Шоїйгіх), види якого зустрічаються в прісних гірських струмках, річках і в прибережній зоні озер (рис. 6.51).
Рис. 6.51. Улотриксові: 1 - улотрикс (ИІоШгіх), 2 - ульва салатна (Иіяа Іасіюса)
Підпорядок ульвові (Шяіпеае). Це макроскопічні багатоклітинні водорості, що мають паренхімну будову. їх талом являє собою двошарову пластинку або має мішкоподібну, трубчасту структуру. Талом у нижній частині звужений і має ризоїди, за допомогою яких водорості прикріплюються до субстрату. Поширені ульвові переважно в морях. До порядку відносяться дві родини -
Mоnоstrоmаtасeаe і Иіяасеае.
У Чорному морі зустрічаються види ульви (Иіуа гі§іПа, и. Іасіцса). Ульва салатна (и. Іасіцса), відома під назвою морського
салату, вживається в їжу (див. рис. 6.51).
Підпорядок хетофороеі (СИае^рИогіпеае). Водорості цього порядку мають різнонитчасту слань, що складається з однорядних ниток. Нитки розгалужуються, і вся слань диференціюється на систему сланких і вертикально розгалужених ниток. Нитки до кінців потоншуються і закінчуються тонким волоском — хетою, звідки і походить назва порядку.
Представниками родини хетофорових є рід хетофора (Chaetophora), види якого поширені в прісних водоймах.
Порядок кладофорові (СіайоркогаО). Порядок об'єднує понад 100 видів водоростей, що відносяться до 15 родів. Це багатоклітинні водорості у вигляді довгих або коротких простих або розгалужених ниток, що ростуть одиночно або зібрані у скупчення, прикріплені протягом всього життя або пов'язані з субстратом лише на початковій стадії розвитку, а пізніше відриваються і живуть вільно в товщі води або у вигляді скупчень переплутаних ниток («твані») на дні водойм. Деякі види (наприклад, Сіайорйога аераргоріїа) утворюють плаваючі кулясті скупчення до 10-20 см в діаметрі, що складаються з розгалужених ниток. Кладофорові зазвичай забарвлені в зелений колір різних відтінків.
Власне нестатеве розмноження здійснюється за допомогою зооспор. Зооспори грушовидні, дво-, чотирижгу-тикові, формуються в майже незмінених вегетативних клітинах — зооспорангіях (рис.
6.52).
Статеве розмноження здійснюється двожгутиковими гаметами, які виникають в незмінених материнських клітинах — гаметангіях. Статевий процес переважно ізогам-ний.
Деякі види родів кладофора (Сіайорйога) і хетоморфа (Сйаеіютогрйа) утворюють таломи різної статі і злиття гамет відбувається тільки в тому випадку, якщо вони розвинулися на різних
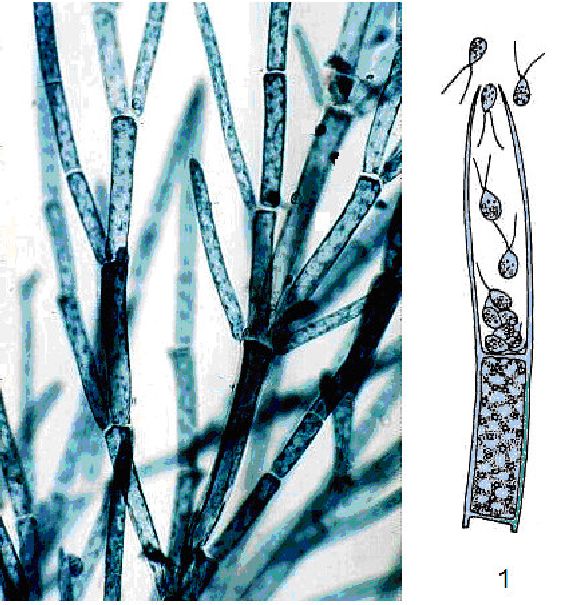
Рис. 6.52. Кладофора (СІагіорИога §іошега1а): 1 - вихід зооспор таломах.
У результаті злиття гамет утворюється зигота, яка проростає відразу після утворення або після деякого періоду спокою.
Мейоз відбувається перед утворенням зооспор.
Кладофорові водорості широко розповсюджені в природі. їх можна виявити всюди, за винятком найпівнічніших широт. Вони мешкають в різноманітних прісних і солонуватих водоймах, а також у морях. У водоймах помірних широт кладофорові вегетують, як правило, в теплий період року. Переважно ведуть прикріплений спосіб життя, зустрічаються на листі і стеблах водних квіткових рослин, деревині, каменях, черепашках молюсків, різних гідротехнічних спорудах.
