СОВРЕМЕННАЯ БОТАНИКА - П. РЕЙВН - 1990
РАЗДЕЛ VI. РЕГУЛЯЦИЯ И РОСТОВЫЕ РЕАКЦИИ
ГЛАВА 24. РЕГУЛЯЦИЯ РОСТА И РАЗВИТИЯ: ГОРМОНЫ РАСТЕНИЙ

Для своего роста растение нуждается в солнечном свете, в двуокиси углерода, которую получает из воздуха, а также в воде и минеральных компонентах (включая азот), поступающих из почвы. Из всего этого создаются вещества, свойственные самому растению, причем простые элементы превращаются в сложные органические молекулы, из которых и состоят живые существа. Как отмечалось в предыдущем разделе, рост не является простым увеличением массы и объема растения. Растение дифференцируется, развивается, приобретает определенную форму, образуя разнообразные клетки, ткани и органы. Каким образом единственная клетка (оплодотворенная яйцеклетка) может стать источником множества тканей и органов, которые образуют поразительный индивид, называемый «нормальным» растением? Многие детали регуляции этих процессов еще неизвестны, но становится ясно, что нормальное развитие растения зависит от взаимодействия внутренних и внешних факторов. Основные внутренние факторы, регулирующие рост и развитие растений, имеют химическую природу, они и являются предметом обсуждения данной главы. Некоторые внешние факторы — свет, температура, длина светового дня, гравитация и др. влияющие на рост — обсуждаются в гл. 25.
Рис. 24-1. Фотопериодизм побуждает растения многих видов переходить каждый год к цветению в одно и то же время. Цветение происходит в результате взаимодействия внешних (условий среды) и внутренних (гормональных) факторов. Фотография цветущей вишни (Prunus) в Бруклине, Нью-Йорке

Растительные гормоны играют основную роль в регуляции роста. Термин «гормон» был введен физиологами животных; им обозначают органические вещества, которые образуются в одной ткани и транспортируются в другую, где вызывают определенный физиологический эффект. Гормоны активны в очень небольших количествах. В побеге ананаса (Ananas comosus), например, содержится всего шесть микрограммов индолилуксусной кислоты, обычного растительного гормона, на килограмм растительной массы. Один предприимчивый физиолог растений вычислил, что массу гормона по отношению к массе побега можно сопоставить с массой иголки в 20 т сена.
Слово гормон произошло от греческого слова hormaein, которое означает «возбуждать». Однако сейчас известно, что многие гормоны оказывают тормозящее действие. Поэтому, может быть, лучше рассматривать их не как стимуляторы, а как химические регуляторы. Но и этот термин нуждается в уточнении, поскольку реакция на отдельный «регулятор» зависит не только от его состава (химической структуры), но и от восприимчивости реципиента (тканевой специфичности).
В дальнейшем будет показано, как постепенно накапливались наши знания о растительных гормонах. В соответствии с этим обсуждение начинается с ауксина; именно его открытие дало толчок изучению гормонов растений.
Ауксин
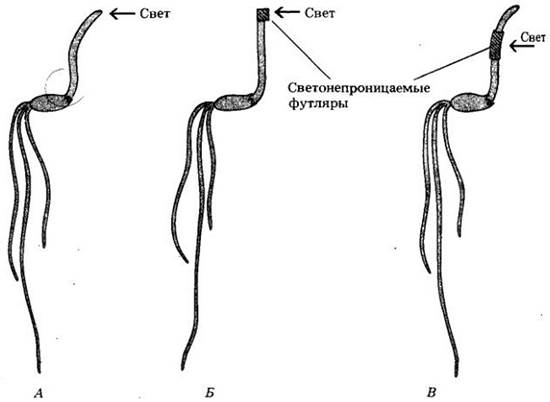
Одни из первых экспериментов по регуляции роста у растений были выполнены Чарльзом Дарвином и его сыном Фрэнсисом и изложены в работе «The Power of Movement inPlants» («Сила движения у растений»), опубликованной в 1881 г. Дарвины систематически наблюдали движение (изгибание) растений по направлению к свету (так называемый фототропизм), используя проростки канареечника (Phalaris canariensis) и овса (Avena sativa). Они показали, что если закрыть верхнюю часть колеоптиля (защитной структуры, укрывающей, подобно футляру, верхушку проростка злаков) цилиндром из фольги или полой стеклянной трубкой, зачерненной тушью, и осветить растение сбоку, то характерного изгибания не произойдет (рис. 24-2). Однако при использовании прозрачных стеклянных трубок искривление происходило как обычно. Согласно Дарвинам, эксперименты показали существование «средства сообщения» между верхушкой, т. е. тканью, воспринимающей световой стимул, и остальной частью колеоптиля, где осуществляется ростовой изгиб. И далее они формулируют: «Мы должны сделать заключение, что если проростки освещаются сбоку, то от верхней их части к нижней передается какой-то стимул, заставляющий последнюю изгибаться».
Рис. 24-2. Эксперимент Дарвинов. А. Проростки нормально изгибаются по направлению к свету. Б. Если верхушку проростка закрыть светонепроницаемым колпачком, изгибания не происходит. Оно наблюдается, если верхушка проростка закрывается прозрачным колпачком. В. Если колпачок помещается ниже верхушки, имеет место характерный изгиб. На основании этих экспериментов Дарвины сделали вывод, что под влиянием света «стимул», вызывающий изгибание, передается от верхушки проростка в нижележащую область, где обычно и происходит изгиб
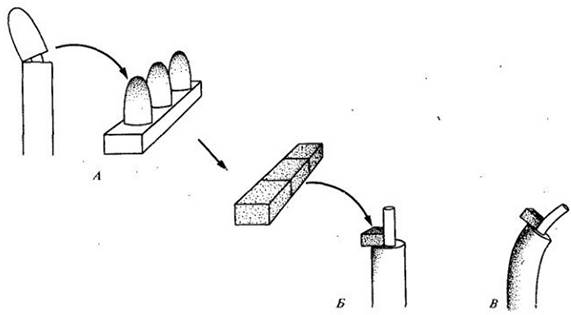
В 1926 г. голландскому физиологу растений Фрицу В. Венту удалось выделить этот «стимул» из верхушек колеоптилей. Вент срезл верхушки колеоптилей у проростков овса (Avena) и помещал их примерно на час на ломтики агара так, чтобы поверхности срезов пришли в контакт с агаром. (Агар — желатинообразное вещество — получают из некоторых красных водорослей и используют в качестве нейтральной ростовой среды.) Затем Вент нарезал агар на маленькие блоки и помещал их на одну сторону декапитированных колеоптилей, которые выдерживались в темноте в течение всего эксперимента. В течение часа он наблюдал отчетливое изгибание в сторону, противоположную той, на которую помещал агаровый блок (рис. 24-3). Агаровые блоки, на которые не накладывались верхушки колеоптиля, не вызывали никакого изгибания или способствовали лишь небольшому изгибанию в ту сторону, на которую был помещен блок. Агаровые блоки, на которые Вент помещал фрагменты колеоптиля, расположенные ближе к его основанию, тоже не оказывали физиологического действия.
Рис. 24-3. Эксперимент Вента. А. Вент удалял у проростков верхушки колеоптилей, помещал их на агар и выдерживал в течение часа. Б. Затем Вент нарезал агар на маленькие блоки, которые накладывал на одну сторону декапитированных проростков. В. Содержащиеся в темноте в ходе всего эксперимента проростки постепенно изгибавшись в сторону, противоположную той, на которой находился агаровый блок. На основании этого Вент сделал заключение, что «стимул», который вызывает изгибание проростка, имеет химическую природу и накапливается на противоположной от света стороне
Этими экспериментами Вент показал, что влияние верхушки колеоптиля связано скорее с выделением какого-то химического вещества, чем с физической, например, электрической, стимуляцией. Это химическое вещество Вент назвал ауксином, от греческого слова auxein — «увеличивать».
