СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ I. РАСТИТЕЛЬНАЯ КЛЕТКА
ГЛАВА 2. ЭУКАРИОТИЧЕСКАЯ КЛЕТКА
Заключение
Все живое состоит из клеток. Клетки крайне разнообразны по структуре и функциям. Одни представляют собой одноклеточные организмы, другие — высокоспециализированные — входят в состав многоклеточных организмов как животных, так и растений. И все же клетки очень сходны между собой. Все они окружены мембраной, называемой плазматической, или плазмалеммой. Плазматическая мембрана ограничивает цитоплазму клетки и ДНК. Все живое содержимое клетки называют протоплазмой.
Клетки подразделяются на две основные группы: прокариотические и эукариотические. Прокариотические (бактериальные) клетки лишены ядер и ограниченных мембранами органелл. В настоящее время к прокариотам относятся бактерии, в том числе и цианобактерии. Эукариотические клетки имеют настоящие ядра и отдельные компартменты — обособленные участки клетки, выполняющие различные функции.
Эукариотические клетки разделены на компартменты с помощью мембран, как правило трехслойных. Такие мембраны называются элементарными. Мембраны контролируют обмен веществ между клеткой и окружающей средой и между органеллами и цитоплазмой.
Растительные клетки обычно имеют полужесткую клеточную оболочку и протопласт — «единицу» протоплазмы, ограниченную клеточной оболочкой. Протопласт состоит из цитоплазмы и ядра. Цитоплазматический матрикс, или основное вещество, растительных клеток, как правило, находится в движении. Это явление известно, как ток цитоплазмы, или циклоз.Плазматическая мембрана, представляющая собой элементарную мембрану, отделяет цитоплазму от клеточной оболочки.
Помимо клеточной оболочки для растительных клеток характерны цитоплазматические вакуоли. Это полости, заполненные клеточным соком, водным раствором разнообразных солей, сахаров и других веществ. Вакуоли играют важную роль в увеличении размера клеток и поддержании упругости тканей. Кроме того, во многих вакуолях разрушаются макромолекулы, а продукты распада вовлекаются в новые метаболические процессы. Вакуоли окружены элементарной мембраной, называемой тонопластом. Молодые клетки обычно содержат многочисленные мелкие вакуоли, которые увеличиваются в размерах и сливаются в одну большую вакуоль, когда клетка достигает зрелости. Увеличение вакуолей приводит к росту клетки.
Ядро — контролирующий центр клетки — часто наиболее заметная структура протопласта. Оно окружено ядерной оболочкой, состоящей из двух элементарных мембран. В ядре находится хроматин, который в процессе клеточного деления конденсируется в отдельные хромосомы. Хроматин и хромосомы состоят из ДНК и белков.
Наряду с вакуолью и клеточной оболочкой характерным компонентом растительных клеток являются пластиды. Каждая пластида окружена оболочкой, состоящей из двух элементарных мембран. Классификация зрелых пластид основана на находящихся в них видах пигментов: хлоропласты содержат хлорофиллы и каротиноиды, хромопласты — каротиноиды, а в лейкопластах пигментов нет. Хлоропласты имеют граны — пачки уплощенных мембранных пузырьков, называемых тилакоидами, в которых сосредоточены пигменты. Различные виды пластид могут развиваться из мелких бесцветных телец — пропластид.
Подобно пластидам, митохондрии окружены двумя элементарными мембранами. Внутренняя мембрана изгибается и образует обширную мембранную систему; на складках внутренней мембраны локализованы ферменты. Митохондрии — это те органеллы эукариотических клеток, в которых происходит дыхание.
В дополнение к органеллам цитоплазма эукариотических клеток содержит две мембранные системы: эндоплазматический ретикулум и аппарат Гольджи. Эндоплазматический ретикулум — это обширная трехмерная система мембран. Она может связываться с ядерной оболочкой и служить местом прикрепления рибосом. Встречаются и свободные рибосомы (не связанные с эндоплазматическим ретикулумом). В эукариотических клетках они находятся как в цитоплазме, так и в ядре. В рибосомах аминокислоты соединяются друг с другом, образуя белки. В процессе синтеза белка отдельные рибосомы объединяются в полирибосомы, или полисомы.
Термином «аппарат Гольджи» обозначают все диктиосомы, или тельца Гольджи. Диктиосомы — это группы плоских, дископодобных цистерн, от которых отпочковываются многочисленные пузырьки. В диктиосомах, по-видимому, накапливаются и упаковываются сложные углеводы и другие вещества, которые в пузырьках переносятся к границе клетки. Пузырьки принимают участие и в образовании плазматической мембраны. Эндоплазматический ретикулум и диктиосомы объединены в единую эндомембранную систему.