Масовий розвиток кладофор згубно впливає на рибне господарство. їх нитки іноді утворюють природні шари завтовшки 20—50 см. Вони пригнічують ріст корисної м'якої рослинності, а в хащах цих ниток заплутується і гине молодь риб.
Істотна і позитивна роль кладофорових. Вони служать кормом для численних безхребетних тварин і цінних промислових риб. Велика їх роль в збагаченні води киснем і очищенні її від забруднень.
Найчастіше в прісних і солоних континентальних водоймах зустрічаються представники родів Сйаеіютогрйа, Rhizoclonium, Сіаборйога, Атоібіеііа.
В Україні найпоширеніша кладофора (Сіайорйога glomerata), яка утворює скупчення на дні прісних водойм. У Чорному морі широко розповсюджена Сіайорйога йаїтайса.
Порядок едогонієві (ОеНоропіаЄ). Ці водорості мають простий (Оейоропіит) або розгалужений (Ви1Ьосйаеі;е) нитчастий талом, прикріплений до субстрату. Таломи можуть досягати в довжину до 50 см, але зазвичай не перевищують декілька міліметрів.
Нестатеве розмноження здійснюється зооспорами, які утворюються по одній в кожній клітині.
Статеве розмноження у едогонієвих водоростей завжди оогамне, причому утворення жіночих і чоловічих статевих клітин може відбуватися на одній і тій же рослині (однодомні види) або на різних рослинах (дводомні види). За будовою чоловічих рослин дводомні види можна розділити на дві групи: 1) з чоловічими талломами, аналогічними за будовою жіночим; 2) з карликовими чоловічими одноклітинними і багатоклітинними рослинами — наннандріями.
Антеридіальні клітини як у однодомних, так і у дводомних видів за формою відмінні від вегетативних (рис. 6.53).
Вони значно коротші за вегетативні клітини, зустрічаються одиночно або групами. У кожному з антеридіїв утворюється один або два сперматозоїди. Оогонії зазвичай більші за вегетативні клітини, в кожному з них розвивається по одній ооспорі. Оогонії можуть
розташовуватися одиночно « ^
Рис. 6.53. Едогонш (Оеаоропшт sp.): а - аоо групами. зигота, б - антеридій, в - оогоній
Запліднення у едогонієвих
водоростей — складний багатоступінчастий процес. Наприклад, у Оейоропіиш borisianum, андрозооспори хемотаксично притягуються до ініціальної клітини оогонію, розвиток якого починається тільки після завершення розвитку наннандрію; що вже сформований оогоній виділяє масивну слизисту оболонку. Запліднена яйцеклітина перетворюється на зиготу (ооспору), яка зазвичай оточена товстою тришаровою оболонкою.
Після періоду спокою ооспора ділиться мейотично і проростає, утворюючи чотири мейозоспори, які, проплававши якийсь час, прикріпляються до субстрату і проростають у нові нитки.
У деяких випадках едогонієві можуть розвиватися масово і засмічують водойми. Масовий розвиток цих водоростей частіше відбувається в заплавних водоймах, ставах і в евтрофних озерах. На Україні найбільше видове багатство едогонієвих водоростей (видів родів Оейоропіиш і ВиІЬосйаеїе) виявлене в евтрофних болотах і дрібних, періодично пересихаючих водоймах, найменше — в річках і оліготрофних болотах.
Клас сифонові ^ірЬопорЬусеае). Це дуже древня група зелених водоростей, що мають сифональну (неклітинну) будову. Талом їх багато- чи одноядерний. Для внутрішньої будови талому сифонових водоростей характерна центральна вакуоля і пристінний шар цитоплазми, в якому розташовані одне або кілька ядер і один або кілька
хлоропластів. Більшість сифонових (90%) — це морські організми.
Сифонові — одна з найстародавніших груп морських водоростей, що пережила свій розквіт у віддалені геологічні епохи. Сучасні представники є залишками стародавньої, колись могутньо розвиненої флори цих водоростей. В даний час їх налічується біля 400— 500 видів, що відносяться до 60 родів.
Сучасні представники морських сифонових водоростей — зазвичай макроскопічні, деякі до 0,5 м і більше, відрізняються не лише значною різноманітністю зовнішнього вигляду, але і дуже складним розчленуванням тіла на стебло-, листко- і кореневищеподібні частини. Прісноводні сифонові водорості зазвичай помітні неозброєним оком, рідше мікроскопічні, мають вигляд малорозгалужених ниток, іноді одиночних округлих клітин, здатних утворювати довгі, трубчасті, позбавлені перегородок вирости.
