СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ V. СТРОЕНИЕ И РАЗВИТИЕ ТЕЛА ПОКРЫТОСЕМЕННЫХ
ГЛАВА 20. КЛЕТКИ И ТКАНИ РАСТЕНИЯ
По мере развития зародыша образование новых клеток, тканей и органов почти полностью ограничивается меристемами, сохраняющими физиологическую молодость тканями, которые имеют основное значение для роста. Как отмечалось в гл. 1, существуют два их главных типа — апикальные и латеральные. Апикальные меристемы обеспечивают главным образом нарастание в длину. Они располагаются на концах (апексах) побегов и корней. Рост, в процессе которого формируются первичные ткани, называют первичным. Говорят, что часть растения, состоящая из этих тканей, — это его первичное тело.
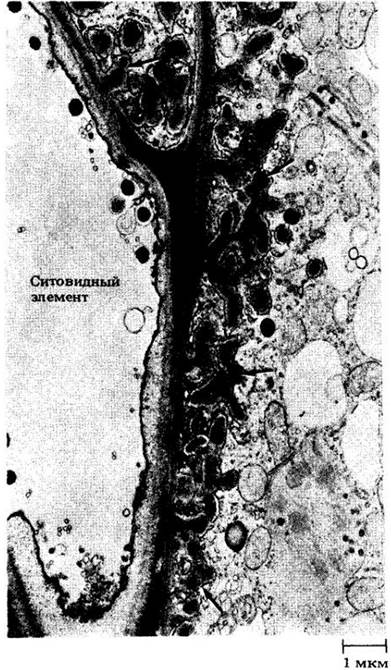
Рис. 20-1. Поперечный срез флоэмы папоротника Platycerium bifurcatum с участками двух передаточных клеток, оболочка которых имеет многочисленные впячивания (см. стрелки). Эти клетки примыкают к более светлому на снимке ситовидному элементу, клетке, проводящей питательные вещества
Латеральные меристемы — сосудистый1 и пробковый камбий — дают вторичные ткани, образующие вторичное тело растения. Камбий дает вторичные ксилему и флоэму, а пробковый камбий главным образом пробку.
1 Сосудистый камбий обычно называют просто камбием. — Прим. ред.
И в апикальных, и в латеральных меристемах некоторые клетки непрерывно делятся. После каждого деления одна из сестринских клеток остается в меристеме, а другая включается в неделящиеся ткани. Первые клетки, продолжающие делиться, называют инициалями, а вторые — производными. Важно отметить, что производные обычно делятся один или более раз, прежде чем начинают дифференцироваться в специализированные типы клеток. Следовательно, меристема включает инициали и их непосредственные производные.
Клеточные деления происходят не только в апикальных и латеральных меристемах. Например, протодерму, прокамбий и основную меристему, представляющие собой частично дифференцированные ткани, называют первичными меристемами, так как (1) они дают начало первичным тканям и (2) многие их клетки остаются некоторое время меристематическими и лишь затем дифференцируются. Первичные меристемы, за исключением возникающих в процессе эмбриогенеза в собственно зародыше, образуются из апикальных.
Рост растения — результат деления и растяжения клеток. С возрастом меристематических тканей общий размер их клеток увеличивается; в конце дифференцировки главным способом роста молодых участков корня, стебля или листа становится растяжение клеток.
Дифференцировка — процесс приобретения клетками с одинаковым генотипом индивидуальных отличий, в том числе и от меристематической клетки, давшей им начало. Часто он начинается еще во время их растяжения. Созрев, т. е. завершив дифференцировку, некоторые клетки остаются живыми, а другие отмирают. Среди тех и других различают множество типов. Вопрос о том, каким образом клетки с одинаковым происхождением становятся такими разными, — один из актуальнейших в современной биологии. Некоторые факторы, участвующие в управлении клеточной дифференцировкой, обсуждаются в последних разделах гл. 8 и в гл. 24 (с. 101).
Основной порядок размещения тканей устанавливается на ранних этапах меристематической деятельности. Их организация и форма растения определяются как делением, так и растяжением клеток. Приобретение свойственной тому или другому виду формы называется морфогенезом (от morphé — форма и genere — создавать).
Системы тканей
Ботаники уже давно выяснили, что важнейшие ткани сосудистого растения объединены в более крупные комплексы. Они называются системами тканей, и их присутствие в корне, стебле и листе отражает фундаментальное сходство этих органов и единство тела растения. Таких систем по меньшей мере три: (1) система основных тканей, (2) система проводящих тканей, (3) система покровных тканей. Как отмечалось в гл. 19, они закладываются в процессе эмбриогенеза и представлены на этом этапе первичными меристемами — соответственно основной меристемой, прокамбием и протодермой.
Система основных тканей состоит из паренхимы (наиболее распространенная ткань), колленхимы и склеренхимы. Система проводящих тканей представлена ксилемой и флоэмой. Эпидерма, наружная кожица первичного тела растения, и перидерма, сменяющая ее вторичная покровная ткань, составляют систему покровных тканей.
Ткани и их клетки
Ткани можно определить, как структурно и (или) функционально обособленные группы клеток. Если они состоят только из одного типа клеток, их называют простыми, а если включают два или более их типа — сложными. Паренхима, колленхима и склеренхима — простые ткани; ксилема, флоэма и эпидерма — сложные.
Паренхима
Паренхима, предшественница всех других тканей, состоит из паренхимных клеток. В первичном теле растения они обычно встречаются в виде сплошных масс в коре стеблей (рис. 20-2) и корней, сердцевине стеблей, мезофилле листьев (см. рис. 20-23) и мякоти плодов. Кроме того, эти клетки образуют вертикальные тяжи в первичной и вторичной проводящих тканях и горизонтальные тяжи (лучи) во вторичных проводящих тканях (см. гл. 23).
Рис. 20-2. Поперечный срез коры стебля бузины канадской (Sambucus canadensis). Видны клетки колленхимы с неравномерно утолщенными оболочками (вверху) и паренхимные клетки (внизу). Светлые пространства между ними — межклетники. У некоторых паренхимных клеток на фронтальных клеточных стенках видна сетчатая структура. Ее светлые (тонкие) участки — первичные поровые поля

Обычно живые в зрелом состоянии, паренхимные клетки способны к делению, и, хотя обычно сохраняют первичные оболочки, иногда приобретают и вторичные. Благодаря способности к делению паренхимные клетки с первичными оболочками играют важную роль в регенерации и заживлении ран. Именно они дают начало придаточным корням на стеблевых черенках. Эти клетки участвуют в фотосинтезе, запасании веществ и секреции, т. е. процессах, протекающих с участием живой протоплазмы. Кроме того, они могут играть определенную роль в передвижении воды и транспорте питательных веществ в растении.