Изгибание колеоптиля овса в сторону, противоположную той, на которую помещают содержащий ауксин агаровый блок, вызывается асимметричным распространением ауксина, что в свою очередь способствует асимметричному увеличению размера клеток колеоптиля; клетки той стороны, на которой находится блок, больше увеличиваются в длину, чем клетки противоположной стороны. Условия выращивания проростков овса и размещения агарового блока были стандартизированы до такой степени, что угол изгиба (измеренный транспортиром) мог быть использован для определения количества ауксина в агаровом блоке. Применение этой методики, известной как биотест на изгиб колеоптиля овса, позволило исследователям выделить и идентифицировать встречающийся в природе ауксин, который был назван индолилуксусной кислотой (сокращенно ИУК). (Биотест позволяет количественно определить концентрацию вещества по его действию на рост соответствующего организма в контролируемых условиях.)
Как видно из рис. 24-4, ИУК очень сходна с аминокислотой триптофаном (см. рис. 3-14). Триптофан является предшественником ИУК в живом растении; при этом известны четыре пути биосинтеза ИУК, каждый со своим посредником. Различные группы растений используют неодинаковые пути образования ИУК из триптофана. Кроме того, некоторые растения, такие, как кукуруза (Zea mays), используют различные пути на разных стадиях развития. Ауксин образуется в кончиках колеоптиля злаков и в верхушках побегов. Хотя ИУК была найдена и в кончиках корней, есть много данных, указывающих на то, что она транспортируется сюда по проводящим тканям из основания корня. Большое количество ИУК, возможно, содержится в зародышах; кроме того, она обнаружена в молодых листьях, цветках и плодах.
Вскоре после того как был открыт ауксин и выяснена его роль в регуляции растяжения клеток, обнаружили его тормозящее влияние на рост боковых почек. Например, если удалить апикальную меристему бобового растения, боковые почки начинают расти. Однако если на поверхность среза сразу же нанести ауксин, рост почек задерживается. Как и в случае фототропической реакции проростков овса, это апикальное доминирование свидетельствует о «взаимодействии» между двумя растительными тканями посредством ИУК. Более того, в обоих случаях «стимул» перемещается от растущей верхушки растения к его основанию. Это происходит потому, что ауксин активно транспортируется от верхушки побега к основанию растения, т. е. в базипетальном направлении. Передвижение скорее происходит по ткани в целом, чем по проводящим элементам ксилемы и флоэмы (соответственно сосудам и ситовидным трубкам). Транспорт, предположительно, связан с взаимодействием ИУК и плазматических мембран растительных клеток.
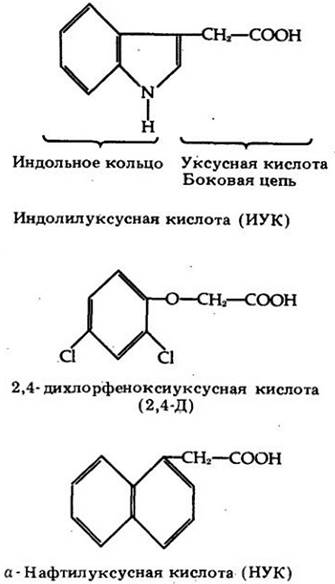
Ауксин оказывает разнообразное влияние на растение в зависимости от времени воздействия, вида растения и особенно от типа ткани. Подобно многим другим физиологически активным соединениям, ауксин токсичен в высоких концентрациях. Уничтожающий сорняки гербицид 2,4-Д является синтетическим ауксином, одним из многих, которые производятся в практических целях для широкого применения (рис. 24-4).
Рис. 24-4. Индолилуксусная кислота (ИУК) — наиболее известный из встречающихся в природе ауксинов. Дихлорфеноксиуксусная кислота (2,4-Д) — синтетический ауксин, широко используемый в качестве гербицида. α-Нафтил- уксусная кислота (НУК) — другой синтетический ауксин — применяется обычно для стимулирования образования придаточных корней у черенков и уменьшения опадения плодов, в садах. Синтетические ауксины в отличие от ИУК нелегко разрушаются ферментами растений и микроорганизмов и поэтому лучше подходят для практического использования
Ауксин и клеточная дифференцировка
Ауксин оказывает влияние на дифференцировку проводящей ткани в растущем побеге. Если вырезать сектор из стебля Coleus таким образом, чтобы перерезать и удалить части сосудистых пучков, то из клеток сердцевины образуются новые проводящие ткани, которые установят связь с пучками неповрежденных участков. Если удалить листья и почки выше места среза, то формирование новых клеток задержится. Если добавить ИУК к оставшемуся черешку листа, расположенному как раз над местом среза, формирование сосудистой ткани возобновится. Ауксин играет важную роль в присоединении сосудистых тяжей развивающихся листьев к сосудистым пучкам стебля.
Сходные реакции отмечаются в каллусах. (Каллус представляет собой массу недифференцированных клеток и образуется при ранении растения и культивировании изолированных клеток in vitro.) Если взять каллус сердцевины сирени (Syringa) и привить на него почку, в каллусе начинает формироваться сосудистая ткань. Сосудистая ткань образуется и при выращивании каллуса на среде, содержащей ауксин и сахар (сахар необходим, поскольку в каллусе нет фотосинтезирующих клеток). Р. Уэтмор с сотр. показал, что, регулируя количество сахара в среде, можно вызвать формирование только ксилемы, ксилемы и флоэмы или только флоэмы. Низкая концентрация сахарозы (от 1,5 до 2,5%) способствует образованию ксилемы, высокая (4%) — образованию флоэмы, а промежуточная — образованию обеих тканей. Это указывает на большую чувствительность регуляторов роста и привлекает внимание к тому немаловажному факту, что гормоны никогда не действуют независимо, а только во взаимосвязи с другими веществами или гормонами.
Ауксин и камбий
У древесных растений ауксин способствует росту камбия. Когда весной начинают делиться клетки меристематической зоны побега, ауксин, поступая из верхушки побега, стимулирует деление клеток камбия, образование вторичной флоэмы и вторичной ксилемы. Вместе с тем эти процессы модулируются в растительном организме и другими регуляторами роста.
Опыты с применением экзогенной ИУК и гибберелловой кислоты показывают, что в интактном растении взаимодействие между ауксинами и гиббереллинами определяет относительную скорость образования вторичной флоэмы и вторичной ксилемы. Например, ИУК и гибберелловая кислота каждая по отдельности стимулируют активность камбия у многих древесных растений. Однако в отсутствие гибберелловой кислоты ИУК стимулирует только развитие ксилемы. Образование флоэмы происходит под влиянием одной гибберелловой кислоты, а максимальное совместное развитие как ксилемы, так и флоэмы наблюдается только в присутствии ИУК и гибберелловой кислоты.
Ауксин и рост корней
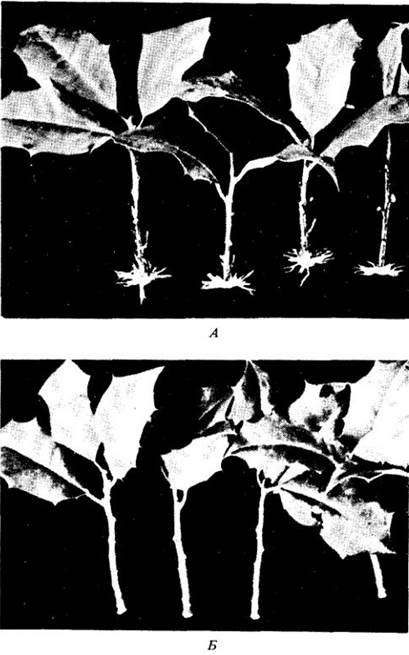
Первое практическое применение ауксина было основано на его способности стимулировать образование придаточных корней у черенков (рис. 24-5). Обработка черенков раствором ауксина имеет особенно большое значение для культивирования древесных растений в питомниках, поскольку позволяет размножать их в больших масштабах вегетативным путем. Однако применение значительных доз ауксина к уже растущим корням обычно тормозит их дальнейший рост.