У сифонових водоростей спостерігається нестатеве і статеве розмноження. Вегетативне розмноження відбувається шляхом фрагментації талому, за допомогою вивідкових бруньок, що виникають на верхівках гілочок, зрідка акінетами. Власне нестатеве розмноження здійснюється зооспорами з двома і більш джгутиками (у деяких сифонових зооспори мають віночок джгутиків на передньому безбарвному кінці клітини), зрідка апланоспорами; у деяких видів нестатеве розмноження відсутнє.
Статеве розмноження представлене ізо-, гетеро- і оогамією. Зигота проростає без стадії спокою (рис. 6.54).
У деяких сифонових статеве розмноження відсутнє.
Сифонові поширені переважно в морях, і лише деякі представники проникли і розселилися у прісних водах.
У класі є три порядки: сифонові, дазикладові, сифоно-кладові.
До порядку сифонових, або бриопсидових (Вгуорхійаіея), належать 5 родин, 3 з яких (Bryopsidaceae, Саиіеграсеае і Сойіасеае) поширені переважно в морях, а представники родин Prоtоsірhоnасеае і

Рис. 6.54. Дихотомосифон
(БісЬоЩшозірЬоп): а - антеридій, б - оогоній
Dichotomosiphonaceae мешкають у прісних водах помірних широт.
Морська водорість каулерпа (Саиіегра) в довжину досягає до 50 см (рис. 6.55). Слань розчленована на стеблоподібну, листоподібну і ризоїдну частини, в середині яких є численні целюлозні перетинки (трабекули), що утворюють внутрішній скелет і механічно зміцнюють рослину.
1
Рис. 6.55. Бриопсидові: 1 - Саиіегра реІШШ, 2 - Сосішт ІотеЩозит
Порядок дазикладові (Dasycladales). Мають слань, що являє собою одноядерну гігантську клітину складної морфології. Вона складається з нерозталуженої центральної осі, на якій розташовані кільця гілок, а в нижній частині — розгалужені ризоїди. Численні хлоропласти дископодібні, еліптичні без піреноїдів. Статевий процес ізогамний.
Найвідоміший рід — ацетабулярія (АсеіаЬиІагіа). У стерильному стані представники його складаються з одноклітинного стовбура до 10 см завдовжки, прикріпленого до субстрату нижнім кінцем за допомогою ризоїдів (рис. 6.56). На вершині цієї осі поступово утворюється перше кільце бічних сегментів. До кінця першого року вегетації вертикальна частина талому відмирає, залишається лише багаторічна ризоїдна система. На другий рік з ризоїдів знову виростає одна або декілька стерильних осей з кільцями бічних гілочок. Цей процес може повторюватися декілька сезонів, поки не наступить час плодоношення. Тоді утворюються специфічні фертильні (плодоносні) пагони з великих одноклітинних мішковидних гілочок — гаметангіїв, що зростаються своїми боками в парасольку. У га-метангіях утворюються товстостінні багатоядерні цисти із спеціальною кришкою. Зрілі цисти містять безліч ізогамет. При руйнуванні стінок гаметангію цисти виходять в зовнішнє середовище і з них вивільняються гамети. Зигота проростає без періоду спокою.


Рис. 6.56. Сифонові (рід Acetabularia): 1- 8 — ранні стадії розвитку; 9 — доросла рослина, парасолька утворена зрощеними гаметангіями; 10 — вихід гамет із
зрілої цисти
Ацетабулярія середземноморська (Acetabularia тейііеггапеа) зустрічається у Чорному морі.
Клас кон'югати (Соці^аІорЬуевав) - одна з найбільших груп зелених водоростей, представлена переважно прісноводними організмами. До класу відносяться близько 4700 видів переважно мікроскопічних одноклітинних, колоніальних і багатоклітинних нитчастих водоростей. Нитки не розгалужені і не прикріплені.
Найяскравіша відмінна риса кон'югат, що дозволяє об'єднати їх в особливий клас, — повна відсутність рухомих стадій розмноження, тобто зооспор і рухомих гамет, і наявність своєрідного статевого процесу, що одержав назву — кон'югація.
Більшість кон'югат — це одноклітинні, симетрично побудовані водорості (Mеsotаеniаlеs, Dеsmidiаlеs) яскраво-зеленого, смарагдового, рідше темно-зеленого кольору. Деякі види кон'югат особливо
круглі форми, мають в діаметрі 10 мкм і навіть менше, тоді як паличкоподібні форми можуть досягати в довжину 1 мм і більше, і їх видно неозброєним оком.
Більшість одноклітинних кон'югат мають клітини перетягнуті посередині, і складаються з двох частинок (десмідієві). У нитчастих форм клітини не перетягнуті. Клітинна оболонка целюлозна, суцільна або сегментована на дві й більше частин, без пор чи з добре помітними порами, гладенька чи з багатою орнаментацією, тому кон'югати вважають найгарнішими водоростями.