Передаточные клетки
В течение последних лет значительное внимание привлек особый тип паренхимных клеток с характерными впячиваниями клеточных оболочек, часто сильно увеличивающими поверхность плазматической мембраны (см. рис. 20-1). Предполагают, что такие клетки, названные передаточными, играют важную роль в переносе растворенных веществ на короткие расстояния. Хотя о существовании их в различных органах знали давно, лишь недавно выяснилось, что они широко распространены по всему растению и, вероятно, повсюду выполняют сходные функции. Передаточные клетки связаны с ксилемой и флоэмой мелких жилок семядолей и листьев многих травянистых двудольных, а также с флоэмой и ксилемой листовых следов в узлах двудольных и однодольных; кроме того, их находят в различных тканях репродуктивных (плацента, зародышевый мешок, эндосперм) и железистых (нектарники, солевые железки, железки плотоядных растений) структур. Каждая из этих зон является потенциальным местом интенсивного транспорта растворенных веществ на короткие расстояния.
Колленхима
Колленхима состоит из колленхимных клеток, которые, как и паренхимные, остаются живыми в зрелом состоянии (рис. 20-2 — 20-5). Обычно она располагается отдельными тяжами или непрерывным цилиндром под эпидермой стебля и черешков листьев, а также окаймляет жилки в листьях двудольных (из нее почти целиком состоят выступы на наружной поверхности черешков и жилок листа сельдерея). В типичном случае вытянутые колленхимные клетки (рис. 20- 3) имеют неравномерно утолщенные нелигнифицированные первичные оболочки, делающие их особенно хорошо приспособленными для укрепления молодых растущих органов (см. описание первичной оболочки в гл. 2). Само название колленхимапроисходит от греческого слова colla — «клей» и возникло в связи с этими характерно утолщенными, блестящими в свежих тканях оболочками (рис. 20-5). Будучи первичными, они легко растяжимы и практически не мешают удлинению той части растения, в которой находятся. Кроме того, поскольку зрелые колленхимные клетки остаются живыми, у них может продолжаться образование толстых гибких оболочек параллельно росту органа в длину.
Рис. 20-3. Продольный срез, показывающий вытянутые клетки колленхимы в стебле тыквы (Cucurbita maxima). Трихом — придаток эпидермы (на рис. 20-22 показано несколько их типов)
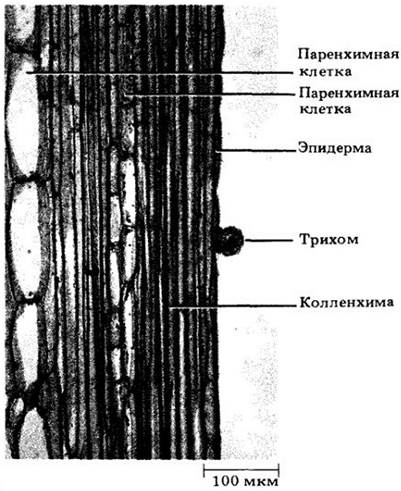
Рис. 20-4. Электронная микрофотография поперечного среза зрелой клетки колленхимы из тычиночной нити пшеницы (Triticum aestivum). Можно видеть протоплазматическое содержимое и неравномерно утолщенную оболочку
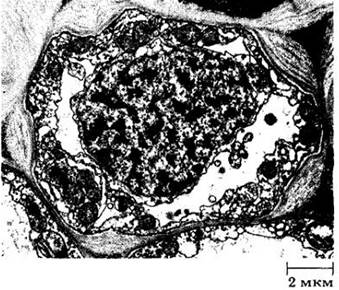
Рис. 20-5. Поперечный срез колленхимы из черешка листа ревеня (Rheum rhaponticum). На свежей ткани (как на этом снимке) неравномерно утолщенные оболочки колленхимных клеток выглядят блестящими

Склеренхима
Склеренхима состоит из склеренхимных клеток, которые могут развиваться в любых или даже во всех частях первичного и вторичного тела растения; протопласты у них в зрелом состоянии часто отсутствуют. Термин происходит от греческого skleros, что означает «твердый», поскольку главный признак склеренхимных клеток — толстые, часто лигнифицированные вторичные оболочки. Благодаря им эти клетки являются важными укрепляющими и опорными элементами завершивших рост в длину частей растения (см. описание вторичных клеточных оболочек в гл. 2).
Различают два типа склеренхимных клеток: волокна и склереиды. Волокна — как правило, длинные тонкие клетки, обычно собранные в тяжи или пучки (рис. 20-6). Экономически важные так называемые лубяные волокна, например, пеньковое, джутовое и льняное, получают из стеблей двудольных или (в частности, манильская пенька) из листьев однодольных. Склереиды разнообразны по форме и часто разветвлены (рис. 20-7), но по сравнению с большинством волокон являются относительно короткими клетками. Они могут встречаться изолированно или группами по всей основной ткани. Ими образована семенная кожура, скорлупа орехов, косточка, или эндокарпий, косточковых плодов; они придают мякоти груш характерный крупитчатый характер (рис. 20-8).
Рис. 20-6. Волокна первичной флоэмы из стебля липы (Tilia americana) на поперечном (А) и продольном (В) срезах. Во вторичных оболочках этих длинных толстостенных волокон — относительно незаметные поры. На рис. Б можно видеть только часть их длины
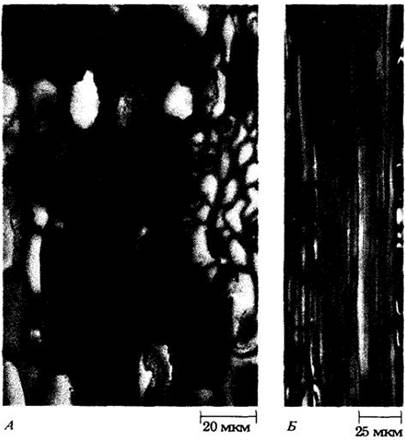
Рис. 20-7. Ветвистая склереида из листа кувшинки (Nymphéa odorata) в обычном (А) и поляризованном (Б) свете. Ее оболочка содержит многочисленные мелкие угловатые кристаллы
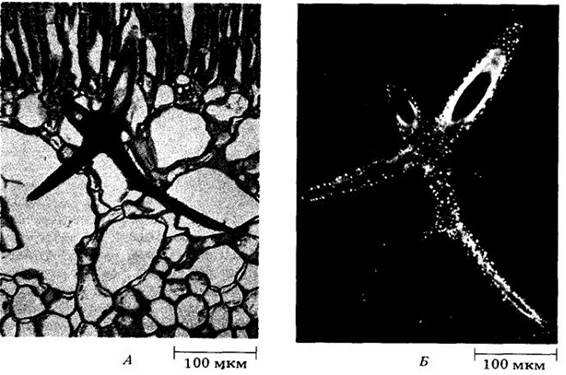
Рис. 20-8. Склереиды (каменистые клетки) из мякоти груши (Руrus communis). Во вторичных оболочках — отчетливые простые поры с многочисленными ответвлениями. Такие поры называют ветвистыми (рамифицированньми). В процессе формирования групп каменистых клеток в мякоти плода груши меточные деления происходят концентрически вокруг некоторых склереид, образовавшихся раньше прочих. Новые клетки дифференцируются в каменистые, увеличивая эти группы

Ксилема
Ксилема — главная водопроводящая ткань сосудистых растений. Она также участвует в транспорте минеральных веществ и запасании питательных соединений, а также выполняет опорную функцию. Вместе с флоэмой ксилема образует непрерывную систему проводящих тканей, пронизывающую все тело растения (рис. 20-9). По происхождению она может быть первичной и вторичной. Первичная ксилема формируется прокамбием, а вторичная — камбием (см. гл. 23).
