СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ I. РАСТИТЕЛЬНАЯ КЛЕТКА
ГЛАВА 2. ЭУКАРИОТИЧЕСКАЯ КЛЕТКА
Эндоплазматический ретикулум
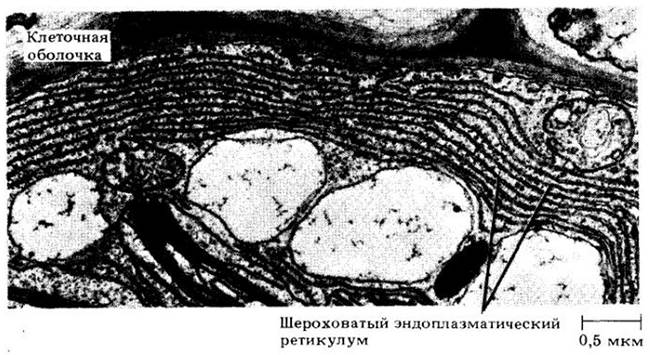
Эндоплазматический ретикулум — это сложная трехмерная мембранная система неопределенной протяженности. В разрезе эндоплазматический ретикулум выглядит как две элементарные мембраны с узким прозрачным пространством между ними (рис. 2-5, В и 2-20). Форма и протяженность эндоплазматического ретикулума зависят от типа клетки, ее метаболической активности и стадии дифференцировки. Например, в клетках, секретирующих или запасающих белки, эндоплазматический ретикулум имеет форму плоских мешочков, или цистерн, с многочисленными рибосомами, связанными с его внешней поверхностью. Эндоплазматический ретикулум, несущий рибосомы, называют шероховатым эндоплазматическим ретикулумом (рис. 2-5, В, 2-19 и 2-20). Полисомы и шероховатый эндоплазматический ретикулум — основные места синтеза белка. Напротив, клетки, которые секретируют липиды, имеют обширную систему трубочек, на которых нет рибосом. Эндоплазматический ретикулум, нс имеющий рибосом, называют гладкими эндоплазматическим ретикулумом. Гладкий эндоплазматический ретикулум обычно имеет трубчатую форму. Шероховатый и гладкий эндоплазматические ретикулумы могут присутствовать в одной и той же клетке. Как правило, между ними имеются многочисленные связи.
Рис. 2-20. Параллельные тяжи шероховатого эндоплазматического ретикулума (эндоплазматического ретикулума, усеянного рибосомами) видны на срезе клетки листа папоротника Vittaria guineensis. Относительно светлые участки — вакуоли
По-видимому, эндоплазматический ретикулум функционирует как коммуникационная система клетки. На некоторых электронных микрофотографиях можно видеть, что он связан с внешней оболочкой ядра. Фактически эти две структуры образуют единую мембранную систему. Когда ядерная оболочка во время деления клетки разрывается, ее обрывки напоминают фрагменты эндоплазматического ретикулума. Легко представить себе эндоплазматический ретикулум как систему транспортировки веществ, например, белков и липидов, в разные части клетки. Кроме того, эндоплазматические ретикулумы соседних клеток соединяются через цитоплазматические тяжи — плазмодесмы, — которые проходят сквозь клеточные оболочки (см. с.39).
Эндоплазматический ретикулум — основное место синтеза клеточных мембран. По-видимому, в некоторых растительных клетках здесь образуются мембраны вакуолей и микротелец, а также цистерны диктиосом.
Аппарат Гольджи
Термин аппарат Гольджи используется для обозначения всех диктиосом, или телец Гольджи, в клетке. Диктиосомы — это группы плоских, дисковидных пузырьков, или цистерн, которые по краям разветвляются в сложную систему трубочек (рис. 2-21). Диктиосомы в клетках высших растений обычно состоят из четырех — восьми цистерн, собранных вместе.
