СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ III. ГЕНЕТИКА
ГЛАВА 8. ХИМИЯ И НАСЛЕДСТВЕННОСТЬ
Как работают гены?
Уотсон и Крик раскрыли химическую природу гена и предположили путь его редупликации. Однако один вопрос остался нерешенным: каким образом через образование специфических белков информация, содержащаяся в молекуле ДНК, влияет на структуру или функцию? Например, как ДНК «делает» вирулентным безвредный пневмококк, определяет форму листа или запах цветка, или почему ваши глаза такого же цвета, что и глаза вашей матери?
Молекулы наследственности
Структура и функция клеток почти целиком зависят от состава клеточных белков, в частности ферментов. Поэтому особый интерес представляют процессы образования специфических белков с использованием генетической информации ДНК, когда определенная линейная последовательность оснований «передается» соответствующей линейной последовательности аминокислот (первичной структуре белка, которая существенна для детерминации его формы и функции). На определенных этапах синтеза белка участвуют сходные с ДНК молекулы — молекулы рибонуклеиновой кислоты (РНК). Их участие предполагалось давно, поскольку клетки, синтезирующие большие количества белка, содержат и много РНК.
РНК несколько отличается от ДНК (см. гл. 3): сахарный компонент в молекуле РНК — это рибоза, а не дезоксирибоза, как в ДНК; кроме того, РНК содержит пиримидин урацил (U) вместо тимина ДНК. РНК редко имеет двухцепочечную структуру; таким образом, ее свойства и активность иные, чем у ДНК. Имеются три основных класса РНК: информационная, транспортная и рибосомальная.
Информационная (матричная) РНК (мРНК) — большая молекула величиной от нескольких сотен до 10 000 нуклеотидов. Она образуется на одной цепи ДНК по тому же принципу комплементарности, который используется и при синтезе дочерней цепи ДНК. Присутствие аденина в родительской цепи ДНК определяет присоединение урацила к образующейся цепи мРНК. Каждая последовательность трех оснований в молекуле мРНК во время трансляции (т. е. синтеза белка на матрице РНК) соответствует одной аминокислоте; такую последовательность называют кодоном.
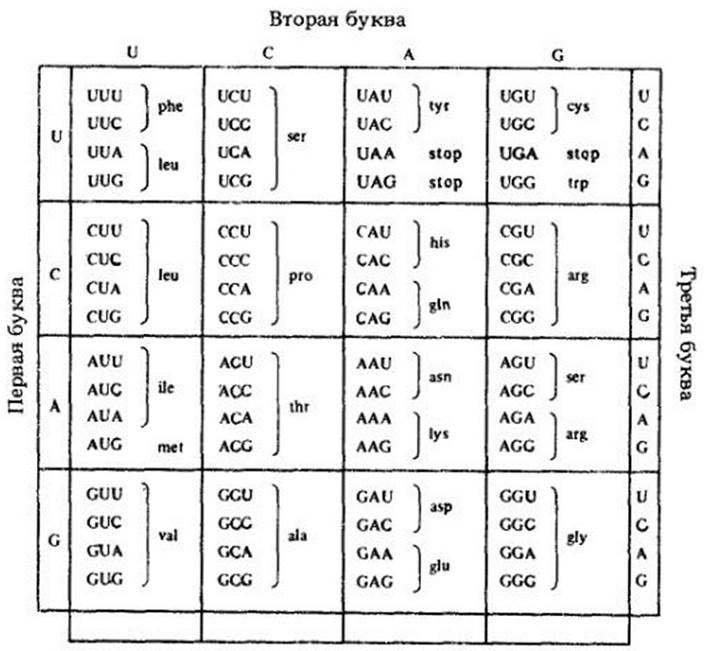
Транспортная РНК (тРНК). Ее иногда называют «словарем языка жизни». Существует несколько различных типов тРНК, по-видимому, по одному специфическому типу для каждого кодона генетического кода, т. е. кода, с помощью которого последовательность оснований в ДНК определяет последовательность аминокислот в образуемом белке (рис. 8-7). Генетический код одинаков у всех организмов, за исключением очень небольших деталей. Это одно из наиболее замечательных открытий в молекулярной биологии. Гены бактерий могут прекрасно работать (при надлежащих условиях) в клетках млекопитающих. Гены растений могут быть введены в бактерию, где будут направлять синтез своих собственных продуктов. Эти данные не только подтверждают общность происхождения всех живых существ, но и лежат в основе разработки методов генной инженерии, столь многообещающих для прогресса человечества (см. гл. 30).