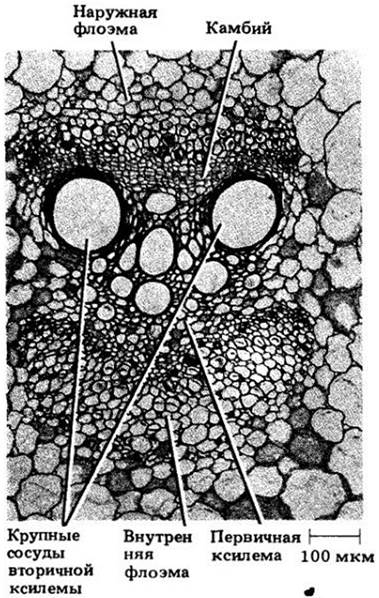
Рис. 20-9. Поперечный срез проводящего пучка из стебля тыквы (Cucurbita maxima). Флоэма располагается по обе стороны ксилемы, между наружной флоэмой и ксилемой развивается камбий
Главные проводящие клетки ксилемы — трахеальные элементы двух типов — трахеиды и членики (элементы) сосудов. И те, и другие вытянуты, в зрелом состоянии лишены протопластов и несут на своих вторичных оболочках поры (рис. 20-10, А — Г). Кроме того, членики сосудов имеют перфорации, т. е. участки без первичной и вторичной стенок, фактически отверстия в стенке. Они присутствуют главным образом на концах члеников, но могут находиться и на боковых поверхностях. Перфорированная часть стенки называется перфорационной пластинкой (рис. 20-11).
Рис. 20-10. Типы клеток вторичной ксилемы, или древесины, дуба (Quercus), выделенные из неокрашенной мацерированной (т. е. разделенной на отдельные клетки) ткани. А, В — широкие членики сосудов. В — узкий членик сосуда. Г — трахеида. Д — волокнистая трахеида. Е — волокно либриформа (самый длинный тип волокон в древесине дуба). Пятнистый вид этих клеток объясняется порами в их оболочках (кроме Е)

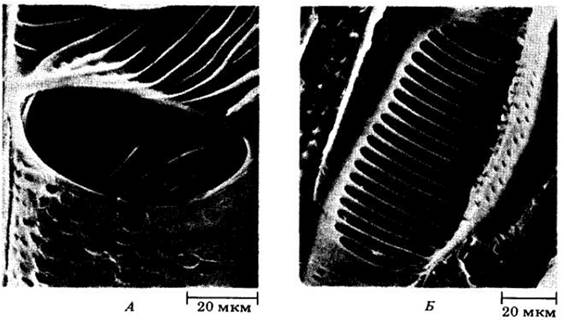
Рис. 20-11. Фотография в сканирующем электронном микроскопе перфорированных концевых стенок члеников сосудов вторичной ксилемы. А. Простая перфорационная пластинка с единственным крупным отверстием между двумя члениками сосудов у липы (Tilia americana). Б. Лестничная перфорационная пластинка между члениками сосуда ольхи красной (Alnus rubra). На стенках около перфорационных пластинок можно видеть поры
Членики сосудов располагаются один за другим в виде длинных непрерывных трубок, называемых сосудами (рис. 20-12). Поры длинных, имеющих скошенные концы трахеид концентрируются на примыкающих друг к другу концах клеток. Трахеиды — единственный тип водопроводящих клеток большинства споровых сосудистых растений и голосеменных; ксилема покрытосеменных, как правило, содержит и членики сосудов.
Рис. 20-12. Фотография в сканирующем электронном микроскопе трех члеников сосуда вторичной ксилемы дуба красного (Quercus rubra). Вид снаружи. Заметны ободки между ними

Интенсивное сравнительное изучение трахеальных элементов у широкого круга сосудистых растений позволило установить, что трахеиды — более примитивный (менее специализированный) тип клеток, чем членики сосудов — главные водопроводящие элементы ксилемы цветковых. Вполне вероятно, что последние возникли из трахеид независимо в нескольких группах сосудистых, включая двудольные и однодольные (покрытосеменные), гнетовые (содержащие сосуды голосеменные), некоторые неродственные виды папоротников, несколько видов селагинелл (плауновидные) и хвощи. Эволюция членика сосуда — прекрасный пример конвергентной эволюции, т. е. развития сходных структур у неродственных или находящихся в дальнем родстве организмов (см. приложение «Конвергентная эволюция» к гл. 22).
Термин трахеальный элемент был введен в семнадцатом столетии итальянским врачом Марчелло Мальпиги, одним из основателей анатомии растений, стремившимся обнаружить черты сходства между ними и животными. Исследуя ксилему, он наблюдал пузырьки воздуха, выделявшиеся из сосуда со спиральными утолщениями оболочек, что позволило ему сравнить эту структуру с трахеей, т. е. воздухопроводящей трубкой насекомых, и позже назвать так же все сосуды ксилемы. С тех пор термин используется для обозначения водопроводящих клеток ксилемы в целом.
Обычно считается, что членики сосудов более эффективно проводят воду, чем трахеиды, так как она может относительно беспрепятственно перетекать из членика в членик через перфорации, в то время как при движении через трахеиды ей приходится преодолевать мембраны пар пор (см. гл. 2). Однако эти мембраны, вероятно, относительно мало препятствуют току воды, поскольку на последних стадиях дифференцировки частично гидролизуются, сохраняя только хорошо проницаемый каркас из целлюлозных микрофибрилл.