Рис. 2-21. Диктиосома состоит из группы плоских мембранных мешочков, связанных с пузырьками, которые, по-видимому, отпочковываются от мешочков. Диктиосома служит«центром упаковки» эукариотической клетки и играет ключевую роль в процессах секреции. А. Цистерны диктиосомы паренхимной клетки стебля хвоща Equisetum hyemale, видимые на срезе. Б. Одиночная цистерна, вид с поверхности. Стрелки указывают на многочисленные секреторные пузырьки вдоль краев цистерн
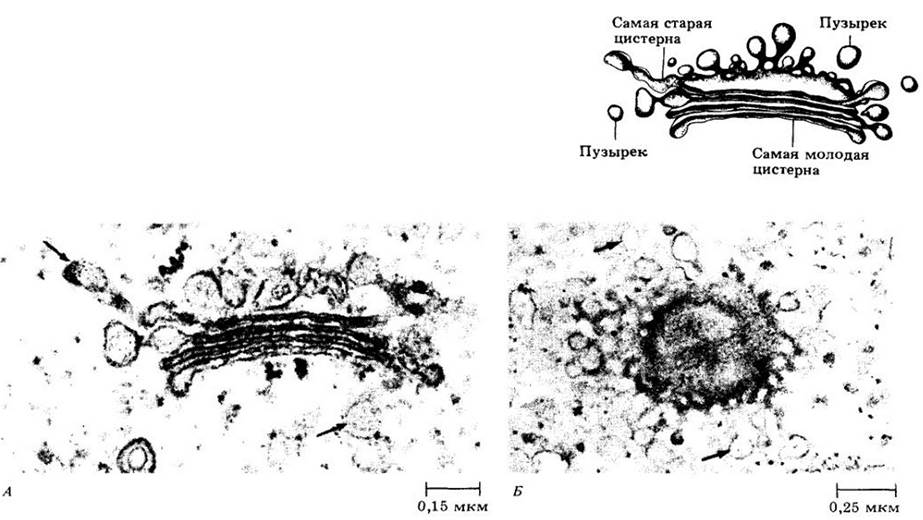
Обычно в пачке цистерн различают формирующуюся и созревающую стороны. Мембраны формирующихся цистерн по структуре напоминают мембраны эндоплазматического ретикулума, а мембраны созревающих цистерн — плазматическую мембрану (рис. 2-22).
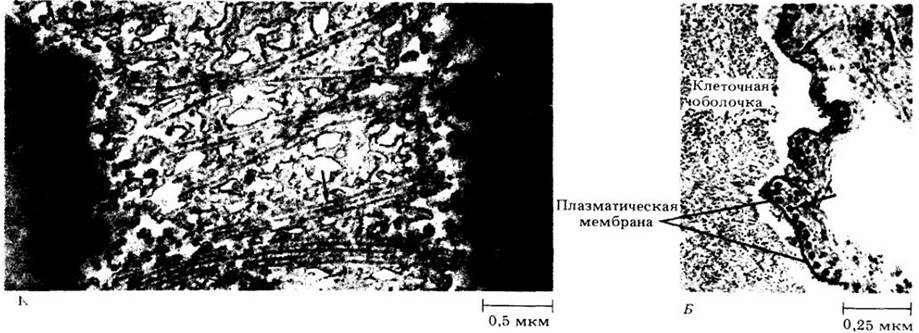
Диктиосомы участвуют в секреции, а у большинства высших растений — в образовании клеточных оболочек. Полисахариды клеточной оболочки, синтезируемые диктиосомами, накапливаются в пузырьках, которые затем отделяются от созревающих цистерн. Эти секреторные пузырьки мигрируют и сливаются с плазматической мембраной (рис. 2-22); при этом содержащиеся в них полисахариды встраиваются в клеточную оболочку. Продукты, накапливающиеся в диктиосомах, не всегда синтезируются ими. Некоторые вещества образуются в других структурах, например, в эндоплазматическом ретикулуме, а затем транспортируются в диктиосомы, где видоизменяются перед секрецией. Хороший пример подобных веществ — гликопротеины (углеводно- белковые соединения), важный строительный материал клеточной оболочки. Белковая часть синтезируется полисомами шероховатого эндоплазматического ретикулума, углеводная — в диктиосомах, где обе части объединяются, образуя гликопротсины.
