СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ II. ЭНЕРГИЯ И ЖИВЫЕ КЛЕТКИ
ГЛАВА 7. ФОТОСИНТЕЗ
Заключение
При фотосинтезе световая энергия превращется в химическую и углерод «фиксируется» в виде органических соединений.
Суммарное уравнение фотосинтеза следующее:
СО2 + 2Н2А + Световая энергия —> (СН2О) + Н2О + 2А,
где Н2А представляет воду или другое соединение, которое может быть окислено, т. е. может отдать электроны.
Первый этап фотосинтеза — это поглощение энергии света молекулами пигментов. Пигменты, участвующие в фотосинтезе эукариот, включают хлорофиллы и каротиноиды, которые упакованы в тилакоиды хлоропластов в виде фотосинтетических единиц, называемых фотосистемами. Свет, поглощенный молекулами пигментов, поднимает их электроны на более высокий энергетический уровень. Молекулы пигментов упакованы в фотосистемах таким бразом, что способны передавать эту энергию на специальную молекулу пигмента, называемую реакционным центром. Имеются две фотосистемы: фотосистема I и фотосистема II.
Не все фотосинтетические реакции требуют света. Совокупность реакций, которая требует света, относится к «световым реакциям», а та, которая не нуждается в свете, относится к «темновым реакциям».
Согласно общепринятой схеме световых реакций фотосинтеза, энергия света улавливается реакционным центром P680 фотосистемы II. Электроны от P680 поднимаются на более высокий уровень к электронному акцептору. Когда электроны удаляются, они замещаются электронами молекулы воды, и образуется кислород. Пары электронов затем «спускаются» по электронотранспортной цепи к фотосистеме I; при переносе электронов возникает протонный градиент, который способствует синтезу АТР из ADP и фосфата (фотофосфорилирование). Тем временем световая энергия, поглощенная фотосистемой I, поступает в ее реакционный центр Р700. Энергизованные электроны обязательно взаимодействуют с коферментом NADP, а в Р700 ушедшие электроны замещаются электронами из фотосистемы II. Энергия, выделяемая в световых реакциях, запасается в виде молекул NADPH2 и АТР, образуемого при фотофосфорилировании. Фотофосфорилирование происходит и при циклическом переносе электронов в обход фотосистемы II.
Подобно окислительному фосфорилированию, происходящему в митохондриях, фотофосфорилирование в хлоропластах — тоже хемиосмотический процесс. Когда электроны переносятся по электронотранспортной цепи от фотосистемы II к фотосистеме I, протоны выкачиваются из стромы в тилакоидное пространство, создавая градиент потенциальной энергии. Когда протоны преодолевают этот градиент, переходя из тилакоидного пространства обратно в строму, они идут через АТР-синтетазу и образуется АТР.
В темповых реакциях, которые происходят в строме хлоропласта, используется NADPH2 и АТР, образуемые в световых реакциях, чтобы восстановить двуокись углерода до органического углерода. Это осуществляется в цикле Кальвина. Молекула двуокиси углерода объединяется с исходным субстратом, пятиуглеродным сахаром — рибулозо-1,5-бисфосфатом (РБФ), и в результате образуется трехуглеродное соединение — 3-фосфоглицерат. При каждом обороте один атом углерода включается в цикл. Три оборота цикла дают трехуглеродную молекулу — глицеральдегид-3-фосфат, две молекулы которого (шесть оборотов цикла) могут объединяться с образованием молекулы глюкозы. При каждом обороте цикла происходит регенерация рибулозобисфосфата.
Растения, у которых цикл Кальвина — единственный путь фиксации углерода и 3-фосфоглицерат — первый стабильный продукт ассимиляции СО2, называются С3-растениями. У так называемых С4-растений двуокись углерода сначала фиксируется на фосфоенолпирувате с образованием оксалоацетата — четырехуглеродного соединения. Оксалоацетат затем быстро превращается либо в малат, либо в аспартат, которые переносят СО2 к РБФ цикла Кальвина. У С4-растений цикл Кальвина функционирует в клетках обкладок проводящих пучков, а С4-путь — в клетках мезофилла. С4-растения более экономно утилизируют СО2, чем С3-растения, частично благодаря тому, что фосфоснолпируваткарбоксилаза не ингибируется О2, и, таким образом, С4-растения обладают способностью поглощать СО2 с минимальной потерей воды. Кроме того, у С4-растений практически отсутствует фотодыхание — процесс выделения СО2 и поглощения О2 на свету.