Рис. 24-5. Черенки падуба (А) обрабатывались ауксином за 21 день до того, как была сделана фотография. Б. Черенки не обрабатывались. Обратите внимание на появление придаточных корней у обработанных растений
Ауксин и рост плодов
Ауксин ускоряет рост плодов. Обычно если цветок не опылен и оплодотворения не произошло, плод не развивается. У некоторых растений для нормального развития плода достаточно оплодотворения одной яйцеклетки; у других же, имеющих многосемянные плоды, например, яблони или дыни, для того чтобы стенка завязи развилась и стала мясистой, необходимо оплодотворение нескольких яйцеклеток. Обрабатывая женские части цветка некоторых растений ауксином, можно добиться получения партенокарпического плода (от слова parthenos, означающего «девственный»), т. е. плода, развившегося без оплодотворения, например, бессемянных томатов, огурцов, баклажанов.
Кроме того, ауксин содержится и в образующихся семенах. У садовой земляники (Fragaria ananassa) семена находятся в плодиках-семянках, погруженных в разросшееся цветоложе, которое неправильно называют плодом. Если семянки удалить из цветоложа во время его развития, последнее перестанет расти. Если удалить узкое кольцо семянок, то цветоложе разрастается только в зоне оставшихся. При воздействии ауксином на цветоложе, лишенное семянок, рост будет продолжаться нормально (рис. 24-6).
Рис. 24-6. А. Нормально развитый «плод» земляники (Fragaria ananassa). Б. Удалены все семянки. В. Оставлены три горизонтальных ряда семянок. Г. Оставлена одна семянка. Д. Удалены все семянки, кроме трех. Если пасту, содержащую ауксин, нанести на цветоложе с удаленными семянками, оно разовьется в нормальный «плод»

Ауксин и опадение частей растения
Ауксин образуется в молодых листьях, но, по-видимому, он не оказывает прямого влияния на скорость роста листа. Тем не менее ауксин воздействует на опадение —сбрасывание листьев или других частей растения. По мере старения листьев некоторые ионы и молекулы, способные к реутилизации, такие, как ионы магния, аминокислоты (продукты разрушения белков) и сахара (продукты гидролиза крахмала), возвращаются в стебель. Затем, по крайней мере у некоторых растений, ферменты разрушают оболочки клеток отделительной зоны у основания черешка (см. рис. 22-32). При этом происходят растворение срединных пластинок и гидролиз целлюлозных волокон. Отделению листа могут предшествовать клеточные деления. Если они происходят, то вновь сформировавшиеся клеточные оболочки, как правило, подвергаются деградации. Под отделительным слоем образуется защитный слой, представленный здоровыми суберинизированными клетками, который перед опадением изолирует лист от остальных частей растения. В конце концов лист удерживается на растении благодаря лишь нескольким тяжам сосудистой ткани, которые могут разрушиться, но мере разрастания паренхимных клеток отделительного слоя. Наряду с другими факторами опадение коррелирует с уменьшением содержания ауксина в листе; во многих случаях оно может быть предотвращено применением ауксина.
Умение регулировать опадение листьев, цветков и плодов имеет большое значение в сельском хозяйстве. Ауксин, а в последнее время и этилен используются для обработки некоторых видов растений. Например, ауксин предотвращает опадение листьев и ягод у вечнозеленого падуба (Ilex aquifolium) и тем самым уменьшает потери при транспортировке. Ауксин предупреждает и преждевременное опадение плодов цитрусовых, что позволяет получать более высокие урожаи апельсинов и грейпфрутов. С другой стороны, большое количество ауксина, вызывая повышенное образование этилена, способствует опадению плодов. На основании этого ауксин используется для их прореживания при выращивании олив, яблонь и других плодовых деревьев.
Ауксин и борьба с сорняками
Синтетические ауксины широко используются в сельском хозяйстве для борьбы с сорняками. В экономическом отношении именно в этом сегодня состоит основное практическое применение регуляторов роста растений во всем мире. Хотя известен ряд соединений, тем не менее феноксиауксины, например, 2,4-Д и его химические производные, составляют примерно 20% всех используемых в данных целях соединений. Дальнейшее применение этих веществ зависит от ряда факторов, в том числе их окупаемости и потенциальной или реальной опасности для здоровья человека.
Каким образом ауксин регулирует растяжение клетки?
Ауксин увеличивает пластичность клеточной оболочки. Когда клеточная оболочка размягчается, клетка увеличивается в размерах за счет тургорного давления. По мере уменьшения тургорного давления вследствие растяжения клетки она поглощает больше воды и продолжает разрастаться до тех пор, пока не встретит достаточное сопротивление со стороны оболочки (см. гл. 4).
Размягчение клеточной оболочки происходит в результате сложных взаимодействий. Одним из первых проявлений действия ауксина можно считать изменения метаболизма клетки, которые вызывают быстрое выкачивание протонов через плазматическую мембрану. Происходящее при этом подкисление клеточной оболочки неизвестным образом приводит к гидролизу сшивок внутри нее и вследствие этого к растяжению клетки за счет тургорного давления. Проверка этой так называемой «гипотезы кислого роста» привела к значительному прогрессу в понимании действия ауксина. Она подтверждается разносторонними экспериментальными доказательствами, включая действие ауксина на выход протонов, вызванное кислыми растворами (pH 5,0 — 5,5) удлинение клеток (сходное с действием ауксина) и ингибирующее действие нейтральных буферов (препятствующее подкислению оболочек) на растяжение клеток под влиянием ауксина.
Хотя «гипотеза кислого роста» и предполагает начальное растяжение клетки под влиянием ауксина, она не может объяснить более длительное его действие на растение. Действительно, большинство исследователей соглашается теперь с тем, что существуют два типа воздействия ауксина на растяжение клетки: быстрое, кратковременное воздействие, вызываемое кислым ростом, и второе, продолжительное воздействие, связанное с регуляцией экспрессии генов. Так, было показано, что ауксин вызывает экспрессию по крайней мере 10 специфических генов, предположительно всех, которые ответственны за процессы роста. Известно далее, что ауксин действует на транскрипцию. Влияние ауксина на экспрессию генов у растений, по-видимому, сходно с действием некоторых гормонов животных.
Цитоксины
Открытие ауксина стимулировало поиск других типов химических веществ, регулирующих рост растений, поскольку по аналогии с животными казалось маловероятным, что рост и развитие растений регулируются лишь одним гормоном. Например, было известно, что ауксин задерживает рост боковых почек, если применяется к декапитированным растениям. Имеются ли естественные гормоны, которые противодействуют влиянию ауксина?
Фолк Скуг с коллегами из Висконсинского университета разработал метод изучения гормональной регуляции роста почек в изолированных тканях и органах растений, культивируемых в пробирках. Если кусочек стебля табака (Nicotiana labacuni) помещался на культуральную среду, содержащую сахар, витамины и различные соли, то клеточные деления, происходящие на поверхности среза, приводили к образованию недифференцированного каллуса и изредка к образованию побега, т. е. стебля с листьями. Добавление в среду достаточного количества ауксина задерживало, как и предполагалось, формирование и рост почек. С другой стороны, аденин способствовал формированию почек в культуре ткани и препятствовал проявлению тормозящего действия ауксина. Концентрация аденина, необходимая для формирования почек, была очень высока, даже слишком, чтобы аденин можно было рассматривать как гормон, — поэтому Скуг и его коллеги приступили к поискам веществ с большей биологической активностью в экспериментальных системах.