Клітини одноядерні. Розміри ядер дуже різноманітні і, як правило, залежать від розмірів клітин. Дуже велике, гомогенне, з яскраво помітним ядерцем посередині, ядро у видів роду Spiгogyгa (Гурпетаіа^). Хлоропластів декілька, різної форми з піреноїдами.
Розмноження кон'югат здійснюється нестатевим і статевим шляхом. Статеве розмноження у кон'югат здійснюється в результаті дуже своєрідного статевого процесу — кон'югації (рис. 6.57). Під час кон'югації відбувається злиття протопластів двох клітин, які виконують тут функцію гамет. У результаті злиття, що відбувається через канал, який спеціально утворюється, виходить одноклітинна, покрита товстою оболонкою зигоспора. Дві клітини, що зливаються (кон'югуючі), зовні абсолютно не відрізняються; фізіологічна диференціація на чоловічу і жіночу клітини хоч і визначена, але дізнатися її можна лише опосередковано за положенням зигоспори.
Рис. 6.57. Конюгація: 1 - у зигнеми (Гурпета), 2 - у спірогіри (Брігоруга)
Іноді зигота утворюється в кон'югаційному каналі, розростаючись аж до стінок кон'югуючих клітин — гаметангіїв. У цьому випадку розділення клітин на чоловічу і жіночу умовне, а сам процес злиття гамет називається ізогамією. Ця форма статевого процесу
характерна для більшості кон'югат. У розвиненіших в еволюційному відношенні зигнемових і десмідієвих поряд з ізогамією існує і фізіологічна анізогамія, або гетерогамія. У цьому випадку одна гамета (чоловіча) активно перетікає через кон'югаційний канал у приймаючу клітину, де і зливається з порівняно нерухомою гаметою (жіночою). В цьому випадку зигота формується тільки в приймаючій (жіночій) клітині.
Після утворення зиготи її оточує тришарова оболонка, і вона перетворюється на зигоспору. Зигоспора після тривалого періоду спокою, який може продовжуватися всю зиму і довше, проростає, даючи початок новим організмам — проросткам (від 1 до 4).
У багатьох видів десмідієвих водоростей зигоспори не виявлені. Кон'югація триває 7—14 днів.
Поширені кон'югати в прісних водоймах, на мохах, скелях, вологому грунті і на високогірних льодовиках.
Багато видів кон'югат використовують в їжу різні безхребетні тварини, а також риби. Деякі види масово розмножуються в стічних водах, сприяючи їх очищенню, інші — здатні виділяти біологічно активні речовини, подібні до пеніциліну.
Поділяють цей клас на чотири порядки: мезотенієві, гонатозиго-ві, зигнемові і десмідієві, з них найпоширеніші два останні.
Порядок зигнемові (Гурпет^аО) включає близько 700 видів нитчастих, нерозгалужених, не прикріплених водоростей. Нитка утворена циліндричними одноядерними клітинами з ослизненою оболонкою. Хлоропласти - осьовий пластинчастий, осьовий зірчастий або пристінний спірально-стрічкоподібний (рис. 6.58).

Рис. 6.58. Хлоропласти зигнемових: 1 - спірогіра ^рігоруга sp.) зі спіральними хлоропластами, 2 - зірчасті хлоропласти зигнеми (Гурпета sр.), 3 - пластинчастий хлоропласт у мужоції (Моиреоііа sр.)
Розмноження вегетативне — розривом нитки на частини або статеве — кон'югацією.
Поширені зигнемові в прісних водоймах всіх континентів.
Спірогіра ^рігоруга) являє собою нитку, утворену витягнутими циліндричними одноядерними клітинами, з одним або кількома стрічкоподібними спіральними хлоропластами з піреноїдами. Утворює зелену твань у водоймах з повільною течією.
За будовою зигнема (Гурпета) подібна до спірогіри, але в цитоплазмі її клітин знаходяться по два зірчастих хлоропласти з піреної-дом в центрі кожного. Між хлоропластами знаходиться ядро, оточене цитоплазмою. Поширена на болотах і заболочених водоймах.
На відміну від спірогіри мужоція (Моиреоїіа) має один пластинчастий хлоропласт, розташований у центрі клітини.
Порядок десмідієві (Desmidiales) об'єднує понад 4 тис. видів. Ці водорості характеризуються дивовижною різноманітністю контурів, красою форм і чудовою симетрією (рис. 6.59). Химерність контурів клітин у поєднанні з різноманітною скульптурою і зеленими хлоропластами завжди привертали до себе увагу не лише науковців, але і любителів, завдяки чому ці водорості є найкраще вивченими у всьому світі.