Микротрубочки — тонкие структуры разной длины, состоящие из белка тубулина. Они играют немаловажную роль в митозе, формировании клеточной пластинки, построении клеточной оболочки и движении жгутиков.
Микрофиламенты — нити сократимого белка актина. Считается, что пучки этих тонких нитей играют решающую роль в циклозе. Вместе с микротрубочками микрофиламенты образуют цитоскелет клетки.
Жгутики (локомоторные органеллы) — это волосовидные структуры, отходящие от поверхности многих эукариотических клеток. Все жгутики эукариотических клеток имеют одинаковую систему организации 9 + 2, означающую, что наружное кольцо из девяти пар микротрубочек окружает две внутренние микротрубочки в центре жгутика.
Клеточная оболочка — это основной отличительный признак растительной клетки. Она определяет структуру клетки и текстуру растительных тканей. Клеточная оболочка обычно состоит из трех слоев: срединной пластинки, первичной и вторичной оболочек. Целлюлоза входит в состав клеточной оболочки в виде жестких фибрилл, состоящих из большого числа молекул целлюлозы. Целлюлозные фибриллы переплетены с нецеллюлозными молекулами матрикса — гемицеллюлоз и пектина. Во всех трех слоях клеточной оболочки может присутствовать и лигнин, особенно характерный для вторичной оболочки.
После отложения вторичной оболочки клетки, как правило, отмирают и формируют трубки для проведения воды или жесткие опорные структуры.
Взаимосвязь клеток растения достигается с помощью плазмодесм, которые пронизывают оболочки и связывают протопласты соседних клеток.
Делящиеся клетки проходят клеточный цикл, состоящий из интерфазы и митоза. Интерфазу можно разделить на три периода: период G1, в течение которого происходит общий рост и размножение органелл: период S, когда удваивается ДНК; период G2, когда синтезируются структуры, участвующие в митозе.
В интерфазе хромосомы находятся в неспирализованном состоянии и их трудно отличить от нуклеоплазмы. Во время митоза хромосомный материал — хроматин — конденсируется. и можно видеть, что каждая хромосома состоит из двух параллельных нитей — хроматид, соединенных центромерой. Центромера делится, и хроматиды, которые теперь называются дочерними хромосомами, увлекаются к противоположным полюсам. Деление клетки завершается образованием вокруг дочерних хромосом новых ядерных оболочек. Таким способом генетический материал поровну распределяется между двумя новыми ядрами.
Митоз, как правило, заканчивается цитокинезом —делением цитоплазмы. У растений и некоторых водорослей цитоплазма делится с помощью клеточной пластинки, которая начинает формироваться в телофазе митоза. После разделения цитоплазмы протопласты откладывают новые клеточные оболочки.
Приложение 1. Возникновение клеточной теории
В XVII в. английский врач Роберт Гук, используя микроскоп собственной конструкции, заметил, что пробка и другие растительные ткани состоят из маленьких ячеек, разделенных перегородками. Он назвал эти ячейки клетками. В современном значении это слово стали употреблять только 150 лет спустя.
В 1838 г. немецкий ботаник Маттиас Шлейден пришел к выводу, что все растительные ткани имеют клеточное строение. На следующий год зоолог Теодор Шванн подтвердил наблюдение Шлейдена на тканях животных и предположил, что клетка — основа жизни. Создание клеточной теории — значительный успех биологии, поскольку она подразумевает единство всех живых систем и объединяет различные направления биологии, изучающие разнообразные организмы.
В 1858 г. известный патолог Рудольф Вирхов сделал общее заключение, что клетки могут появляться только от других клеток: «Где существует клетка, там должна быть и предшествующая клетка, точно так, как животное происходит только от животного, а растение только от растения. ...Над всеми живыми формами, будь то организмы животных или растений, или их составные части, господствует вечный закон непрерывного развития».
Концепция Вирхова с точки зрения эволюции приобретает еще большую значимость. Существует непрерывная связь между современными клетками — и организмами, в состав которых они входят, — и примитивными клетками, которые впервые появились на Земле по крайней мере 3,5 млрд, лет назад.
Приложение 2. Мир микроскопии
Большинство клеток можно увидеть только с помощью микроскопа. При описании размеров клеток используют микрометры и нанометры (см. таблицу). Невооруженный человеческий глаз имеет разрешающую способность около 1/10 мм, или 100 мкм. Это означает, что, если вы смотрите на две линии, которые находятся друг от друга на расстоянии менее чем 100 мкм. они сливаются в одну. Точно так же две точки, лежащие на расстоянии менее 100 мкм, кажутся одной расплывчатой точкой. Чтобы различить структуры, расположенные более тесно, применяют оптические приборы, например, микроскопы. Лучший световой микроскоп имеет
разрешающую способность около 0,2 мкм, или 200 нм, т. е. примерно в 500 раз улучшает человеческий глаз. Теоретически построить световой микроскоп с большим разрешением невозможно.