Первоначально Скуг и его коллеги работали с кокосом (Cocos nucífera), поскольку ван Овербик, а позднее Ф. Стьюард со своей исследовательской группой показали, что кокосовое молоко (которое представляет собой жидкий эндосперм) является богатым источником веществ, способствующих росту культур ткани. После многолетних исследований Скугу и его сотрудникам удалось достигнуть тысячекратной очистки фактора роста, но выделить его они нс смогли. Тогда, меняя ход исследования, ученые испытали различные пуринсодержащие вещества — главным образом нуклеиновые кислоты, — надеясь обнаружить новые ростовые факторы.
Следуя этим новым путем, Карлос О. Миллер из лаборатории Скуга исследовал хранящиеся в лаборатории образцы нуклеиновых кислот известной видовой специфичности. При этом оказалось, что ДНК спермы сельди вызывает деление клеток табака. Были заказаны новые образцы ДНК спермы сельди, но, к изумлению исследователей, эти свежие препараты не работали. Тогда в последний раз Миллер обратился к другому старому образцу, и тот (!) оказался активным. Видимо, найденный фактор был лишь продуктом разрушения ДНК. Соответственно исследователи пришли к выводу, что различные препараты ДНК — старые или те, «старение» которых вызывалось искусственно нагреванием в кислом растворе, — содержали фактор, вызывающий деление клеток.
Миллеру, Скугу и их сотрудникам удалось в конце концов добиться успеха в выделении фактора роста из одного препарата ДНК и установить его химическую природу. Они назвали это вещество кинетинсм, а класс регуляторов роста, к которому он принадлежит, — цитокининами, поскольку они стимулируют цитокинез, т. е. деление клеток. Как показано на рис. 24-7, кинстин похож на пурин аденин, который можно считать его прототипом. Кинетин, однако, не найден у растений и не относится к числу природных цитокининов. Он имеет относительно простую структуру, и биохимики вскоре синтезировали ряд других соединений, по биологической активности близких к цитокининам. В конце концов первый природный цитокинин был выделен из зерен кукурузы (Zea mays) и назван зеатином. Он считается самым активным из природных цитокининов; в настоящее время синтезировано несколько более активных соединений.
Рис. 24-7. Обратите внимание на сходство между пурином аденином и этими четырьмя цитокининами. Кинетин и 6-бензиламинопурин (БАП) — синтетические цитокинины, обычно используемые в практике. Зеатин и i6Ade выделены из растений

Цитокинины выделены из многих семенных растений, где в основном обнаружены в активно делящихся тканях, включая семена, плоды и корни. Эти гормоны найдены и в пасоке — вытекающем из мест срезов, сломов и других ран, соке многих растений. Недавно цитокинины были идентифицированы у двух видов бессемейных растений: хвоща Equisetum arvenseи папоротника Dryopieris crassirhizoma.
Хотя практическое применение цитокининов не так широко, как у ауксина, они имеют большое значение для исследований регуляции развития растений. Поскольку применение цитокининов занимает центральное место в экспериментах с культурами тканей (см. приложение), они, несомненно, будут играть очень важную роль в биотехнологии будущего. Обработка боковых почек цитокинином, как правило, вызывает их рост даже в присутствии ауксина, изменяя тем самым апикальное доминирование.
Цитокинины и деление клеток
Изучение взаимодействий ауксина с цитокининами помогает физиологам понять, каким образом растительные гормоны обеспечивают развитие структуры, свойственной тому или другому растению. По-видимому, недифференцированная растительная клетка может следовать по одному из двух путей: либо она растет, делится и снова растет, и делится, либо же растягивается, не подвергаясь делению. Клетка, которая неоднократно делится, остается по существу недифференцированной, или эмбриональной, в то время как растягивающаяся клетка дифференцируется и становится специализированной. Добавление к культуре ткани стебля табака ИУК вызывало быстрое растяжение и формирование гигантских клеток. Добавление одного лишь кинетина вызывало слабый эффект или вообще никакого. ИУК вместе с кинетином способствовала быстрому делению и появлению большого числа относительно мелких недифференцированных клеток. Другими словами, добавление кинетина вместе с ИУК (но не одного кинетина) переключало клетки на путь меристематической деятельности.
В другой работе с культурой ткани топинамбура (Helianthus tuberosus) было показано, что ионы кальция могут влиять на совместное действие ауксина и цитокинина. Как было показано, совместное применение ИУК и низких концентраций кинетина способствовало увеличению размеров клеток, в то время как добавление в культуральную среду Са2+вызывало устойчивое смещение ростовых процессов от растяжения клеток к их делению. Высокие концентрации кальция препятствуют растяжению клеточных оболочек, клетка меняет направление своего развития и делится. Таким образом, не только гормоны влияют на активность друг друга, но в эти сложные взаимодействия могут включаться и негормональные факторы, например, кальций.
Цитокинины и формирование органов в культуре тканей
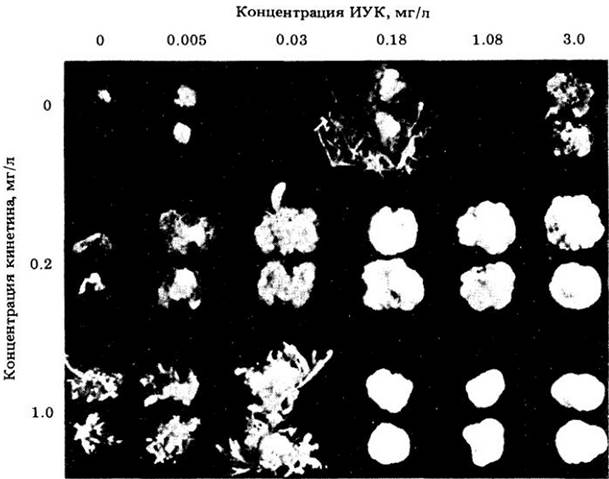
Высокая концентрация ауксина способствует образованию корней в ткани каллуса. В каллусной ткани сердцевины табака соответствующие концентрации ауксина и кинетина вызывают образование корней или почек: при высоком содержании ауксина образуются корни; при высокой концентрации кинетина формируются почки: если оба гормона присутствуют примерно в равных количествах, в каллусе продолжается образование недифференцированных клеток (рис. 24-8).
Рис. 24-8. Влияние возрастающей концентрации ИУК при различном содержании кинетина на рост и формирование органов в каллусе табака, культивируемом на питательном агаре. Можно видеть, что без добавления ИУК или кинетина происходит незначительный рост. Более высокие концентрации ИУК способствуют развитию корней, но задерживают формирование почек (последний эффект наблюдается и в том случае, если ИУК используют в комбинации с кинетином). По мере повышения концентрации кинетина усиливается развитие почек, однако приведенные концентрации слишком высоки, чтобы способствовать росту корней
Цитокинины и старение листа
У большинства видов листья начинают желтеть, как только их удаляют с растения. Это пожелтение, связанное с утратой хлорофилла, можно предупредить обработкой цитокининами. Если, например, срезанные листья дурнишника (Xanthium strumarium) поместить в чистую воду, то они пожелтеют примерно через 10 дней. Если же в воде присутствует кинетин (10 мг/л), хлорофилл нс разрушается и сохраняется свежий вид листа. Если на срезанные листья пятнами нанести раствор, содержащий кинетин, то пятна остаются зелеными, в то время как остальные части листа желтеют. Более того, если такой лист содержит радиоактивные аминокислоты, меченные 14С, можно увидеть, что аминокислоты мигрируют из других частей листа в те зоны, которые обработаны цитокинином. Подобные исследования, проведенные на редьке и других растениях, позволили высказать гипотезу, что старение листьев, а возможно, и других частей растения происходит вследствие постепенного «выключения» отдельных генов с последующей потерей способности к синтезу мРНК и белка. Было сделано предположение, что цитокинины препятствуют выключению генов и тем самым способствуют продолжению синтеза ферментов и образованию таких соединений, как хлорофилл.