Клітини десмідієвих складаються з двох симетричних частинок — напівклітин, з'єднаних між собою вузькою частиною — перешийком. Зовнішній вигляд клітин у більшості випадків залежить від того, з якого боку їх роздивляються під мікроскопом. Клітини десмідієвих мають три площини симетрії. Різноманітність контурів клітини при розгляді з різних боків має велике значення в систематиці десмідієвих. Деякі види мають дуже схожі контури при розгляді їх спереду, але при цьому різко відрізняються один від одного при розгляді збоку або зверху. Своєрідною будовою відрізняються клітини видів роду С^їегіит. Вони зазвичай звужені до кінців, не-перешнуровані і мають серповидну форму: спинний край — більш-менш опуклий, а черевний — увігнутий або майже прямий. Проте яскрава симетрія напівклітин і в цьому випадку зберігається.
Розмноження вегетативне — поділом клітини по перешийку. Кожна напівклітина відновлює відсутню частину клітини (рис. 6.60). Статеве розмноження — кон'югація. Зигота проростає двома проростками.

Рис. 6.59. Десмідієві водорості: 1 - Micrasterias laticeps, 2 - Micrasterias truncata, 3 - Micrasterias radiosa, 4 - Micrasterias denticulate, 5 - Euastrum didelta, 6 - Cosmarium depressum, 7 - C. contractum, 8 - Micrasterias radiata (два екземпляри), 9 - Cosmarium pyramidatum, 10 - C. botrytis, 11 - Closterium littorale, 12 - Micrasterias arcuata
1
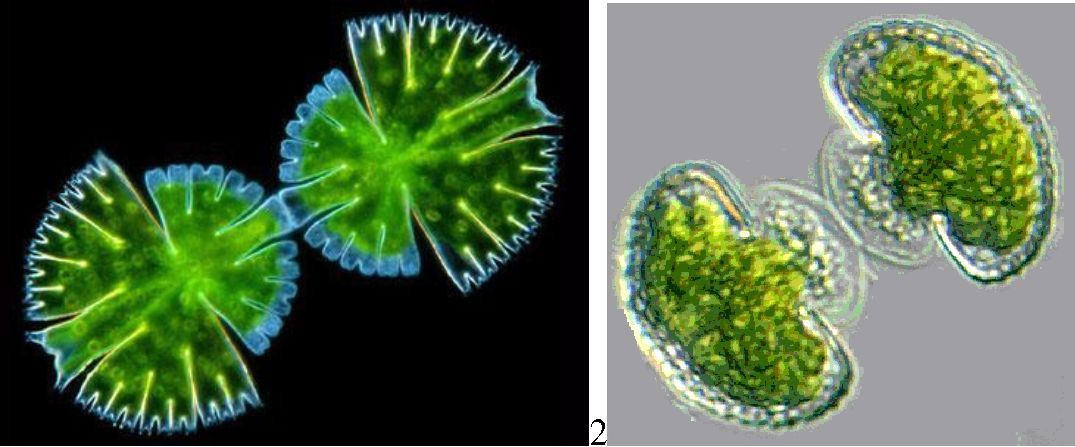
Рис. 6.60. Вегетативне розмноження у Micrasterias rotata (1) та y Cosmarium undulatum (2)
Харові водорості, харофіти, об'єднують близько 300 видів макроскопічних водоростей. Харові — найбільші з прісноводих водоростей. Довжина їх талому зазвичай складає 20-30 см, але може досягати і 1—2 м. Всі частини їх тіла, включаючи органи розмноження, добре помітні неозброєним оком. Талом зеленого кольору, безбарвні лише ризоїди.
Харові водорості мають складно і своєрідно побудоване тіло, не зовсім типове для нижчих рослин, членисто-кільчастої будови. За зовнішнім виглядом талому харові водорості нагадують вищі рослини, зокрема хвощі. Вони мають осьову частину, схожу на стебло і бічні вирости, подібні до розташованих у вигляді кілець листків. Місця розташування кілець листків називають вузлами, а частина стебла між ними — міжвузлям. Кожне міжвузля талому має довжину кілька сантиметрів і складається з однієї багатоядерної гігантської видовженої клітини, часто вкритої дрібними клітинами кори. Вузол складається з кількох дрібних одноядерних, зібраних у диск, клітин, які у процесі поділу диференціюються у бокові гілочки стебла, а також у кільця листків.
Оболонка клітин щільна, зсередини целюлозна, а ззовні утворена калозою, у якій може відкладатися вуглекисле вапно.