Рис. 8-7. Генетический код, состоящий из 64 кодонов (триплеты оснований) и соответствующих им аминокислот (см. рис. 3-14). Из 64 кодонов только 61 используется для кодирования аминокислот. Остальные три кодона — это «стоп-сигналы» для окончания синтеза белка. Поскольку 61 триплет кодирует 20 аминокислот, то, очевидно, должны быть «синонимы», т. е. разные триплеты, кодирующие одну и ту же аминокислоту. Лейцин, например, кодируется шестью триплетами. Большинство «синонимов» отличается только по третьему нуклеотиду. Здесь показаны триплеты, кодирующие 20 аминокислот, и три «стоп-сигнала»
Каждая молекула тРНК состоит примерно из 80 нуклеотидов, которые соединены в одну длинную цепь, образующую петли (рис. 8-8). Отчасти специфичность отдельной тРНК определяется структурой се антикодона, или последовательностью трех оснований, благодаря которой она узнает определенный триплетный кодон на мРНК.
Рис. 8-8. А. Двухмерная структура молекулы тРНК, которая состоит примерно из 80 нуклеотидов, связанных в единую цепь. Цепь всегда заканчивается последовательностью ССА. Аминокислота присоединяется к специфической тРНК именно в этом месте. Некоторые нуклеотиды обнаружены у всех тРНК (выделены серым цветом). Другие варьируют от молекулы к молекуле. Буквами D, у,![]() и Т обозначены необычные модифицированные нуклеотиды, характерные для молекулы тРНК. Эти необычные нуклеотиды предотвращают образование некоторых водородных связей и способствуют установлению других, определяя тем самым характерную складчатую структуру молекулы. Каждый участок молекулы тРНК, по- видимому, имеет свою неповторимую функцию. К акцепторному концу присоединяется аминокислота. Левая петля, известная как петля
и Т обозначены необычные модифицированные нуклеотиды, характерные для молекулы тРНК. Эти необычные нуклеотиды предотвращают образование некоторых водородных связей и способствуют установлению других, определяя тем самым характерную складчатую структуру молекулы. Каждый участок молекулы тРНК, по- видимому, имеет свою неповторимую функцию. К акцепторному концу присоединяется аминокислота. Левая петля, известная как петля![]() имеет строение, одинаковое для всех молекул тРНК; она, вероятно, «управляет» связью тРНК с рибосомами. Правая петля, петля DНU, — напротив, неодинакова в различных молекулах тРНК; она, вероятно, участвует в выборе аминокислоты, присоединяемой с помощью фермента к акцепторному концу. Некоторые нуклеотиды соединены друг с другом водородными связями (показано цветными линиями). Непарные нуклеотиды третьей петли (цветной участок) названы антикодоном. Они служат для узнавания кодона мРНК. Б. Молекула образует складки, что создает трехмерную структуру. Это — фотография модели
имеет строение, одинаковое для всех молекул тРНК; она, вероятно, «управляет» связью тРНК с рибосомами. Правая петля, петля DНU, — напротив, неодинакова в различных молекулах тРНК; она, вероятно, участвует в выборе аминокислоты, присоединяемой с помощью фермента к акцепторному концу. Некоторые нуклеотиды соединены друг с другом водородными связями (показано цветными линиями). Непарные нуклеотиды третьей петли (цветной участок) названы антикодоном. Они служат для узнавания кодона мРНК. Б. Молекула образует складки, что создает трехмерную структуру. Это — фотография модели
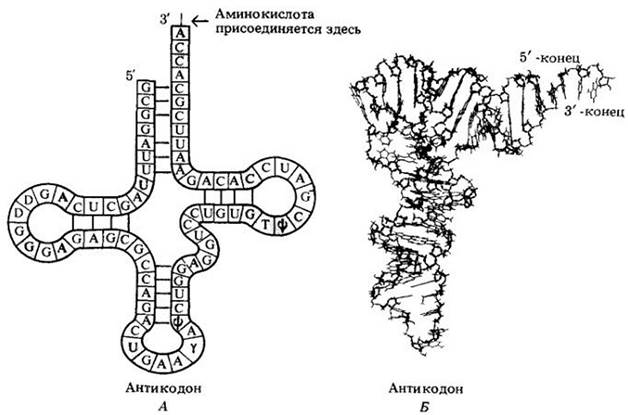
Установлены последовательности оснований многих типов тРНК. Эти последовательности различны, но все тРНК имеют сходное число оснований и одинаковую форму.
Специфичность тРНК определяется и ее способностью связываться с определенной аминокислотой — той, что соответствует антикодону. Эта способность зависит от
работы активирующих ферментов, которые узнают определенные аминокислоты и тРНК. Активирующие ферменты являются ключевыми элементами трансляции генетической информации; они связывают аминокислоту с соответствующей ей тРНК.