Одно из объяснений предотвращения старения отдельных листьев при помощи цитокинина заключается в том, что листья в норме не синтезируют достаточно кинетина, чтобы удовлетворить собственные потребности. Таким образом, до сих пор неизвестно, в каких частях растения образуется цито- кинин: наиболее вероятно, что в корне или в районе быстрого растяжения клеток ниже верхушки побега. Известно, что и развивающийся плод становится местом образования цитокинина, по крайней мере на самых ранних стадиях роста.
Как действуют цитокинины?
Со времени выделения первых цитокининов из препаратов нуклеиновых кислот физиологи растений предполагали, что эти гормоны могут каким-то образом действовать совместно с нуклеиновыми кислотами. Молекулы транспортной РНК (тРНК) содержат ряд необычных оснований (см. рис. 8-8). В некоторых случаях природный цитокинин ¡6Аdе (6N-изопентиладенин), представляющий собой необычное основание, включается в молекулу тРНК. Например, ¡6Аdе обнаружен в молекулах сериновой и тирозиновой тРНК вблизи антикодона. Однако до сих пор неизвестно, связаны ли его присутствие и положение в молекуле тРНК со стимуляцией деления клеток. Известно, что конечное действие цитокинина вызывает изменение экспрессии гена, возможно, на транскрипционном уровне контроля.
Этилен
Открытие ауксина за несколько лет более или менее непосредственно привело к выделению кинетина и выяснению действия цитокининов на рост и развитие растений. С другой стороны, действие этилена на растения было известно задолго до того, как была открыта его связь с ауксином; следовательно, задолго до этого он рассматривался как растительный гормон.
«Ботаническая» история этилена, простого углеводорода (Н2Сг=СН2), отсылает нас к XIX в., когда городские улицы освещались лампами, в которых горел светильный газ. Немецкие ученые заметили, что утечка светильного газа из газопровода вызывает опадение листьев у деревьев, растущих вдоль теневой стороны городских улиц. Когда газ стал более интенсивно использоваться для освещения улиц, об этом явлении сообщили и многие исследователи других стран.
В 1901 г. аспирант Ботанического института Петербургского университета Дмитрий Нелюбов показал, что активным компонентом светильного газа является этилен. Нелюбов заметил, что, если подвергнуть проростки гороха действию светильного газа, они начинают расти в горизонтальном направлении. Из всех компонентов светильного газа данный эффект вызывал только этилен, при этом он был активен уже при таких низких концентрациях в воздухе, как 0,06 млн-1. Выводы Нелюбова были подтверждены другими исследователями, и теперь известно, что этилен оказывает большое влияние на многие, если не на все, процессы роста, развития и старения растений.
Несмотря на то что этилен при физиологических условиях температуры и давления представляет собой газ, до некоторой степени он растворяется в цитоплазме. Этилен рассматривается как растительный гормон, во-первых, потому, что является естественным продуктом метаболизма, а во- вторых, потому, что взаимодействует с другими растительными гормонами в следовых количествах. В частности, действие этилена может наблюдаться во время критических периодов — созревания плодов, опадения плодов и листьев, старения — в жизненном цикле растения.
Сейчас этилен — это самый легкий для определения гормон. В газообразном виде он выделяется тканями и поэтому для проведения анализа не требует экстракции или очистки с помощью газовой хроматографии. Биосинтез этилена начинается с реакции взаимодействия аминокислоты метионина и АТР, в результате которой образуется S-аденозилметионин, сокращенно SAM (рис. 24-9). Затем SAM расщепляется на два соединения, одно из которых содержит кольцо, состоящее из трех атомов углерода, и называется 1-аминоцикло- пропан-1-карбоновой кислотой (АЦПК). Позже АЦПК превращается в этилен, СО2 и аммиак при помощи ферментов тонопласта. По-видимому, реакция образования АЦПК проходит более эффективно при высоких концентрациях ауксина, загрязнении воздуха и ранении растений.
Рис. 24-9. Метионин служит исходным веществом для синтеза этилена во всех тканях высших растений. 1 -Аминоциклопропан-1 -карбоновая кислота является непосредственным предшественником этилена. Показана схема биосинтеза этилена

Этилен и созревание плодов
По мере созревания в плоде происходит ряд изменений. У мясистых плодов разрушается хлорофилл и проявляются другие пигменты, изменяющие окраску плода. Одновременно мясистая часть плода становится мягкой. Эти процессы происходят в результате ферментативного разрушения пектина — основного компонента срединной пластинки клеточной оболочки. Когда срединная пластинка размягчается, клетки могут перемещаться относительно друг друга. В это же время крахмал, органические кислоты или, как в случае авокадо (Persea americana), масла превращаются в сахара. В результате этих изменений плоды становятся заметными, вкусными и привлекают животных, которые, поедая их, рассеивают семена.
Во время созревания многих плодов, например, томатов, авокадо, яблок и груш, активизируется клеточное дыхание, что сопровождается усиленным поглощением кислорода. Эту фазу называют климактерической, а сами плоды — климактерическими (плоды, у которых обнаруживается равномерное снижение дыхания, или постепенное созревание, например, цитрусовые, виноград и земляника, называют неклимактерическими плодами). Связь между климактерическими и другими процессами при созревании плодов неизвестна, но созревание плодов может задерживаться путем подавления климактерической активизации дыхания. Например, холод подавляет его, и у некоторых плодов, помещенных в условия низких температур, не происходит климактерического усиления дыхания. Плоды могут очень долгое время храниться в вакууме. В таких условиях количество доступного кислорода минимально, что способствует подавлению клеточного дыхания, и этилен, ускоряющий наступление климактерического подъема дыхания, удерживается на низком уровне. После климактерической фазы наступает старение, и плод легко поражается грибами и другими микроорганизмами.
В начале 1900-х гг. многие садоводы ввели в практику улучшения окраски и увеличения сахаристости плодов цитрусовых «подлечивание» их в помещениях с керосиновой печкой. (Задолго до этого китайцы ускоряли созревание плодов, окуривая их ладаном.) Предполагалось, что созреванию плодов способствует тепло печки. Однако честолюбивые садоводы, установившие более современное оборудование, нашли, к своему огорчению, что это не так. Как показали последующие эксперименты, на самом деле действующими агентами являются неполные продукты сгорания керосина. Самым активным продуктом сгорания оказался этилен. Даже такое незначительное количество этилена в воздухе, как 1 млн-1, ускоряло наступление климактерического подъема дыхания.
Еще в 1910 г. сообщалось, что газы, выделяемые апельсинами, ускоряют созревание бананов, однако только через 25 лет был идентифицирован этилен как естественный продукт, выделяемый многими плодами и растительными тканями. Количество этилена, образующегося в растении, невелико, и понадобилось разработать новые и очень чувствительные методы, позволившие доказать, что образование этилена отмечается еще до начала климактерического подъема дыхания, во время которого обнаруживаются наибольшие количества этилена. С этих пор этилен стал рассматриваться в качестве естественного регулятора роста растений. Он был найден в плодах (во всех изученных типах), цветках, листьях, облиственных стеблях и корнях многих видов растений, а также у некоторых видов грибов.
Действие этилена на созревание плодов имеет практическое значение. Чаще всего он используется для ускорения созревания томатов, которые собирают зелеными, хранят в отсутствие этилена и обрабатывают им непосредственно перед продажей. Этилен используется и для ускорения созревания орехов и винограда.
Этилен и опадение частей растения
Этилен способствует опадению листьев, цветков и плодов у различных растений. В листьях этилен, возможно, активизирует ферменты, вызывающие разрушение клеточных оболочек, связанное с опадением. Этилен применяют на практике для более легкого отделения плодов вишни, ежевики, винограда и черники, позволяя, таким образом, провести механическую уборку урожая. Он используется и для прореживания плодовой завязи у персиков и слив в больших фруктовых садах.