Всю центральну частину клітини займає центральна вакуоля з клітинним соком, а протоплазма розподілена по периферії клітини. У товстішому внутрішньому шарі протоплазми, що межую з вакуолею, розташовані численні ядра, в тонкому шарі, що прилягає до оболонки, зосереджені численні хлоропласти. Вони мають форму дрібних дисковидних тілець, подібних до хлорофілових зерен вищих рослин. Хлоропласти розподіляються рівномірно, забарвлюючи всю клітину в яскраво-зелений колір.
Для харових водоростей характерне статеве і нестатеве розмноження. Вегетативне розмноження відбувається шляхом вкорінення і подальшого розростання частин основних пагонів або за допомогою особливих одноклітинних або багатоклітинних бульб. Цій же меті служать особливі відростки, або вторинні пагони, що ростуть із старих листових вузлів і бульб. Статеве розмноження відбувається за допомогою статевих органів: антеридіїв і оогоніїв. Антеридії і оогонії утворюються з особливих клітин, що утворюються деякими периферичними клітинами листкових вузлів. Вони можуть виникати
або на одному таломі (однодомні види), або на різних таломах (дводомні види).
Оогоній має розміри близько 1 мм і складається з яйцеклітини і покриву, утвореного з п’яти вузьких спіральних клітин. Знизу оогоній має одноклітинну ніжку, а зверху - коронку з 5-10 коротких клітин.
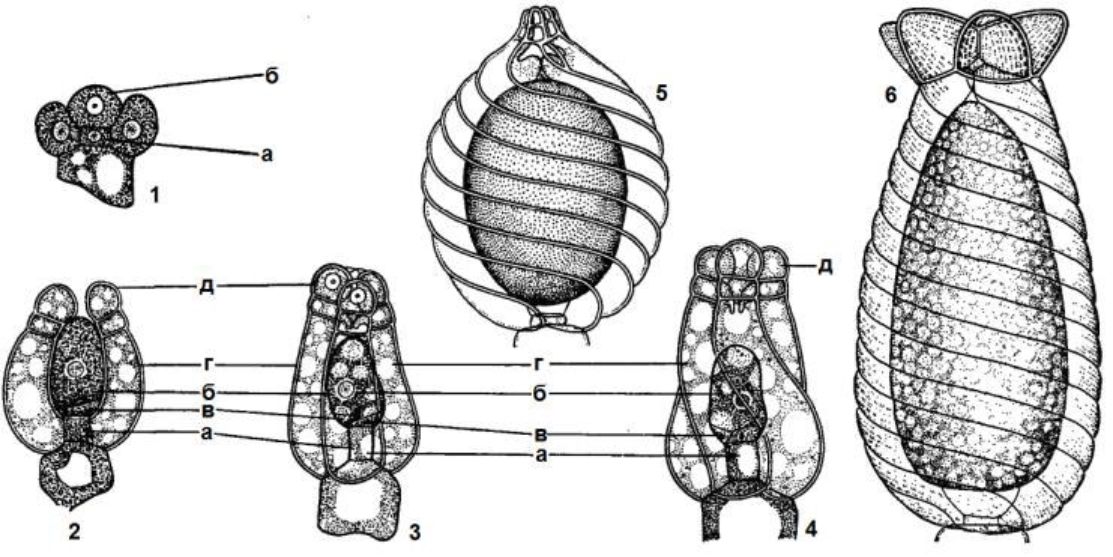
Рис. 6.61. Будова оогонію харових:
1-4 - послідовні стадії утворення оогонію (1 - початкова стадія, 2 - 4 - наступні стадії формування яйцеклітини і покривних спіральних клітин); 5 - зрілий оогоній нітели 6 - зрілий оогоній хари а - клітина листкового вузла, б - яйцеклітина, в - поворотні клітини, г - покривні клітини, д - клітини коронки
Антеридій має розміри близько 0,5 мм. Він кулястий, вкритий ззовні вісьмома плоскими клітинами, що зрослись своїми краями (рис. 6.62). В середину антеридію від кожної клітини відходять вирости з голівками, на яких утворюються антеридіальні нитки. Кожна з клітин ниток дає численні дводжгутикові сперматозоїди.
Антеридії та оогонії утворюються з клітин листкових вузлів (рис. 6.63).
Із заплідненої яйцеклітини розвивається ооспора, вкрита чотирма оболонками. Після періоду спокою ядро ооспори ділиться мейозом. Одна з гаплоїдних клітин ооспори дає початок новій рослині, а інші утворюють запас поживних речовин.
Живуть харові водорості переважно у прісних водоймах — ставках і озерах, але зустрічаються і в тимчасових невеликих водоймах: ямах, канавах, рідше в струмках і ріках. Є види, що населяють солонуватоводні водойми. У типово морських умовах харові не зустрічаються. У більшості випадків, вони утворюють густі зарості. Для багатьох харових необхідний підвищений вміст у воді вап
на. Харові водорості поширені по всій земній кулі, у водоймах всіх континентів, окрім Антарктиди і островів.