Эндомембранная концепция
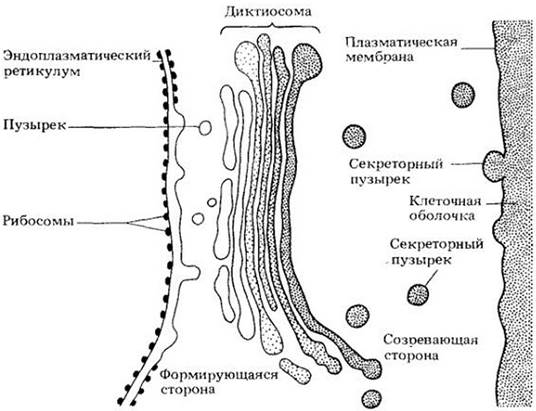
Мембраны — динамические, подвижные структуры, которые постоянно наменяют свою форму и площадь. На подвижности клеточных мембран основана концепция эндомембранной системы. Согласно этой концепции, внутренние мембраны цитоплазмы (за исключением мембран митохондрий и пластид) представляют собой единое целое и берут начало от эндоплазматического ретикулума. Новые цистерны диктиосом образуются из эндоплазматического ретикулума через стадию промежуточных пузырьков, а секреторные пузырьки, отделяющиеся от диктиосом, в конечном итоге способствуют формированию плазматической мембраны (рис. 2-22). Таким образом, эндоплазматический ретикулум и диктиосомы образуют функциональное целое, в котором диктиосомы играют роль промежуточных структур в процессе преобразования мембран, подобных эндоплазматическому ретикулуму, в мембраны, подобные плазматической.
Важно отметить, что даже в тканях, клетки которых слабо растут и делятся, постоянно происходит обновление мембранных компонентов.
Рис. 2-22. Схематическая иллюстрация эндомембран ной концепции. Новые мембраны синтезируются на шероховатом эндоплазматическом ретикулуме. Мелкие пузырьки отпочковываются от гладкой поверхности эндоплазматического ретикулума и переносят содержащиеся в них вещества и мембраны к формирующейся стороне диктиосомы. Секреторные пузырьки, отпочковывающиеся от цистерн созревающей стороны диктиосомы, мигрируют затем к плазматической мембране и сливаются с ней, добавляя новую порцию мембран и строительных материалов
Микротрубочки
Микротрубочки, обнаруженные практически во всех эукариотических клетках, — это тонкие цилиндрические структуры диаметром около 24 нм. Длина их варьирует. Каждая микротрубочка состоит из субъединиц белка, называемого тубулином. Субъединицы образуют 13 продольных нитей, окружающих центральную полость. Микротрубочки представляют собой динамические структуры, они регулярно разрушаются и образуются вновь на определенных стадиях клеточного цикла (см. рис. 2-15). Их сборка происходит в особых местах, которые называются центрами организации микротрубочек и в растительных клетках имеют слабо выраженную аморфную структуру.
У микротрубочек много функций. В растягивающихся и дифференцирующихся клетках микротрубочки, расположенные около внутренней поверхности плазматической мембраны, по-видимому, участвуют в образовании клеточной оболочки, контролируя упаковку целлюлозных микрофибрилл, которые откладываются цитоплазмой на растущую клеточную оболочку (рис. 2-23). Направление растяжения клетки определяется в свою очередь ориентацией целлюлозных микрофибрилл в клеточной оболочке. Микротрубочки направляют пузырьки диктиосом к формирующейся оболочке. подобно нитям веретена, которые образуются в делящейся клетке, и, по-видимому, играют роль в формировании клеточной пластинки (первоначальной границы между дочерними клетками). Кроме того, микротрубочки — важный компонент жгутиков и ресничек, в движении которых, по-видимому, играют немаловажную роль.