Метаболизм по типу толстянковых (САМ) обнаружен у многих суккулентных растений. У CAM-растений фиксация СО2 фосфоенолпируваткарбоксилазой с образованием С4- сосдинений происходит ночью, когда открыты устьица. С4- соединения, запасенные в ночное время, затем в течение дня, когда устьица закрыты, отдают СО2 в цикл Кальвина на РБФ. У CAM-растений и цикл Кальвина, и С4-путь осуществляются в одной клетке; таким образом, оба данных пути, которые в С4-растениях пространственно разделены, у САМ- растений разделены во времени.
Приложение 1. Свет и жизнь
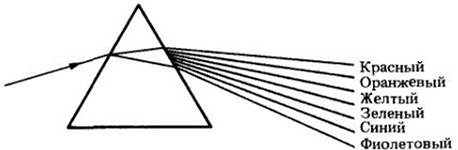
Почти 300 лет назад английский физик Исаак Ньютон (1642 — 1727) получил спектр видимого света, пропустив его луч через призму. С помощью этого эксперимента было показано, что белый свет состоит из целого ряда разных цветов — от фиолетового на одном конце спектра до красного на другом. Разделение их возможно благодаря тому, что лучи различного цвета, проходя через призму, преломляются под разными углами.
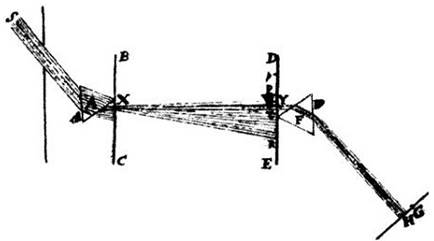
Рисунок к одной из статей Ньютона, иллюстрирующей его «ключевой эксперимент» по разложению «белого» цвета на составляющие. Солнечный свет проникает в темную комнату слева и, проходя через призму А, дает цветной спектр. Отверстие Y пропускает лучи только одного цвета. Проходя через вторую призму F, они преломляются, но не изменяют окраски. Таким образом Ньютон продемонстрировал, что сама призма не меняет цвет лучей, а просто по-разному преломляет лучи разного цвета
В XIX в. гениальный английский физик Джеймс Клерк Максвелл (1831 — 1879) установил, что видимый нами свет Y является лишь малой частью непрерывного спектра электромагнитных колебаний. Как показал Максвелл, вся образующая этот спектр радиация распространяется в виде волн. Их длины, т. е. расстояния от одного максимума волны до другого, варьируют в широких пределах — он нанометров у рентгеновских лучей до километров у низкочастотных радиоволн. Чем меньше длина волны, тем больше ее энергия. В видимом спектре у красного света длина волны максимальная, а у синего — минимальная. Общим свойством радиации является се способность распространяться в вакууме со скоростью 300 000 км/с.
К 1900 г. стало ясно, что волновая теория света не всегда адекватно объясняет экспериментальные факторы. Например, в 1888 г. был поставлен на первый взгляд довольно простой опыт. Оказалось, что цинковая пластинка, облученная ультрафиолетом, приобретает положительный заряд. Вскоре было установлено, что металл заряжается, так как радиация за счет своей энергии выбивает электроны из его атомов. Впоследствии обнаружилось, что этот фотоэлектрический эффект свойствен всем металлам, причем у каждого из них он проявляется при свойственной только данному элементу критической длине волны. Другими словами, необходимо облучение волнами определенной или более короткой (т. е. имеющей большую энергию) длины. Гипотеза, сформулированная Бором и другими физиками, о том, что электронные орбитали атомов имеют специфические энергетические уровни, основана на этих наблюдениях.
У некоторых элементов, например, натрия, калия и селена, эта критическая длина волны находится в пределах видимого спектра. Поэтому обычный свет может вызвать в них движение электронов (электрический ток). Работа электрических «глаз», открывающих, например, двери, а также фотоэкспонометров и телевизионных камер основан на этом принципе превращения световой энергии в электрическую.
Волна или частица?
Это довольно сложный вопрос. Волновая теория утверждает, что чем ярче свет, тем больше сила, «выбивающая» электроны из атомов. Однако важна длина волны, а не яркость. Даже эффект, тогда как более яркий, но с большей длиной волны его не дает. Более того, при увеличении яркости критического света возрастает число выбиваемых из металла электронов, но не скорость, с которой они покидают атомы. Чтобы увеличить ее, нужно использовать более коротковолновый свет. Кроме того, энергии нет необходимости накапливаться в металле. Даже слабый свет с критической длиной волны действует мгновенно.
Чтобы объяснить эти явления, Альберт Эйнштейн в 1905 г. сформулировал квантовую теорию света. Согласно ей, свет состоит из частиц энергии, называемых фотонами, или квантами света. Энергия фотона (или кванта) всегда обратно пропорциональна длине световой волны, т. е. чем длиннее волна, тем ниже се энергия. Фотоны фиолетового света, например, несут почти вдвое больше энергии, чем фотоны красного света, имеющего максимальную длину волны в видимой области.