Этилен и детерминация пола цветков
Этилен, по-видимому, играет большую роль в детерминации пола цветков у однодомных растений (тех, у которых мужские и женские цветки развиваются на одном растении). Например, у тыквы (сем. Cucurbitaceae) высокий уровень гиббереллинов способствует образованию мужских цветков, а обработка этиленом — женских. В работе с огурцами (Сиcumis sativus) было обнаружено, что женские почки выделяют большие количества этилена, чем мужские. Кроме того, огурцы, выращиваемые в условиях короткого дня, способствующего образованию женских цветков, выделяют больше этилена, чем огурцы, выращиваемые в условиях длинного дня. Следовательно, у тыквенных этилен, по-видимому, принимает участие в детерминации пола и увеличивает количество женских цветков.
Этилен и ауксин
У ряда растений ауксин в определенных концентрациях вызывает усиленное образование этилена. Полагают, что те воздействия на плоды и цветки, которые приписываются обычно ауксину, на самом деле связаны с влиянием ауксина на образование этилена.
Форма клетки и ее размеры частично контролируются совместным действием ауксина и этилена, которые влияют на растяжение клетки противоположным образом. К примеру, у гороха огородного (Pisum sativum) концентрации ИУК, которые приводят к отложению поперечно ориентированных микрофибрилл клеточной оболочки, вызывают удлинение стебля, но задерживают его рост в толщину. Напротив, более высокие концентрации ИУК, которые индуцируют более интенсивный синтез этилена и приводят к отложению продольно ориентированных микрофибрилл, увеличивают радиальный рост и разрастание стебля в толщину.
Важно понять, что окончательная форма и размеры клеток, подвергшихся воздействию этилена, являются результатом его взаимодействия не только с ауксином, но также с гибберелловой кислотой и с цитокинином. Более того, нормальный рост и развитие требуют взаимодействий многих
гормонов, в том числе цитокининов, гиббереллинов, абсцизовой кислоты, а также ауксина и этилена.
Абсцизовая кислота
Иногда выживание растения зависит от его способности ограничить свой рост или репродуктивную активность. После открытия гормонов, стимулирующих рост, физиологи предположили, что должны существовать гормоны ингибирующего действия. И наконец в 1949 г. было установлено, что покоящиеся почки ясеня и картофеля содержат большие количества ингибиторов роста, которые блокируют действие И УК на колеоптили овса. Когда покой почек нарушался, содержание ингибитора уменьшалось. Подобные ингибиторы стали называть дорминами.
В течение 1960-х гг. разные исследователи сообщили о нахождении в листьях и плодах веществ, ускоряющих опадение. Одно из них, названное абсцизином, было идентифицировано химически. В 1965 г. была установлена химическая структура одного из дорминов, и оказалось, что он и абсцизин представляют собой одно вещество. Это вещество было названо абсцизовой кислотой.
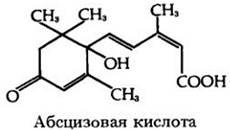
Абсцизовая кислота (АБК) выделена главным образом из оснований завязей плодов. Особенно богатым ею оказался плод хлопчатника (Gossypium). Наибольшее количество абсцизовой кислоты обнаруживается ко времени опадения плодов. Абсцизовая кислота, образующаяся в клетках центральной паренхимы корневого чехлика, участвует в реакции корней на гравитацию (см. с. 117).
Обработка абсцизовой кислотой распускающихся почек превращает их в зимующие почки; при этом листовые примордии преобразуются в почечные чешуи. Ингибирующее действие АБК на почки может быть преодолено применением гиббереллина. Появление альфа-амилазы — фермента, индуцируемого гиббереллином в семенах ячменя (Hordeum vulgare), — задерживается абсцизовой кислотой, которая, по-видимому, подавляет синтез белка в целом. С другой стороны, ауксин, очевидно, взаимодействует с плазматической мембраной и вместе с тем способствует ускорению синтеза специфических белков. Следовательно, ауксин по своему действию является антагонистом абсцизовой кислоты.
Если каплю абсцизовой кислоты нанести на лист, обработанная поверхность быстро желтеет, в то время как остальная часть остается зеленой. (Цитокинины, как мы знаем, обладают противоположным эффектом.) Пока не известно, имеет ли здесь место прямое или опосредованное действие.
В настоящее время абсцизовая кислота не имеет практического применения, возможно, потому, что ее физиология и биохимия изучены недостаточно. В будущем она может приобрести огромное значение для сельского хозяйства засушливых районов. Есть основания предполагать, что устойчивость некоторых растений к стрессовым условиям, таким, как засуха, непосредственно связана с их способностью вырабатывать абсцизовую кислоту. Более того, известно, что абсцизовая кислота способствует закрыванию устьиц некоторых растений, предотвращая тем самым потерю воды листьями и понижая общую потребность растения в воде. Если генетикам удастся включить гены, ответственные за синтез абсцизовой кислоты, в генотип нужных растений, то появится возможность вывести новые сельскохозяйственные сорта, приспособленные к росту в засушливых районах.
Гибберелины
В отличие от других гормонов открытие гиббереллинов не связано с открытием ауксина. Действительно, их изучение началось совершенно независимо от работ с ауксином.
В 1926 г., т. е. в том же году, когда Вент первым выполнил свои эксперименты с агаровыми блоками, Куросава в Японии изучал болезнь риса (Oryza sativa), известную под названием «болезнь бешеных проростков», при которой растения быстро росли, становились веретеновидными, бледно- окрашенными, слабыми и в конце концов полегали. Куросава показал, что болезнь вызывается химическим веществом, выделяемым грибом Gibberella fujikuroi, который паразитирует на проростках риса. По родовому названию этого гриба данное химическое вещество было названо гиббереллином.
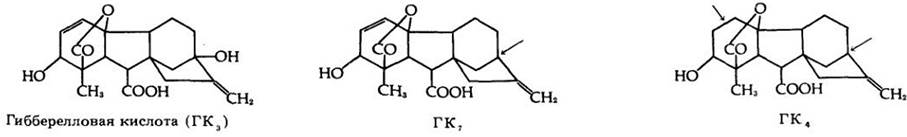
Гиббереллин был выделен и идентифицирован химически японскими биохимиками в 1930-х гт., но в течение нескольких десятилетий он привлекал к себе мало внимания. Затем в 1956 г. гиббереллин успешно выделили не только из гриба, но и из растения (семян Phaseolus vulgaris). После этого были очищены гиббереллины многих видов растений, и, вероятно, они имеются у всех растений. Гиббереллины содержатся во всех частях растения в различных концентрациях, но самая высокая их концентрация найдена в незрелых семенах. К настоящему времени выделено и идентифицировано более 65 гиббереллинов. Они мало различаются по структуре (рис. 24-10) и биологической активности. Лучше всего изучена гибберелловая кислота (ГК3), которая образуется грибом Gibberella fujikuroi.
Рис. 24-10. Три природных гиббереллина (в настоящее время известно более 65). Гибберелловая кислота (ГК3) встречается в значительных концентрациях у грибов и проявляет наибольшую биологическую активность во многих экспериментах. Стрелки указывают небольшие структурные различия гиббереллинов ГК7 и ГК4
У интактных растений гиббереллины оказывают значительное влияние на удлинение стебля. Заметное усиление роста побега — самая общая реакция, отмеченная у высших растений; часто стебли становятся длинными и тонкими, а листья бледнеют. Гиббереллины стимулируют как деление клеток, так и их растяжение и действуют как на листья, так и на стебли.