Рис. 2-23. Микротрубочки (отмечены стрелками), видимые на продольном (А) и поперечном (Б) срезах клеток листа папоротника Botrychium virginianum. А. Срез прошел более или менее параллельно клеточной оболочке у внутренней поверхности; видна плазматическая мембрана. Б. Можно видеть, что микротрубочки отделены от клеточной оболочки плазматической мембраной
Микрофиламенты
Микрофиламенты, подобно микротрубочкам, найдены практически во всех эукариотических клетках. Они представляют собой длинные нити толщиной 5 — 7 нм, состоящие из сократительного белка актина. Пучки микрофиламентов встречаются во многих клетках высших растений (рис. 2-24) и, по-видимому, играют определяющую роль в токах цитоплазмы. Микрофиламенты вместе с микротрубочками образуют гибкую сеть, называемую цитоскелетом.
Рис. 2-24. Пучок микрофиламентов в клетке листа папоротника олений рог (Platycerium bifurcatum)

Основное вещество
Еще недавно основное вещество клетки считали гомогенным богатым белком раствором с малым количеством структур или вообще беструктурным. Однако последние исследования клеток животных показывают, что основное вещество обладает сложной структурой. Под высоковольтным электронным микроскопом можно увидеть, что основное вещество представляет собой трехмерную решетку, построенную из тонких (диаметром 3 — 6 нм) тяжей, заполняющих всю клетку. Другие компоненты цитоплазмы, в том числе микротрубочки и микрофиламенты, подвешены к этой микротрабекулярной решетке.
Микротрабекулярная решетка делит клетку на две фазы: богатую белком (тяжи решетки) и богатую водой, заполняющую пространство между тяжами. Вместе с водой решетка имеет консистенцию геля — это настоящий живой гель.
Считается, что к микротрабекулярной решетке прикреплены органеллы, она осуществляет связь между отдельными частями клетки и направляет внутриклеточный транспорт. Есть данные, что и клетки растений имеют микротрабекулярную решетку.
Липидные капли
Липидные капли — структуры сферической формы, придающие гранулярность цитоплазме растительной клетки под световым микроскопом. На электронных микрофотографиях липидные капли выглядят аморфными (рис. 2-25). Похожие, но обычно более мелкие капли встречаются в пластидах (рис. 2-7, А).
Рис. 2-25. Компоненты цитоплазмы в паренхимной клетке утолщенного стебля, или клубнелуковицы, полушника (Isoeles muricata). По сторонам и ниже митохондрии, расположенной почти в центре электронной микрофотографии, можно видеть две липидные капли. Над митохондрией, несколько в стороне, находится цистерна эндоплазматического ретикулума, которая кажется раздувшейся. Некоторые вакуоли, возможно, произошли таким путем от эндоплазматического ретикулума. Часть вакуоли видна вверху микрофотографии. Плотный материал в вакуоли — таннин
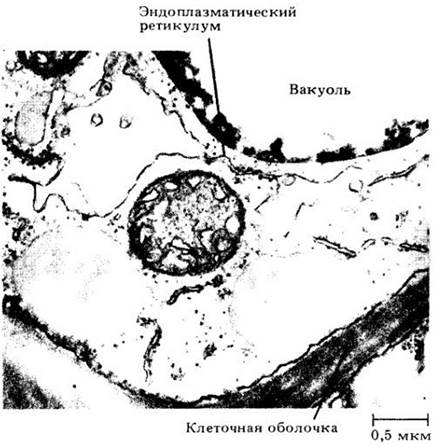
Липидные капли первоначально принимали за органеллы и называли сферосомами. Считалось, что они окружены двуслойной или однослойной мембраной. Однако последние данные показывают, что у липидных капель нет мембраны, но они могут быть покрыты белком.
Эрганические вещества
Эргастические вещества — это «пассивные» продукты протопласта: запасные вещества или отходы. Они могут появляться и исчезать в разные периоды клеточного цикла. Мы уже говорили о некоторых эргастических веществах: зернах крахмала, кристаллах, антоциановых пигментах и липидных каплях. Другими примерами подобных веществ могут служить смолы, камеди, таннины и белковые тела. Эргастические вещества входят в состав клеточной оболочки, основного вещества цитоплазмы и органелл, в том числе вакуолей.