Волновая теория позволяет физикам описывать некоторые свойства света одним математическим способом, а квантовая — другим. Обе эти модели не противоречат друг другу, а являются взаимодополняющими. Для более полного описания такого сложного явления, как свет, необходимо использовать их обе.
Сосуществование двух этих теорий иллюстрирует одну из тонкостей научного метода. Если ученый определяет и измеряет свет как волны, тот и проявляет волновые свойства. Если же использовать квантовый подход, то свет «выглядит» как поток частиц. Как сказал А. Эйнштейн: «Наши наблюдения зависят от теории, которой мы пользуемся».
Приспособленность к свету
Как показал Максвелл, видимый свет — лишь небольшая часть общего спектра радиации. С физической точки зрения разница между светом и темнотой, столь очевидная для человека, состоит только в нескольких нанометрах длины волны. «Качественных» границ у светового спектра нет. Почему же лишь малая часть электромагнитного излучения ответственна за зрение, фототропизм (стремление организмов к свету), фотопериодизм (сезонные изменения, которые происходят в существах в зависимости от длины дня и ночи), а также за процесс фотосинтеза, от которого зависит вся жизнь? По сути вся биологическая активность связана с
Белый свет, проходя через призму, разлагается на спектр различных цветов. Это обусловлено тем, что лучи каждого цвета слегка различаются по длине волны одними и теми же длинами волн. Что это, простое совпадение?

Один из выдающихся специалистов, изучающих процессы взаимосвязи света и живых организмов, — Джордж Уолд из Гарвардского университета отвечает на эти вопросы отрицательно. Он полагает, что жизнь, где бы она ни существовала, должна зависеть от одного и того же участка широкого спектар радиации. В основе его предположения лежат две гипотезы. Во-первых, живое состоит из громадных сложных молекул, которые имеют сложные взаимосвязи и специфические конфигурации, поддерживаемые в основном водородными или другими, еще более слабыми связями. Радиация с более мощной, чем у синего света, энергией может разорвать их, нарушив структуру и функцию этих молекул. При длине волны менее 200 нм она выбивает электроны из атомов, образуя ионы, поэтому называется ионизирующей. Энергия излучения с длиной волны больше, чем у видимого света, сильно поглощается водой, образующей большую часть массы живых организмов. Если свет с такой длиной волны и достигнет органических молекул, то сможет лишь увеличить их подвижность, но не изменит их структуру. Только излучение видимой части спектра способно возбуждать молекулы, т. е. другой, вызывая за счет этого изменения биологических систем.
Вторая идея заключается в том, что видимый свет в отличие от других участков спектра электромагнитной радиации был «выбран» организмами как наиболее доступный. Основная часть солнечного излучения, достигающая нашей планеты, лежит в пределах этой области. Имеющие более высокую энергию (т. е. более короткие) волны экранируются кислородом и озоном в высоких слоях атмосферы, а значительная часть инфракрасной радиации поглощается водяными парами и углекислотой, не успевая достичь земной поверхности.
Это можно назвать «приспособленностью окружающей среды»; пригодность условий для жизни и соответствие живых систем физическим параметрам среды — явления взаимосвязанные. Если бы такая взаимосвязь отсутствовала, жизнь была бы невозможной.
Приложение 2. Цикл углерода
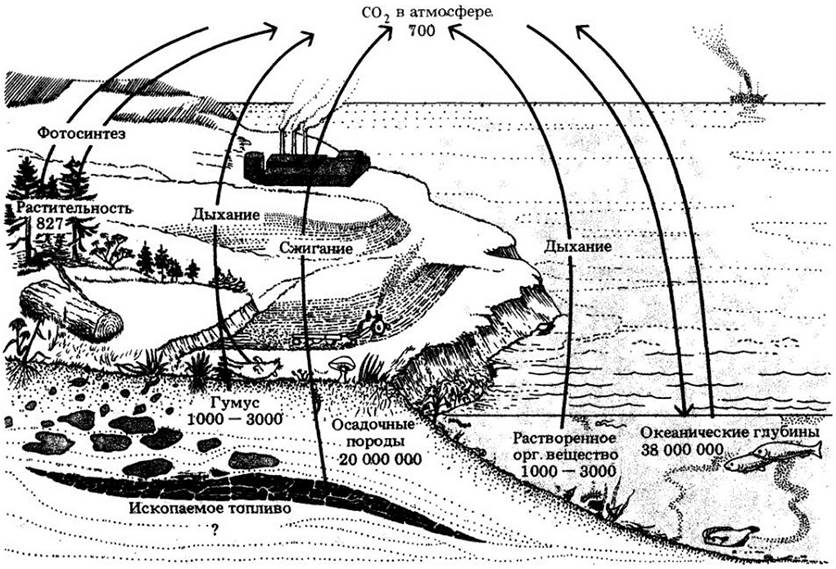
В процессе фотосинтеза живые системы поглощают углекислоту из атмосферы и включают ее в органические, углерод содержащие соединения. В процессе дыхания эти соединения распадаются вновь до СО2 и Н2О. Эти процессы в глобальном масштабе и образуют цикл углерода. Главными фотосинтезирующими компонентами в этом цикле являются растения и фитопланктон, морские водоросли и цианобактерии. Они синтезируют углеводы из углекислоты и воды и выделяют кислород в атмосферу. При фотосинтезе около 75 млрд, т углерода связывается в углеродсодержащие соединения за год.