Рис. 6.62. Будова антеридію харових:
а - внутрішня базальна («пляшкоподібна») клітина, б - зовнішня базальна клітина, в - клітини листкового вузла, г - стінка, д - вирости, ж - головки, з - антеридіальні нитки
Рис. 6.63. Частина стебла хари (поздовжній зріз): а — клітини стеблової і листової кори; б — антеридії, в — оогонії
Походять харові водорості від високоорганізованих зелених водоростей з порядку хетофторових класу улотриксових.
Роль у природі харових водоростей порівняно невелика, проте там, де вони поселяються, виявляється їх вплив на гідрологічний режим і біологічні особливості водойм. їх жорсткі зарості є чудовим захистом для молоді риб і дрібних тварин. Відмерлі таломи утворюють лікувальні грязі. Зібрані водорості використовують як добриво, особливо на кислих грунтах, завдяки високому вмісту вапняку в їх таломах. У водоймах, де ростуть харові водорості, пригнічується розвиток личинок комарів. Досить великі ооспори харових поїдаються водоплавними птахами.
Відомо близько 300 видів сучасних харових, об'єднаних в 3 родини і 6 родів: Nitellaceae (2 роди), Nitellopsidaceae (1 рід) і Characeae (3 роди). На Україні до найбільш поширених видів харових відносяться Chara vulgaris, Ch. canescens Ch. tomentosa.
Водорості разом із вищими рослинами і автотрофними бактеріями складають ланку продуцентів у трофічних ланцюгах. Згідно оцінок різних учених внесок водоростей у загальну продукцію органічних речовин на планеті складає 26-90%.
У процесі фотосинтезу водорості продукують вільний кисень у водному середовищі, який йде на дихання водних тварин і рослин. Крім того, водорості відіграють важливу роль у загальному балансі кисню в атмосфері. Дуже багато кисню, який виділяється наземною рослинністю, витрачається у процесах окислення органічних сполук мікроорганізмами. У той же час, у водоймах розклад речовин проходить на дні в анаеробних умовах, а кисень виділяється у процесі фотосинтезу фітопланктоном з товщі води. Так відновлюється запас кисню в атмосфері. Океани служать головним регулятором балансу кисню атмосфери.
Значення водоростей в еволюції атмосфери і біосфери Землі. Найдревнішим виявленим залишкам фотоавтотрофних організмів приблизно 3,2 млрд. років. Вважається, що життя виникло на Землі приблизно 4 млрд. років тому. Тоді в атмосфері Землі, яка переважно складалася з водяної пари, вуглекислого газу, азоту, оксиду сірки, аміаку, метану і соляної кислоти, переважали відновлювальні процеси. Озоновий шар в атмосфері, який захищає все живе на планеті від згубної дії жорсткого ультрафіолетового випромінювання, тоді був відсутній.
Коли кількість кисню, який виділявся древніми синьозеленими водоростями, перевищила здатність водного середовища до його зв’язування, газ почав виділятися над поверхнею океану. Відновлювальні властивості атмосфери почали перетворюватися в окислювальні. Поступово в атмосфері сформувався шар озону. Це дало можливість живим організмам освоювати сушу.
Геологічне значення водоростей. Накопичуючи у своєму тілі карбонат кальцію разом з коралами брали участь у формуванні підводних скель і рифів, поширених у тропічній частині океанів (переважно червоні, деякі зелені і синьозелені водорості).
В утворенні крейдяних порід, крім форамініфер, брали участь золотисті водорості з вапняковою черепашкою. Це встановлено за допомогою електронного мікроскопа - крейда майже на 95% складається з коколітів - залишків панциря.
Водорості також були вихідним матеріалом для утворення рідких і твердих нафтоподібних сполук - сапропелів, горючих сланців, нафти.
Золотисті водорості накопичують у черепашках солі кальцію, діатомові - кремнезем.
Водорості у ґрунті сприяють збереженню родючості. Вони освоюють непридатні для життя субстрати: вулканічну лаву, туф, відвали гірських порід.
Отже, водорості відіграли і відіграють дуже важливу роль в розвитку життя на планеті. Однак вони зазнають зростаючого антропогенного впливу. Особливої охорони потребують водорості-макрофіти, які є об’єктом промислу. В деяких західноєвропейських країнах уже розроблені законодавство, яке обмежує промисел водоростей.
Охорона мікроводоростей повинна базуватися на стабілізації екологічних зв’язків у водних екосистемах. Через забруднення води окремі види водоростей можуть викликати «цвітіння» води.