Во вторичной и поздней первичной ксилеме (метаксилеме) вторичные стенки трахеид и члеников сосудов покрывают всю их первичную оболочку, за исключением участков пор и перфораций (рис. 20-10, А — Г). Впоследствии эти оболочки становятся жесткими и перестают растягиваться. В период роста в длину корней, стеблей и листьев у многих сформировавшихся первыми трахеальных элементов ранней первичной ксилемы (протоксилемы) вторичные оболочки образуют кольца или спирали (рис. 20-13). Такие кольчатые и спиральные утолщения позволяют трахеальным элементам растягиваться, хотя они часто и разрушаются во время общего удлинения органа. В первичной ксилеме на характер утолщения оболочки в значительной степени влияет интенсивность растяжения. Если оно слабое, возникают в основном пористые элементы, а если сильное, появляется много элементов с кольчатыми и спиральными утолщениями. На рис. 20-14 показаны некоторые стадии дифференцировки членика сосуда со спиральными утолщениями.
Рис. 20-13. Части трахеальных элементов ранней первичной ксилемы (протоксилемы) клещевины (Ricinus communis). А. Кольчатые (слева) и спиральные утолщения клеточной оболочки в несколько растянутых элементах. Б. Утолщения в виде двойной спирали у растянутых элементов. Элемент слева растянут так сильно, что витки спирали у него далеко разошлись
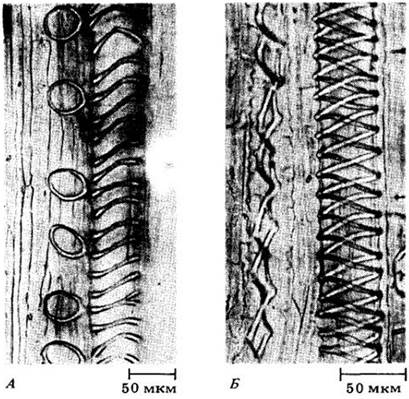
Рис. 20-14. Cxe.ua развития членика сосуда. А. Молодой сильно вакуолизированный членик без вторичной оболочки. Б. Клетка расширилась, появляются вторичные утолщения; первичная стенка в месте будущей перфорации утолщается. В. Формирование вторичных утолщений закончено, и клетка находится на стадии лизиса; ядро деформировано, тонопласт разорван, клеточная стенка в месте перфорации разрушается. Г. Зрелая клетка без протопласта, открытая с обоих концов

Кроме трахеид и члеников сосудов ксилема включает паренхимные клетки, запасающие различные вещества. Они обычно образуют вертикальные тяжи, а во вторичной ксилеме находятся также в лучах. В ксилеме также присутствуют волокна (см. рис. 20-10, Д, Е) и склереиды. Многие из этих волокон в зрелом состоянии остаются живыми и выполняют двойную — запасающую и опорную — функцию.
Флоэма
Флоэма — главная проводящая питательные вещества ткань сосудистых растений (см. рис. 20-9). Она может быть первичной и вторичной по происхождению. Как и первичная ксилема, ранняя первичная флоэма (протофлоэма) часто растягивается и разрушается при удлинении органа.
Основные проводящие клетки флоэмы — ситовидные элементы двух типов — ситовидные клетки (рис. 20-15) и членики ситовидной трубки (рис. 20-16 и 20-17). Термин «сито» относится к скоплениям пор (ситовидным полям), сквозь которые соединяются друг с другом протопласты соседних ситовидных элементов. В ситовидных клетках поры узкие, и структура ситовидных полей на всех стенках довольно однообразна. Большая часть полей сосредоточена на стыкующихся концах этих длинных узких клеток (рис. 20-15, А). В члениках ситовидных трубок канальцы ситовидных полей на одних стенках шире, чем на других. Часть стенки, несущая ситовидные ноля с более крупными канальцами, называется ситовидной пластинкой (рис. 20-16, Б и 20-17, А, Б). Хотя ситовидные пластинки могут встречаться на всех стенках, они находятся главным образом на концах клеток. Членики ситовидных трубок располагаются впритык друг за другом продольными рядами, называемыми ситовидными трубками. Таким образом, принципиальное различие между двумя типами ситовидных элементов — присутствие ситовидных пластинок у члеников ситовидных трубок и их отсутствие у ситовидных клеток.
Рис. 20-15. А. Продольный (радиальный) срез вторичной флоэмы тисса (Taxus canadensis) с вертикально ориентированными ситовидными клетками, тяжами паренхимных клеток и волокон. Можно видеть участки двух горизонтальных лучей, пересекающих вертикальные клетки. Б. Участок вторичной флоэмы тисса; видны ситовидные поля с каллозой (окрашена в синий цвет) на стенках ситовидных клеток и альбуминовые клетки (см. с. 25), образующие здесь верхний ряд клеток луча
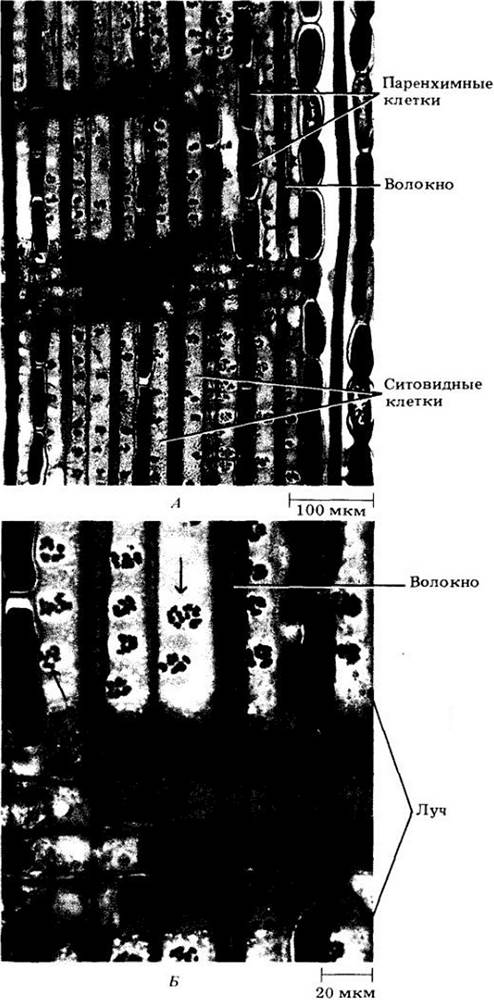
Рис. 20-16. А. Продольный (радиальный) срез вторичной флоэмы липы (Tilia americana) с члениками ситовидной трубки и хорошо заметными группами толстостенных волокон. Б. Сложные (состоящие из двух шли более ситовидных полей) ситовидные пластинки члеников ситовидных трубок липы. Каждое ситовидное поле пронизано канальцами, окаймленными цилиндрами каллозы, которая на этом срезе окрашена в синий цвет
Ситовидная клетка более примитивна, чем членик ситовидной трубки. У большинства споровых сосудистых растений и голосеменных это единственный тип клеток, проводящих питательные вещества, в то время как у покрытосеменных присутствуют только членики ситовидных трубок.