Часть углеводов используется самими фотосинтезирующими организмами. Растения выделяют СО2 из корней и листьев, а морские водоросли и цианобактерии выделяют СО2 в воду, где поддерживается равновесие с углекислотой воздуха. Около 500 млрд, т углерода запасено в виде растворенной углекислоты в морях и 700 млрд. т — в атмосфере. Часть углеводов используется животными, которые питаются растениями, водорослями и другими организмами и выделяют при этом углекислоту. Громадное количество углерода содержится в отмерших остатках растений и других организмов. Кроме того, опавшие листья, раковины, фекалии и другие отбросы, которые накапливаются в почве или падают на дно океана, разлагаются редуцентами — небольшими беспозвоночными, бактериями и грибами. В результате этих процессов СО2 выделяется в воздух и воду. Далее, большой запас углерода лежит ниже поверхности почвы, в глубоких слоях земли в виде каменного угля и нефти, которые образовались много миллионов лет назад.
Естественные процессы фотосинтеза и дыхания сбалансированы между собой. В течение многих миллионов лет содержание углекислоты в атмосфере, насколько мы можем судить, оставалось постоянным. По объему это очень малая часть атмосферы — около 0,03%. Это очень важно, поскольку углекислота, как и другие компоненты атмосферы, поглощает тепло солнечных лучей.
Начиная с 1850 г. концентрация углекислоты в атмосфере начала расти, частично за счет использования ископаемого топлива, увеличения пахотных угодий, истребления лесов, особенно в тропиках. Некоторые экологи предсказывают, что увеличение углекислотного «покрова» увеличит температуру на Земле и соответственно приведет к расширению площадей, занятых пустынями. Другие, настроенные более оптимистично, предвидят повышение фотосинтетической активности растений и водорослей, связанное с увеличением количества углекислоты. Большинство, однако, испытывает тревогу в связи с тем фактом, что, хотя последствия нашей деятельности трудно предсказуемы, мы активно ее продолжаем.
Цикл углерода. Стрелками показано движение атомов С. Числа представляют собой оценки количеств запасенного углерода, выраженные, а миллиардах тонн. Количество углерода, выделяемое при дыхании и сжигании топлива, как считается, начало превосходить его количество, фиксируемое фотосинтетическим путем
Приложение 3. Изотопный состав углерода у С3- и С4-растений
В свободной атмосфере углекислота существует в форме 12СО2, ,13СО2 и 14СО2. В отличие от 14С формы 13С и 12С представляют собой стабильные изотопы. Растения ассимилируют их в неодинаковой степени. Сейчас установлено, что С3-растения в меньшей степени усваивают 13С, чем С4-растения. Этот «отсев» 13С осуществляется в основном на уровне реакции карбоксилирования с участием рибулозобисфосфат- карбоксилазы. Этот фермент у С3-растений сильнее «отсеивает» 13С, чем фосфоенолпируваткарбоксилаза С4-растений. В результате С4-растения содержат больше 13С, чем С3- растения.
Концентрация стабильных изотопов в растениях выражается величиной δ 13С относительно содержаний 12С и 13С для стандартного соединения (эта величина измеряется на масс-спектрометре). Значение δ 13С для С3-растений равно около -27‰ (по сравнению со стандартом), а у С4-растений — примерно -11‰. Показатель δ 13С используется для определения принадлежности растения к С3- или С4-типу.
Он важен и для экологов-физиологов. Поскольку в экосистеме растения служат пищей для растительноядных животных, анализ величины δ 13С в содержимом их желудка и фекалиях позволяет определить, какие растения — С3- или С4 — предпочитают употреблять в пищу те или иные растительноядные.