Отметим, что разрешающая способность и увеличение не одно и то же. Если с помощью лучшего светового микроскопа вы получили фотографию двух линий, расположенных на расстоянии менее 0,2 мкм, то как бы вы ни увеличивали изображение, линии будут сливаться в одну. Используя более сильные линзы, можно получить большее увеличение, но не улучшить разрешение.
Трансмиссионный (просвечивающий) электронный микроскоп
Разрешающая способность электронного микроскопа почти в 400 раз больше светового. Это достигается за счет использования потока электронов вместо видимого света. Современные электронные микроскопы имеют разрешающую способность около 0,5 нм, примерно в 200 000 раз большую, чем человеческий глаз. (Диаметр атома водорода около 0,1 нм.)
В трансмиссионном (просвечивающем) микроскопе пучок электронов, проходя сквозь образец, оставляет его изображение на экране. Участки образца, пропускающие больше электронов, т. е. «электронопрозрачные», выглядят на экране светлыми, а «электроноплотные» — темными.
Просвечивающий электронный микроскоп имеет один крупный недостаток. Электроны обладают очень маленькой массой и должны двигаться в вакууме; при этом электронный пучок может пройти только сквозь очень тонкие образцы. Для их приготовления объект нужно зафиксировать и заключить в твердый материал, а затем по специальной методике приготовить тонкие срезы. Следовательно, высокая разрешающая способность электронного микроскопа может быть использована для изучения только фиксированных препаратов. Поэтому очень трудно идентифицировать изменения, произошедшие в препарате во время его приготовления.
Некоторые типы клеток, почти все вирусы и многие внутриклеточные структуры можно увидеть только с помощью трансмиссионного электронного микроскопа. Рис. 2-1 — одна из многих микрофотографий, помещенных в данной книге, которые получены с помощью трансмиссионного электронного микроскопа.
Сканирующий электронный микроскоп
В сканирующем (растровом) электронном микроскопе электроны, которые регистрируются и преобразуются в изображение, идут от поверхности образца. Электронный пучок фокусируется в тонком зонде и им сканируют образец. В результате этого образец испускает вторичные электроны слабой энергии. Различные участки поверхности испускают неодинаковое количество вторичных электронов. Меньшее количество испускают углубления и борозды и поэтому кажутся темными, большее — пики и выступы, которые выглядят светлыми. В результате получают трехмерное изображение. Электроны, отраженные поверхностью, и вторичные электроны собираются, усиливаются и передаются на экран. Рис. 2-10 и 2-16 так же, как многие другие в этой книге, получены с помощью сканирующего электронного микроскопа.
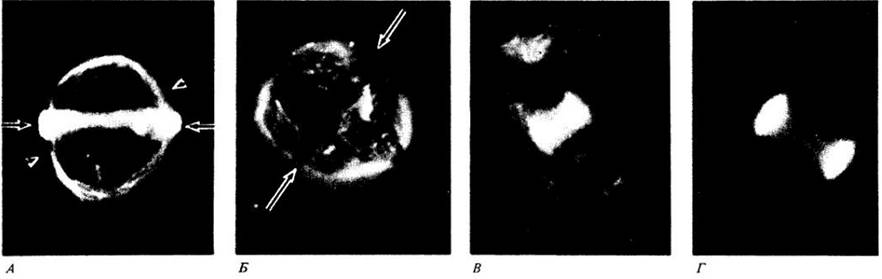
Приложение 3. Иммунофлуоресцентная микроскопия тубулина и микротрубочек
Иммунофлуоресцентная микроскопия позволила выяснить локализацию микротрубочек на разных стадиях деления клетки. В отличие от других методов она дает возможность получить трехмерное изображение. Здесь показаны клетки кончика корня лука (Allium сера), которые были окрашены специально приготовленными флуоресцентными антителами к тубулину и сфотографированы под флуоресцентным микроскопом. Флуоресцентные антитела, которые связались с белком тубулином, показали локализацию микротрубочек.
А. Препрофазный поясок микротрубочек (горизонтальные стрелки) окружает клетку перед профазой. Другие микротрубочки (наконечники стрелок) оконтуривают ядерную оболочку (не видна). Б. Препрофазный поясок исчез, и, место, которое он прежде занимал (отмечено стрелками), выглядит как нефлуоресцирующая полоса. Микротрубочки веретена распространились от этого района до полюсов веретена. В. В ранней телофазе, после того как хромосомы разделились, новые микротрубочки формируют фрагмопласт, в котором происходит образование клеточной пластинки. Г. По мере образования клеточной пластинки микротрубочки концентрируются там, где она распространяется от центра к стенкам делящейся клетки.