Оболочка ситовидных элементов обычно считается первичной. На срезах флоэмной ткани норы ситовидных полей и канальцы ситовидных пластинок, как правило, запечатаны углеводом каллозой, представляющим собой полисахарид из спирально закрученных цепей остатков глюкозы. Присутствие ее здесь долгое время озадачивало ботаников; казалось нелогичным, что участки, предназначенные для передвижения веществ от клетки к клетке, содержат соединение, по- видимому затрудняющее этот транспорт. Теперь выяснилось, что большая часть каллозы, если не вся она, откладывается здесь в ответ на повреждение, связанное с изготовлением микропрепарата.
В противоположность трахеальным ситовидные элементы в зрелом состоянии сохраняют живые протопласты, отличающиеся от протопластов всех прочих живых клеток растения тем, что полностью лишены ядра или содержат только его остатки. Кроме того, в большинстве зрелых ситовидных элементов отсутствует отчетливая граница между цитоплазмой и вакуолями. В молодом ситовидном элементе — несколько вакуолей, каждая из которых отделена от цитоплазмы тонопластом, или вакуолярной мембраной. На заключительной стадии дифференцировки тонопласты исчезают и разграничение между цитоплазматическим и вакуолярным содержимым перестает существовать. В зрелом ситовидном элементе все оставшиеся компоненты протопласта распределяются вдоль стенок; они представлены плазматической мембраной, гладким эндоплазматическим ретикулумом вблизи нее и небольшим числом пластид и митохондрий. Рибосомы, диктиосомы, микротрубочки и ядра отсутствуют.
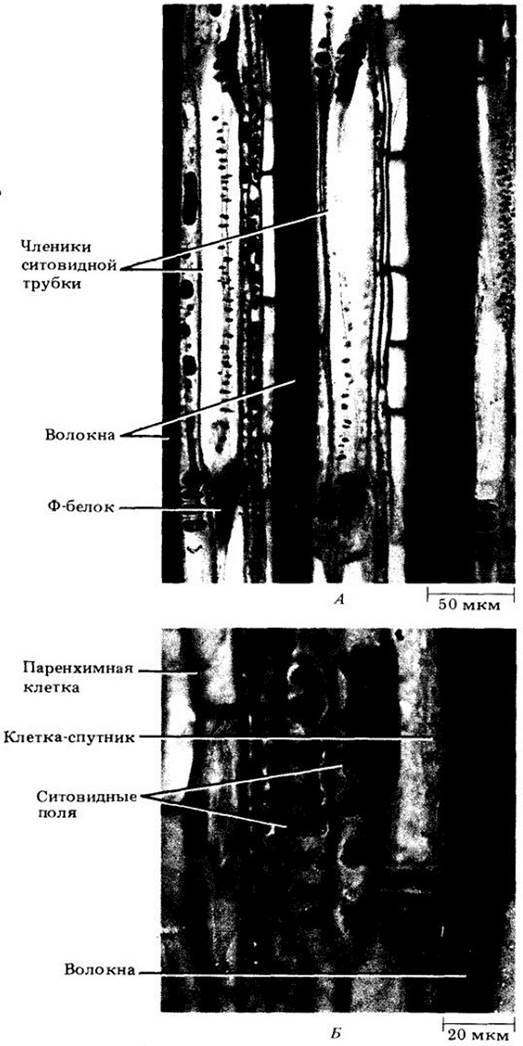
Для протопластов члеников ситовидных трубок двудольных (и некоторых однодольных) характерно наличие белкового вещества, так называемой слизи, или Ф-белка («Ф» обозначает флоэму). Ф-белок появляется в молодом членике в виде дискретных слизевых, или Ф-белковых, тел (рис. 20-17, В, Д), удлиняющихся и распадающихся на последней стадии дифференцировки. На срезах флоэмной ткани «слизевые пробки» Ф-белка обычно наблюдаются вблизи ситовидных пластинок (рис. 20-17, Г). Как и каллоза, они отсутствуют в неповрежденных клетках, поэтому считаются результатом травмирования содержимого члеников во время приготовления микропрепарата. В нормальных зрелых члениках ситовых трубок Ф-белок, по-видимому, располагается вдоль стенок, переходя из клетки в клетку через канальцы ситовидных полей и ситовидных пластинок, т. е. канальцы выстланы, а не закупорены им (рис. 20-17, А, Б). Функция Ф-белка не выяснена, однако некоторые ботаники полагают, что вместе с раневой каллозой он участвует в закупорке канальцев при повреждении ткани, предотвращая тем самым вытекание содержимого из ситовидных трубок.