Гиббереллины и карликовые мутанты
Наиболее явно действие гиббереллинов проявляется при обработке карликовых мутантов, дефектных по одному гену (рис. 24-11). В результате такие растения невозможно отличить от нормальных высоких немутантных растений. У кукурузы, например, идентифицированы четыре типа карликовых мутантов, у каждого из которых нарушены определенные стадии биосинтеза гиббереллина. В настоящее время изучение биохимии гормонов у этих мутантных растений привело к очень важному заключению. Хотя растения кукурузы и содержат девять различных соединений, которые соответствуют по своему действию гиббереллинам (т. с. вызывают ростовые реакции), однако только конечное соединение этого метаболического ряда может оказывать прямое действие. Другие восемь гиббереллинов должны пройти дальнейшие этапы биохимических превращений, прежде чем смогут вызвать ростовую реакцию.
Рис. 24-11. Растение, показанное справа, обрабатывалось гиббереллином; растение слева служило контролем. Опыт проводили на карликовых мутантах обычной фасоли (Phaseolus vulgaris)

Гиббереллины и семена
Семена большинства растений до прорастания находятся в состоянии покоя. У некоторых покой прерывается холодом или светом. Для многих видов, включая салат, табак и дикие овсы, гиббереллины могут заменить требующиеся для прерывания покоя холод или свет и способствовать росту зародыша и появлению проростков. Характерно, что гиббереллины стимулируют удлинение клеток и тем самым способствуют проникновению корня через сдерживающую рост семенную кожуру или оболочку плода. Подобное действие гиббереллина имеет по крайней мере одно практическое применение. Гибберелловая кислота ускоряет прорастание семян и таким образом обеспечивает однородность исходного материала, идущего на изготовление ячменного солода для нужд пивоваренной промышленности.
Гиббереллины и ювенильность
Ювенильные стадии некоторых растений иногда отличаются от взрослых по форме листьев. Среди однолетних двудольных фасоль (Phaseolus), у которой молодые листья являются простыми, а старые — сложными (трехлисточковыми), представляет собой прекрасный пример гетерофилии (см. рис. 19-2). Среди многолетников у многих видов эвкалиптов (Eucalyptus) обнаруживаются поразительные различия между молодыми и старыми листьями (рис. 24-12).
Рис. 24-12. Молодые (А) и старые (Б) листья Eucalyptus globulus, между которыми видны большие различия, встречающиеся у данного вида. Молодые листья более сочные и расположены супротивно. Под верхней эпидермой имеется единственный слой палисадной паренхимы. Старые листья жесткие, располагаются по спирали и свешиваются вниз. У них обе поверхности обращены к свету, и поэтому палисадная паренхима располагается с обеих сторон листовой пластинки

Плющ (Hederá helix), многолетняя лиана, представляет собой другой хорошо изученный пример. Если взрослые растения плюща вьются у стены вашего дома, сравните верхние ветви с нижними. Форма листьев и поведение ветвей окажутся различными. Молодые ветви укореняются быстро, у взрослых ветвей этого не наблюдается. Взрослые ветви цветут, молодые — нет. Если у взрослой ветви отрезать апикальную меристему, то формируются пазушные почки, из которых развиваются новые взрослые ветви. Однако, если такую почку обработать гиббереллином, она развивается в типичную ювенильную ветвь.
Гиббереллины и цветение
Некоторые растения, такие, как капуста (Brassica olerácea var. capitata), морковь (Daucus carota) и двулетняя белена (Hyoscyamus niger), до цветения образуют розетки. (В розетке развиваются листья, но междоузлия не удлиняются.) Цветение таких растений можно вызвать, помещая их в условия длинного дня или на холод (как в случае двулетников); можно и совместить оба фактора. После соответствующего выдерживания стебли удлиняются (данное явление называют стрелкованием), и растение начинает цвести. Применение гиббереллина вызывает стрелкование и цветение растений без соответствующей обработки холодом и воздействия длинного дня (рис. 24-13). Рост стеблей достигается за счет увеличения числа клеточных делений и удлинения клеток, образующихся в результате этих делений. Гиббереллин, таким образом, может быть использован для быстрого получения семян у двулетних растений. Обрабатывая гибберелловой кислотой, например, капусту, семена можно получить лишь за один вегетационный период.
Рис. 24-13. Стрелкование стебля капусты (Brassica olerácea var. capitata), вызванное обработкой гиббереллином. Растение справа обрабатывалось один раз в неделю в течение восьми недель

Гиббереллины, пыльца и развитие плода
Как было показано, гиббереллины стимулируют прорастание пыльцы и рост пыльцевых трубок у ряда растений, в том числе лилии, лобелии, петунии и гороха. Подобно ауксину.
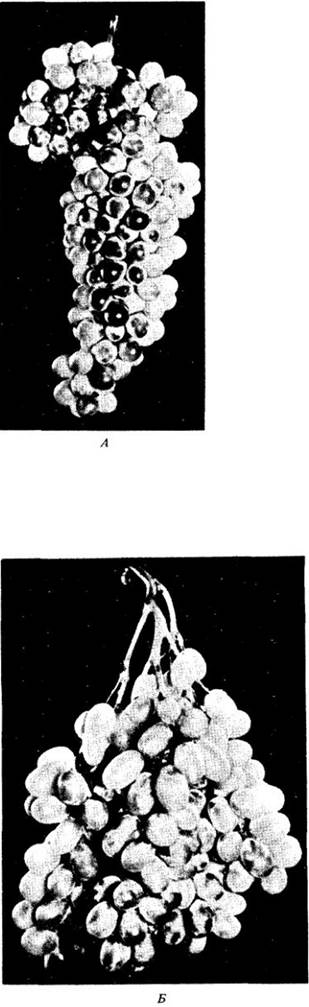
гиббереллины могут вызвать образование партенокарпических плодов у яблони, смородины, огурцов и баклажанов. У мандариновых и персиковых деревьев, а также миндаля гиббереллины могут ускорить развитие плода в тех случаях, когда с помощью ауксина этого сделать не удается. Однако гиббереллины применяются главным образом при выращивании столового винограда. В США большие количества гибберелловой кислоты ежегодно расходуются при выращивании таких сортов винограда (Vitis vinifera), как ThompsonSeedless («Томпсон бессемянный»). Обработка приводит к образованию более крупных ягод и рыхлых кистей винограда (рис. 24-14).
Рис. 24-14. Действие гибберелловой кислоты на рост винограда сорта Thompson Seedless. А. Необработанная гроздь, оставшаяся тонкой и плотной. Б. Обработанная гибберелловой кислотой кисть стала рыхлой
Как действуют гиббереллины?
Наиболее важные исследования механизма действия гиббереллина были проведены одновременно учеными Японии, Австралии и США. Детальное изучение последовательности процессов, происходящих при прорастании семени ячменя и на ранних стадиях развития зародыша, позволило выявить ключевую роль, которую играют гиббереллины в этой цепи событий. Кроме того, эти исследования наилучшим образом продемонстрировали, как гормоны «соединяют» в единое целое биохимию и физиологию различных тканей растения.
В семенах ячменя (Hordeum vulgare) и других злаков имеется специализированный слой клеток эндосперма, так называемый алейроновый слой (см. рис. 19-4), расположенный под семенной кожурой. Клетки алейронового слоя богаты белком. Когда семена, набухнув, начинают прорастать, зародыш выделяет гиббереллины. В ответ на их действие клетки алейронового слоя синтезируют гидролитические ферменты, из которых главным является альфа-амилаза, превращающая крахмал в сахара (рис. 24-15). Ферменты разрушают запасные питательные вещества эндосперма до сахаров и аминокислот, которые поглощаются щитком и затем транспортируются к растущим частям зародыша (рис. 24-16). Таким образом зародыш получает необходимые для роста вещества в тот самый момент, когда они ему требуются.