Рибосомальная РНК (рРНК) обнаружена в рибосомах вместе со специфическими белками. Рибосома состоит из двух субъединиц, каждая из которых построена из определенных РНК и белков. Например, у Escherichia coli малая субъединица рибосомы содержит один тип рРНК, а большая — два типа. мРНК и тРНК вместе подходят к рибосоме. Возможно, рибосомы определяют расположение мРНК, тРНК, аминокислот и строящихся белков относительно друг друга в процессе белкового синтеза. Белки рибосом тоже активны; один из них, например, относится к ферментам, образующим пептидные связи.
В клетках эукариот рРНК образуется на ДНК ядрышка. Хотя рибосомы этих клеток несколько крупнее, чем у Е. coli, они имеют сходное строение и выполняют те же функции, что и у бактерий.
Транскрипция РНК на ДНК
РНК транскрибируется, или копируется, с ДНК по принципу комплементарности, т. е. аналогично процессу репликации ДНК (рис. 8-9 и 8-10). В клетках эукариот обнаружены ферменты, называемые РНК-полимеразами, для транскрипции тРНК, рРНК и мРНК. Молекулы предшественника мРНК, синтезированные на матрице ДНК в ядрах эукариот, в 10 —20 раз крупнее, чем мРНК бактерий. Если РНК эукариот пометить с тем расчетом, чтобы радиоактивный урацил включался в образующуюся молекулу непродолжительное время, то длинные молекулы предшественника РНК в ядре обнаруживают радиоактивность. Однако только около 10%. этой радиоактивности оказывается вне ядра, подтверждая, что большая часть предшественника, синтезированного в ядре эукариот, не играет роли в образовании белков. С некодирующих областей генов эукариот — интронов — транскрибируются участки РНК, которые никогда не покидают ядра. Последовательности генов эукариот, кодирующие аминокислотную последовательность белка, называют экзонами.
Рис. 8-9. Ген бактерии в действии. Микрофотография (слева) показывает несколько цепей мРНК, образовавшихся одновременно. (На диаграмме справа цепи выделены цветом.) Самая длинная цепь начала синтезироваться первой. По мере того как каждая цепь отсоединяется от активного фрагмента хромосомы (молекулы ДНК), рибосомы прикрепляются к цепи мРНК и транслируют ее с образованием белка. Молекулы белка на микрофотографии не видны
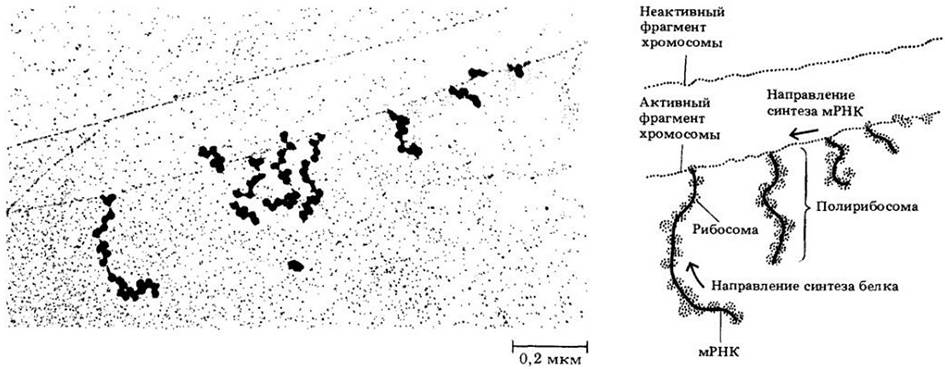
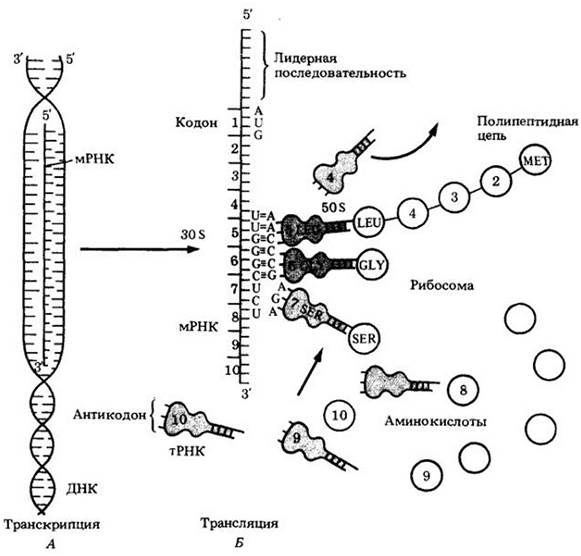
Рис. 8-10. Схематическое изображение процессов транскрипции и трансляции. А. В процессе транскрипции нить ДНК служит матрицей для синтеза комплементарной молекулы мРНК. Б. Во время трансляции РНК трех типов взаимодействуют со специфической группой ферментов и белков для образования новой полипептидной цепи. Рибосомальная РНК (рРНК) является компонентом рибосомы, которая осуществляет синтез белка. Рибосомы бактерий содержат большие (50S) и малые (30S) субъединицы. тРНК обеспечивает включение правильных аминокислот в растущую полипептидную цепь. Информационная РНК (мРНК) передает содержащуюся в гене информацию рибосоме. Информация закодирована в виде последовательности нуклеотидных триплетов, каждый из которых кодирует определенную аминокислоту. Каждый кодон узнается комплементарным антикодоном молекулы тРНК, которая ранее уже связалась с соответствующей аминокислотой. На рисунке большинство аминокислот показано кружками с цифрами; аминокислота глицин (GLY) только что«подвезена» к рибосоме, соответствующей транспортной РНК. Затем глицин соединится пептидной связью с лейцином (LEU), и растущая цепь удлинится на одну аминокислоту. После рибосома переместится на длину кодона мРНК и таким образом займет положение для связывания тРНК, несущей серин (SER)