Рис. 20-17. Флоэма стебля тыквы (Cucurbita maxima) на электронных (А, Б) и световых (В — Д) микрофотографиях. А. Продольный срез участков двух зрелых члеников ситовидных трубок и ситовидной пластинки, на котором видно пристенное распределение Ф- белка (см. стрелки). Б. Простая (с одним ситовидным полем) ситовидная пластинка между двумя зрелыми члениками ситовидной трубки. Канальцы открыты. В. Поперечный срез с двумя незрелыми члениками ситовидной трубки. Слизевые тельца, или тельца Ф-белка, можно видеть в членике слева, а незрелую ситовидную пластинку — в членике справа выше. Заметны мелкие, густо окрашенные клетки-спутники. Г. Поперечный срез со зрелыми члениками ситовидных трубок. В членике слева можно видеть слизевую пробку, а в членике справа — зрелую ситовидную пластинку. Заметны мелкие, густо окрашенные клетки-спутники. Д. Продольный срез со зрелыми и незрелыми члениками ситовидных трубок. Стрелками показаны Ф-белковые тельца в незрелых клетках
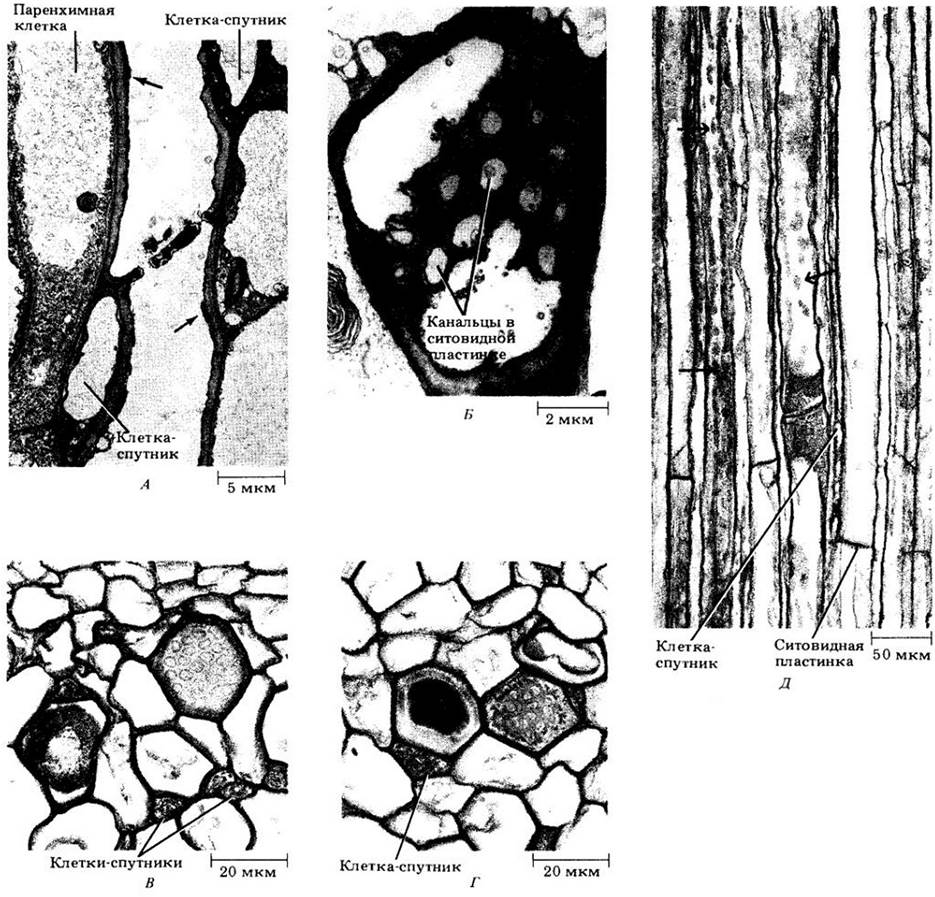
Членики ситовидных трубок обычно сопровождаются специализированными паренхимными клетками-спутниками (рис. 20-17, В — Д), содержащими все типичные компоненты растительных клеток, включая ядро. Членик и связанные с ним клетки-спутники имеют общее происхождение (возникают из одной и той же материнской клетки) и соединены между собой многочисленными плазмодесмами. На рис. 20-18 показаны некоторые стадии дифференцировки членика ситовидной трубки с Ф-белком. Функции клеток- спутников очень важны, так как они в значительной мере ответственны за активную секрецию веществ в членики ситовидной трубки и выведение их оттуда. Этот вопрос будет обсуждаться в гл. 27 при детальном рассмотрении механизма флоэмного транспорта у покрытосеменных.
Рис. 20-18. Схема дифференцировки членика ситовидной трубки. А. Его материнская метка делится. Б. В результате формируются молодой членик ситовидной трубки и клетка-спутник. После деления в цитоплазме, отделенной от вакуоли тонопластом, возникло одно или несколько Ф-белковых тел. Стенки членика на этой стадии утолщены, и места будущих канальцев ситовидной пластинки представлены плазмодесмами. В. Ядро дегенерирует, тонопласт разрушается, и Ф-белковые тельца равномерно распределяются по цитоплазме, выстилая клеточные стенки; одновременно плазмодесмы развивающихся ситовидных тгастинок начинают расширяться в канальцы. Г. В зрелом членике ситовидной трубки ядро и вакуоль отсутствуют. Все остальные компоненты протопласта, включая Ф-белок, располагаются вдоль стенок; канальцы ситовидных пластинок открываются

Ситовидные клетки голосеменных, как правило, сопровождаются специализированными паренхимными альбуминовыми клетками (рис. 20-15, Б). Хотя они обычно не связаны с ситовидными клетками общим происхождением из одной материнской клетки, считается, что их функции аналогичны выполняемым клетками-спутниками. Как и последние, альбуминовые клетки содержат ядро и другие типичные цитоплазматические компоненты живых клеток.
Ситовидные элементы большинства видов, вероятно, недолговечны и отмирают менее чем через год после своего появления. Однако это справедливо не во всех случаях. Во вторичной флоэме липы американской (Tilia americana) некоторые ситовидные элементы остаются живыми и, по- видимому, выполняют проводящие функции в течение 5 — 10 лет. Известно также, что они живут по многу лет у многолетних однодольных, в том числе более ста лет у некоторых пальм в основании главного стебля. Когда ситовидные элементы отмирают, гибнут и сопровождающие их клетки-спутники или альбуминовые клетки, что может рассматриваться как еще одно доказательство тесной связи между этими структурами.
В первичной и вторичной флоэме встречаются и другие паренхимные клетки (рис. 20-15 — 20-17), служащие главным образом для запасания, а иногда также волокна (рис. 20-15 и 20-16) и склереиды.
Эпидерма
Эпидерма — наружный слой клеток первичного тела растения, образующий систему покровной ткани листьев, частей цветка, плодов и семян, а также стеблей и корней, еще не подвергшихся существенному вторичному утолщению. Эпидермальные клетки весьма разнообразны по структуре и функциям. Эпидерма кроме обычных своих клеток, образующих основную ее площадь, может включать устьица (рис. 20-19, 20-21), разнообразные придатки, или трихомы (рис. 20-22), а также другие типы клеток, специализированные для выполнения особых функций.