Жгутики и реснички
Жгутики и реснички — это похожие на волоски структуры, которые отходят от поверхности многих эукариотических клеток. Они относительно тонки и имеют постоянный диаметр (около 0,2 мкм), но их длина колеблется от 2 до 150 мкм. Условно более длинные и немногочисленные из них называют жгутиками, а более короткие и многочисленные — ресничками. Четких различий между этими двумя типами структур тем не менее не существует, и мы будем использовать термин жгутик для обозначения обоих.
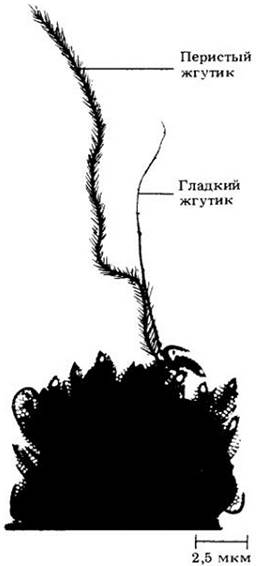
У некоторых водорослей и грибов жгутики являются локомоторными органами, с помощью которых организмы передвигаются в воде. У растений (например, мхов, печеночников, папоротников и некоторых голосеменных) только половые клетки (гаметы) имеют жгутики. Некоторые жгутики (перистые) несут один или два ряда мелких боковых отростков, другие (гладкие) лишены подобных образований (рис. 2-26).
Рис. 2-26. Два типа жгутиков — перистый и гладкий — обнаружены у одной клетки колониального организма Synura pe tersenii (золотистой водоросли). Перистый жгутик (слева) более длинный, чем гладкий (справа)
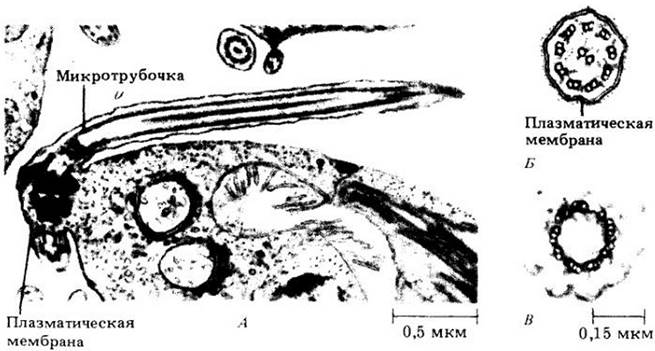
Внутреннее строение жгутиков удалось установить только с помощью электронного микроскопа. Каждый жгутик имеет определенную организацию (рис. 2-27). Наружное кольцо из девяти пар микротрубочек окружает две дополнительные микротрубочки, расположенные в центре жгутика. Содержащие ферменты «ручки» отходят от одной микротрубочки каждой из наружных пар. Эта основная схема организации 9+2 обнаружена во всех жгутиках эукариотических организмов.
Движение жгутиков и ресничек осуществляется автономно; жгутики и реснички способны двигаться и после отделения от клеток. Некоторые исследователи считают, что движение жгутиков основано на скольжении микротрубочек, при этом наружные пары микротрубочек движутся одна вдоль другой без сокращения. Скольжение пар микротрубочек относительно друг друга вызывает локальное изгибание жгутика. По-видимому, скольжение происходит в результате контакта «ручек» одной пары микротрубочек с микротрубочками соседней пары.
Жгутики «вырастают» из цитоплазматических цилиндрических структур, называемых базальными тельцами, образующими и базальную часть жгутика (рис. 2-27). Базальные тельца имеют внутреннее строение, напоминающее строение жгутика, за исключением того, что наружные трубочки в базальном тельце собраны в тройки, а не в пары и две центральные трубочки отсутствуют.
Рис. 2-27. А. Продольный срез жгутика гаметы зеленой водоросли Vivaria. Обратите внимание на то, что мембрана, окружающая собственно жгутик, представляет собой единое целое с плазматической мембраной. Б. Поперечный срез через жгутик Ulvaria. Видна типичная структура 9+2. В. Поперечный срез через базальное тельце жгутика Vivaria. Обратите внимание на то, что базальное тельце имеет кольцо из 9 триплетов микротрубочек и не имеет центральных микротрубочек