Длинные молекулы предшественников мРНК эукариот после синтеза в ядре подвергаются «процессингу». Крупные фрагменты предшественника мРНК, транскрибированные с интронов, вырезаются из середины молекулы специальными ферментами. Остающиеся фрагменты, транскрибированные с экзонов, сшиваются вместе (сплайсинг) и образуют зрелую форму мРНК эукариот, которая выходит из ядра в цитоплазму. Зрелая мРНК эукариот имеет практически такую же величину, что и мРНК бактерий.
По-видимому, не только мРНК, но и рРНК, и тРНК первоначально транскрибируются с хромосом эукариот в виде крупных молекул-предшественников, которые затем подвергаются высокоспецифичному процессингу. Это наводит на мысль о том, что в генах эукариот имеются важные последовательности, которые не «имеют отношения» к функциональной РНК и синтезированному в итоге белку, но играют другую роль в процессе транскрипции генов.
Следовательно, мРНК эукариот, на которой синтезируется белок, в действительности сшита из фрагментов предшественника мРНК, который транскрибирован непосредственно с хромосомной ДНК. Таким образом, у бактерий и эукариот генетическая информация передается с гена на молекулу мРНК и затем на полипептидную цепь («центральная догма»1) генетики). Однако у эукариот процессинг добавляет еще один этап: ген —> первичный транскрипт —> мРНК —> полипептидная цепь.
1 В настоящее время популярное когда-то понятие «центральной догмы» можно считать устаревшим, поскольку оно подчеркивало направленность передачи информации с ДНК на РНК, между тем как в 1970 г. было открыто явление обратной транскрипции — синтеза ДНК на РНК с помощью фермента, имеющегося у некоторых вирусов. Таким образом, передача информации может идти как с ДНК на РНК, так и с РНК на ДНК. — Прим. перев.
Трансляция мРНК (синтез белка)
В ходе трансляции мРНК последовательность кодонов считывается в направлении 5' — 3', как и в процессе репликации ДНК (рис. 8-10). Начальный участок молекулы мРНК участвует в связывании рибосом. У бактерий малая субъединица рибосомы несет специальную тРНК (связывающую N-формилметионин) и проходит вдоль молекулы мРНК пока не «встретит» кодон AUG, соответствующий N-формилметио- нину; после этого малая субъединица связывает молекулу N- формилметионина в определенном участке рибосомы, образуя комплекс инициации. Эукариоты, напротив, не имеют инициирующего синтез белка кодона AUG; вместо него они «используют» химически модифицированную форму гуанина, а именно 7-метилгуанин.
Как у бактерий, так и у эукариот растущий конец белковой цепи присоединен молекулой тРНК к комплексу рибосома-мРНК (рис. 8-10). Следующая аминокислота, связанная с определенной молекулой тРНК, входит в комплекс и с помощью антикодона присоединяется к кодону (следующим трем основаниям мРНК). Аминогруппа входящей аминокислоты отдает рибосоме протон и образует пептидную связь с растущей белковой цепью. В итоге вся цепь теперь связана с вновь подошедшим комплексом тРНК-аминокислота, тогда как тРНК, с которой аминокислота только что была удалена, отделяется. Рибосома продолжает двигаться вдоль мРНК (шаг ее равен одному кодону) вместе с полипептидной цепью. При этом на каждом шаге добавляется еще одна аминокислота, и весь процесс повторяется.
Синтез поли пептидных цепей прекращается, когда рибосома «встретится» с одним из трех терминирующих кодонов: UAA, UAG или UGA. Эти так называемые нонсенс-кодоны не кодируют аминокислот; их присутствие приводит к расщеплению связи между последней аминокислотой белка и соответствующей тРНК. После этого синтез белка прекращается.