Рис. 20-19. Поверхность ним ней эпидермы листа эвкалипта (Eucalyptus globulus) в сканирующем электронном микроскопе. Можно видеть устьице и многочисленные волокнистые отложения эпикутикулярного воска
Рис. 20-20. Электронная микрофотография устьица кукурузы (Zea mays). Поперечный срез через зрелые толстостенные замыкающие клетки, каждая из которых примыкает к околоустьичной клетке
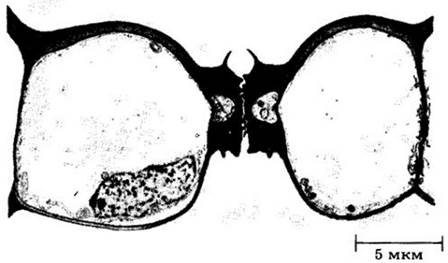
Рис. 20-21. Развивающееся устьице сверху (А) и в разрезе (Б). В. Схема зрелого устьица, показывающая его связь с эпидермой и субэпидермальными клетками. Замыкающие клетки возникают при неравном делении протодермальной клетки. Меньшая из двух образующихся при нем клеток называется материнской клеткой устьица или замыкающих клеток; именно ее деление непосредственно дает две замыкающие клетки (А, Б). После формирования замыкающих клеток межклеточное вещество их общей стенки набухает и растворяется, оставляя щель. Этот процесс сопровождается неравномерным утолщением оболочек, замыкающих клеток: стенки у щели гораздо толще, чем примыкающие к соседним эпидермальным клеткам. Зрелые устьица могут быть приподняты над поверхностью эпидермы или погружены в нее. Часто непосредственно под ними находится крупная воздушная, или дыхательная, полость. В отличие от обычных эпидермальных замыкающие клетки содержат хлоропласты

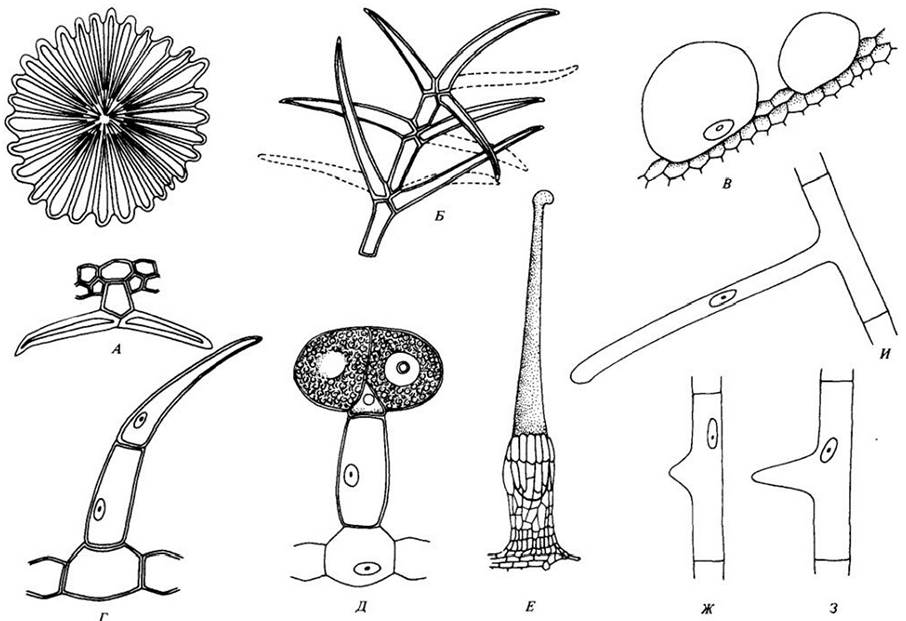
Рис. 20-22. Трихомы. А. Вид сверху (вверху) и на срезе (внизу) щитовидного волоска, или чешуйки, листа маслины (Olea europea). Б. Древовидный волосок платана (Platanus orientalis). В. Водяной пузырек «хрустальной травки» (Mesembryanthemiun crystallinum). Г. Короткий не разветвленный волосок стебля томата (Lycopersicon esculentum). Д. Железистый волосок стебля томата. Е. Жгучий волосок крапивы (Urtica). Он состоит из длинной игловидной части и широкого основания, окруженного другими эпидермальными клетками. Прикасаясь к телу, кончик обламывается, и ядовитое содержимое клетки (гистамин и ацетилхолин) впрыскивается в кожу. Ж — И. Стадии развития корневого волоска, простого трубчатого выпячивания эпидермальной клетки
У большинства растений эпидерма однослойная. Однако в протодерме листа некоторых растений происходят параллельные поверхности (периклинальные) деления, в результате чего образуется многослойная эпидерма (см. рис. 20-23). Она обнаружена, например, в листьях таких известных комнатных растений, как фикус (Ficus elástica) и пеперомия (Реperomia). Считается, что такая эпидерма служит водозапасающей тканью.
Рис. 20-23. Поперечный срез верхней части листовой пластинки каучуконоса Ficus elástico. Видна толстая кутикула, покрывающая многослойную эпидерму, состоящую преимущественно из крупных клеток. Лопатовидное образование в самой большой эпидермальной клетке состоит главным образом из карбоната кальция, отложенного на целлюлозном стержне. Ниже крупных светлых эпидермальных клеток располагаются клетки мезофилла

В основной своей массе эпидермальные клетки плотно примыкают друг к другу, обеспечивая хорошую механическую защиту частей растения. У надземных частей растений их стенки покрыты кутикулой, сводящей к минимуму потери воды и состоящей в основном из кутина и воска (см. гл. 3). У многих растений воск выделяется на поверхность кутикулы либо гладкими слоями, либо в виде палочек или нитей, поднимающихся над поверхностью (так называемого эпикутикулярного воска) (рис. 20-19; см. также рис. 3-11). Именно он создает беловатый или голубоватый налет на некоторых листьях и плодах.
Среди плоских, плотно сомкнутых эпидермальных клеток рассеяны специализированные, замыкающие клетки устьиц с хлоропластами (рис. 20-20 и 20-21), которые регулируют раскрытие устьичных щелей на надземных частях растения, тем самым контролируя проникновение в эти части и выход из них газов, включая пары воды (механизм открывания и закрывания устьиц обсуждается в гл. 27). Хотя устьица находятся на всех надземных органах, их особенно много на листьях. Они часто связаны с эпидермальными клетками особой формы, которые называются околоустьичными, или побочными (рис. 20-20).
Трихомы выполняют разные функции. Корневые волоски облегчают поглощение воды и минеральных веществ из почвы. Недавние исследования растений аридных местообитаний показали, что обильное опушение их надземных частей увеличивает отражение солнечной радиации, понижая гем самым их температуру и уменьшая испарение воды. Многие эпифиты, например, бромелиевые, используют трихомы листьев для поглощения воды и минеральных веществ. И напротив, у лебеды (Atriplex) секреторные трихомы удаляют из тканей листа токсичные соли, препятствуя их накоплению в растении. Трихомы могут защищать от насекомых. Например, у многих видов наблюдается положительная корреляция между густотой опушения и устойчивостью к поражению ими. Крючковатые волоски некоторых растений прокалывают насекомых и их личинок (рис. 20-24). Железистые (секреторные) волоски могут обеспечивать химическую защиту.