Під «цвітінням» води розуміють інтенсивний розвиток водоростей у товщі води, у результаті чого вона набуває різного забарвлення. При цьому колір води змінюється від синьо-зеленого, яскраво-зеленого, сірого до жовто-зеленого, яскраво червоного, червоно-бурого і коричневого. Це залежить від забарвлення організмів, які викликають «цвітіння» і від їх чисельності.
Цвітіння води спостерігається і у континентальних водоймах (прісних і солоних), і в морях і океанах. В морях і океанах найчастіше «цвітіння» викликають діатомові і динофітові водорості, а також синьозелені. Для морів тропіків і субтропіків (прибережні частини Південної Америки і Африки, біля берегів Індії і Японії) характерні червоні припливи, обумовлені інтенсивним розвитком дино-фітових водоростей. Червоне море отримало свою назву через масовий розвиток синьозеленої водорості Oscillatoгia егЦйгаеа червоного кольору. В континентальних водоймах теж спостерігається червоне цвітіння.
У прісних водоймах літом «цвітіння» води найчастіше викликається синьозеленими і динофітовими водоростями, а весною і восени - діатомовими. Біомаса водоростей дніпровських водосховищ у період «цвітіння» досягає 0,5 кг/м3 і більше.
Вода набуває неприємного запаху, у ній з’являються токсичні
сполуки, проявляється дефіцит кисню, що може призвести літнього замору риб і до загибелі гідробіонтів. У людини вживання такої води може викликати гаффську хворобу.
Контрольні запитання та завдання
1. Описати особливості будови тіла водоростей.
2. У чому полягають особливості монадного типу структури водоростей? Серед яких водоростей він поширений?
3. У чому полягають особливості амебоїдного типу структури водоростей? Серед яких водоростей він поширений?
4. У чому полягають особливості гемімонадного типу структури водоростей? Серед яких водоростей він поширений?
5. У чому полягають особливості паренхіматозного типу структури водоростей? Серед яких водоростей він поширений?
6. У чому полягають особливості нитчастого і різнонитчастого типів структури водоростей?
7. У чому полягають особливості сифонального типу структури водоростей? Серед яких водоростей він поширений?
8. У чому полягають особливості сифоноклаального типу структури водоростей? Серед яких водоростей він поширений?
9. Які існують екологічні групи водоростей?
10. У чому полягають особливості водоростей водних місцезростань? Які пристосування до водного способу життя у водоростей планктону? У чому полягають особливості нейстонних водоростей?
11. У чому полягають особливості бентосних водоростей?
12. Які пристосування є у аерофільних водоростей?
13. У чому полягають особливості ґрунтових водоростей?
14. Як класифікують водорості? Назвіть відділи водоростей.
15. Описати особливості будови синьозелених водоростей.
16. Дати характеристику найважливіших представників синьозелених водоростей.
17. Які ціанеї викликають «цвітіння» води?
18. Яке значення синьозелених водоростей у природі і в житті людини?
19. Описати особливості будови евгленових водоростей.
20. Яке значення евгленових водоростей у природі і в житті людини? Назвати основних представників.
21. Описати особливості будови і розмноження динофітових водоростей.
22. Яке значення динофітових водоростей у природі і в житті людини?
23. Описати особливості будови і розмноження рафідофітових водоростей. Яке значення рафідофітових водоростей у природі і в житті людини?
24. Описати особливості будови і розмноження криптофітових водоростей. Яке значення криптофітових водоростей у природі і в житті людини?
25. Які особливості будови і розмноження золотистих водоростей?
26. Яке значення золотистих водоростей у природі і в житті людини? Назвати основних представників.
27. Які особливості будови і розмноження жовтозелених водоростей?
28. Яке значення жовтозелених водоростей у природі і в житті людини? Назвати основних представників.
29. Описати особливості будови і розмноження червоних водоростей.
30. Як класифікують червоні водорості?
31. Дати характеристику найважливіших представників червоних водоростей.
32. Яке значення червоних водоростей у природі і в житті людини?
33. Описати особливості будови і розмноження бурих водоростей.
34. Як класифікують бурі водорості?
35. Дати характеристику найважливіших представників бурих водоростей.
36. Яке значення бурих водоростей у природі і в житті людини?
37. Описати особливості будови і розмноження зелених водоростей.
38. Як класифікують зелені водорості? Описати особливості будови різних класів зелених водоростей.
39. Дати характеристику найважливіших представників зелених водоростей і їх значення у природі.
40. Описати особливості будови і розмноження харових водоростей.
41. Яке значення водоростей у природі і в житті людини?
42. Як виникає «цвітіння» води і у чому його небезпека?