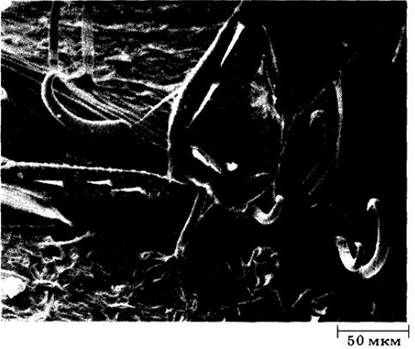
Рис. 20-24. Нимфы и взрослые особи цикадок (Empoasca sp.) часто захватываются крючковидными трихомами листьев фасоли (Phaseolus vulgaris). На этой фотографии, полученной с помощью сканирующего электронного микроскопа, виден трихом, погруженный в пленчатую ткань между члениками ноги взрослого насекомого
Перидерма
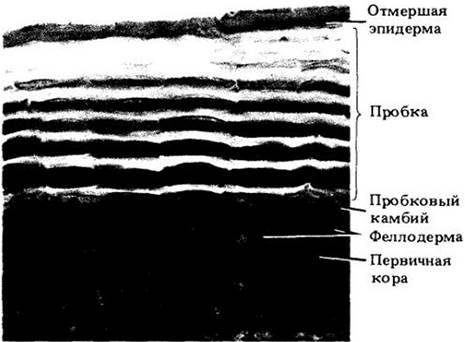
Перидерма обычно замещает эпидерму стеблей и корней в ходе их вторичного роста. Она состоит главным образом из защитной пробковой ткани (феллемы) из мертвых в зрелом состоянии клеток с сильно суберинизированными стенками, пробкового камбия (феллогена) и живой паренхимной ткани феллодермы (рис. 20-25). Феллоген образует на своей внешней поверхности пробковую ткань, а на внутренней — феллодерму. Происхождение пробкового камбия различно и зависит от вида и части растения. Перидерма будет детально рассматриваться в гл. 23.
Рис. 20-25. Поперечный срез перидермы ствола яблони (Malus sylvestris). Здесь она состоит главным образом из опробковевших меток, откладывающихся феллогеном наружу радиальными рядами. Внутрь от него находится единственный слой клеток феллодермы
Заключение
Общие сведения о растительных тканях и типах составляющих их клеток представлены в табл. 20-1, а признаки, локализация и функции разных типов клеток — в табл. 20-2.
Таблица 20-1. Растительные ткани и типы составляющих их клеток
Ткань |
Типы клеток |
Эпидерма |
В основном паренхимные; замыкающие клетки и трихомы; склеренхимные |
Перидерма |
В основном паренхимные; склеренхимные |
Ксилема |
Трахеиды; членики сосудов; склеренхимные; паренхимные |
Флоэма |
Ситовидные клетки или членики ситовидных трубок; альбуминовые клетки или клетки-спутники; паренхимные; склеренхимные |
Паренхима |
Паренхимные |
Колленхима |
Колленхимные |
Склеренхима |
Волокна или склереиды |
Таблица 20-2. Общая характеристика типов клеток
Тип клеток |
Признаки |
Локализация |
Функции |
Паренхима |
Форма обычно полиэдрическая (многогранная), разнообразная. Клеточные оболочки: первичная или первичная и вторичная, иногда лигнифицированные, суберинизированные или кутинизированные. Живые |
По всему растению в виде паренхимной ткани коры, сердцевины и сердцевинных лучей, в ксилеме и флоэме |
Дыхание, расщепление веществ, фотосинтез; накопление запасов, проведение веществ; заживление ран и регенерация |
Колленхима |
Форма вытянутая. Клеточная оболочка неравномерно утолщена, не лигнифицирована, только первичная. В зрелом состоянии живые |
На периферии (под эпидермой) в молодых удлиняющихся стеблях; часто в виде тканевого цилиндра или отдельными участками; в тяжах вдоль жилок некоторых листьев |
Опора первичного тела растения |
Волокна |
Форма, как правило, очень вытянутая. Клеточные оболочки: первичная и утолщенная вторичная (часто лигнифицированные). Часто (не всегда) мертвые в зрелом состоянии |
Иногда в коре стеблей, чаще всего связаны с ксилемой и флоэмой; в листьях однодольных |
Опорная |
Склереиды |
Форма разнообразная; как правило, короче волокон. Клеточные оболочки: первичная и утолщенная вторичная, как правило, лигнифицированные. В зрелом состоянии живые или мертвые |
По всему растению |
Механическая; защитная |
Трахеиды |
Форма вытянутая, концы скошенные. Клеточные оболочки: первичная и вторичная, лигнифицированные; пористые, но не перфорированные. В зрелом состоянии мертвые |
Ксилема |
Главные водопроводящие элементы голосеменных и споровых сосудистых растений; встречаются также у покрытосеменных |
Членики сосуда |
Форма вытянутая (обычно короче трахеид). Клеточные оболочки: первичная и вторичная, лигнифицированные, с порами и перфорациями; несколько члеников сосудов, смыкаясь концами, составляют сосуд. В зрелом состоянии мертвые |
Ксилема |
Главные водопроводящие элементы покрытосеменных |
Ситовидные клетки |
Форма вытянутая, со скошенными концами. Клеточная оболочка у большинства видов первичная, с ситовидными полями; часто в клеточной оболочке и в канальцах ситовидных полей присутствует каллоза. В зрелом состоянии живые с остатками ядра или без них; границы между вакуолью и цитоплазмой нет |
Флоэма |
Главные проводящие питательные вещества элементы голосеменных и споровых сосудистых растений |
Альбуминовые клетки |
Форма, как правило, удлиненная. Клеточная оболочка первичная. В зрелом состоянии живые; сопровождают ситовидные клетки, но обычно не образуются из общих с ними материнских клеток. Имеют многочисленные плазмодесмальные связи с ситовидной клеткой |
Флоэма |
По-видимому, участвуют в транспорте питательных веществ в ситовидную клетку и из нее |
Членики ситовидной трубки |
Форма вытянутая. Клеточная оболочка первичная, с ситовидными полями, которые на концах клетки имеют гораздо более крупные канальцы, чем на боковых стенках, и называются здесь ситовидными пластинками; с клеточной оболочкой и канальцами ситовидных полей часто связана коллоза. В зрелом состоянии живые, без ядра или только с его остатками; у двудольных и некоторых однодольных содержат так называемый Ф-белок; несколько члеников ситовидной трубки, смыкаясь концами, составляют ситовидную трубку |
Флоэма |
Главные проводящие питательные вещества элементы покрытосеменных |
Клетки-спутники |
Форма разнообразная, как правило, вытянутая. Клеточная оболочка первичная. В зрелом состоянии живые; тесно связаны с члениками ситовидной трубки и образуются из общих с ними материнских клеток. Имеют многочисленные плазмодесмальные связи с члениками ситовидной трубки |
Флоэма |
По-видимому, участвуют в транспорте питательных веществ в ситовидную трубку и из нее |