Рис. 24-15. Обработка гиббереллином (ГКз) способствует высвобождению сахара из эндосперма только при наличии алейронового слоя. Действительно, алейроновый слой является источником альфа-амилазы, разрушающей запасной крахмал эндосперма
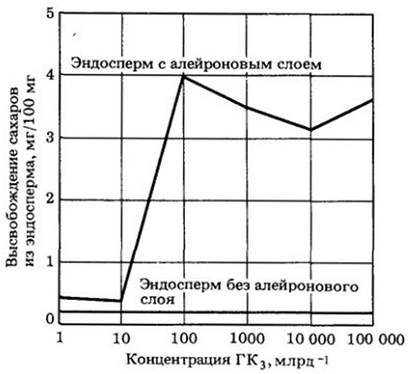
Рис. 24-16. Действие гиббереллина в семенах ячменя. А. Гибберелловая кислота (ГК), образующаяся в зародыше, мигрирует в алейроновый слой, стимулируя синтез гидролитических ферментов. Эти ферменты расщепляют запасной крахмал эндосперма до растворимых и способных к диффузии сахаров, и аминокислот. Затем сахара и аминокислоты поглощаются щитком (семядолей) и передвигаются к побегу и корню, способствуя их росту. Б. Каждое из этих трех семян было разрезано пополам и зародыш удален. За 48 ч до того, как была сделана фотография, верхнее семя слева было обработано простой водой. Семя в центре было обработано раствором гиббереллина концентрацией 1 млрд-1, а семя внизу справа обрабатывалось гиббереллином из расчета 100 млрд-1. Видно, как в обработанных семенах началось разрушение запасного крахмала

Исследователи полагают, что гиббереллины активируют некоторые гены, направляющие синтез специфической мРНК и ферментов. Однако еще не доказано, что гиббереллины действуют непосредственно на гены, хотя отмечено, что имеет место синтез как РНК, так и ферментов. Независимо от деталей механизма действия гиббереллина в алейроновых клетках ясно, что они образуют высокодифференцированную ткань, приспособленную к удовлетворению (при посредничестве гиббереллинов) потребностей растущего зародыша. Остается неизвестным, одинаков ли механизм действия гиббереллинов в семенах и других частях растения.
Заключение
Гормоны представляют собой важные химические регуляторы роста животных и растений, которые образуются в одних тканях организма и транспортируются в другие, где проявляется их физиологическое действие. Гормоны биологически активны в чрезвычайно малых количествах.
Природный гормон ауксин образуется в апикальной меристеме побегов и кончиках колеоптилей. Ауксин перемещается только по направлению к основанию растения, где регулирует удлинение побега и колеоптиля. способствуя главным образом растяжению клеток. Исследования показывают, что его действие на растяжение клетки достигается косвенным путем благодаря ослаблению связей целлюлозных фибрилл клеточной оболочки, что позволяет клетке растягиваться. Ауксин играет роль и в дифференцировке проводящих тканей и вызывает деление клеток камбия. Он тормозит рост боковых почек, поддерживая тем самым апикальное доминирование. То же количество ауксина, которое способствует росту стебля, задерживает рост главной корневой системы. Ауксин ускоряет формирование придаточных корней у черенков и задерживает опадение листьев, цветков и плодов. В плодах ауксин, выделяемый семенами или пыльцевыми трубками, стимулирует рост стенки завязи. Предполагают, что его способность оказывать такие разнообразные воздействия зависит от неодинаковой чувствительности к нему тканей-мишеней.
Цитокинины, относящиеся ко второму классу ростовых гормонов, были открыты благодаря их способности ускорять деление клеток и формирование почек в культуре растительных тканей. По своей структуре они близки к некоторым компонентам нуклеиновых кислот. Цитокинины могут действовать совместно с ауксинами, вызывая деление клеток invitro. В культуре клеток сердцевины табака высокая концентрация ауксина ускоряет формирование корня, в то время как высокая концентрация цитокинина ускоряет формирование почек. В интактных растениях цитокинины ускоряют рост боковых почек, действуя как антагонисты ауксина. Цитокинины предотвращают старение листьев, стимулируя синтез белка.
Этилен представляет собой газ, образующийся в результате неполного сгорания углеводородов. В то же время он относится к естественным регуляторам роста и вызывает ряд отчетливых физиологических реакций, например, созревание плодов и опадение частей растения.
Абсцизовая кислота, найденная в покоящихся почках и плодах, — это гормон, тормозящий рост. В ряде экспериментов абсцизовая кислота вызывает действие, противоположное всем трем стимулирующим рост гормонам.
Таблица 24-1. Растительные гормоны (фитогормоны) и их действие
Название Ауксин |
Химическая природа Индолил-3-уксусная кислота, фенилуксусная кислота |
Физиологическое действие и роль Апикальное доминирование, гравитропизм и фототропизм, дифференцировка проводящих тканей; тормозит опадение, стимулирует синтез этилена, подавляет или стимулирует (у ананаса) цветение, стимулирует развитие плода (партенокарпические плоды), вызывает образование корней на черенках |
Цитокинин |
N6-производные аденина, соединения фенилмочевины |
Апикальное доминирование, рост побега, развитие плода; задерживает старение листьев |
Этилен |
СН2 = СН2 |
Созревание плодов (особенно климактерических плодов, таких, как яблоки, бананы, авокадо), старение листьев и цветков, опадение частей растения |
Гиббереллин |
Гибберелловая кислота (ГК3), ГК1 |
Стимулирование цветения у длиннодневных растений и двулетников, удлинение побегов; регулирует образование ферментов в семенах злаков |
Абсцизовая кислота |
АБК |
Закрывание устьиц; возможно, необходима для опадения частей растений и сохранения покоящегося состояния у некоторых видов |
Гиббереллины впервые были выделены из паразитических грибов, нарушающих нормальный рост сеянцев риса. Впоследствии обнаружилось, что они являются природными ростовыми гормонами многих растений. Обработка гиббереллинами карликовых мутантов растений восстанавливает их нормальный рост, а у растений с розеточной формой роста вызывает стрелкование. Гиббереллины стимулируют прорастание семян злаков. В семенах ячменя зародыш выделяет гиббереллины, которые способствуют синтезу в алейроновых клетках эндосперма некоторых ферментов, в том числе альфа-амилазы, переводящей крахмал в сахара. Сахар используется зародышем и способствует прорастанию семян.
Действие растительных гормонов в значительной степени зависит от свойств тканей-мишеней и окружающей их химической среды. Основные гормоны растений и контролируемые ими физиологические процессы приведены в табл. 24-1.
Приложение. Растительные гормоны, культура ткани и биотехнология
Успехи в исследовании гормонов и биохимии ДНК сделали возможным вмешательство в генетику растений. Термин биотехнология используется для описания практической реализации этой возможности. Среди важнейших методов биотехнологии следует отметить культуру ткани, которая может быть осуществима только благодаря разработанным теперь представлениям о гормонах, регулирующих развитие. В идеальных случаях культура ткани используется для получения целого растения из одиночных, генетически измененных клеток. Кроме того, культура ткани может быть использована для получения в течение короткого периода времени многих идентичных копий (клонов) растений.
В настоящее время культура ткани широко используется для размножения растений путем клонирования, поскольку получаемые при этом индивиды генетически идентичны. В основном в культуре ткани используются сейчас три различных способа размножения, причем каждый имеет свои достижения в управлении развитием, которое может осуществляться при помощи растительных гормонов.
1. Регенерация из каллуса и(или) протопластов. Растения, которые могут быть размножены этим способом, особенно ценны для биотехнологии. Основанная на исследованиях, проведенных Скугом и его сотрудниками, данная методика предусматривает использование цитокининов для образования побегов из неорганизованной массы паренхимных клеток, называемой каллусом. Затем побеги обрабатывают ауксином для образования корней, и после этого развивающиеся растения могут быть перенесены в почву. Регенерация из каллуса была впервые проведена на табаке, а затем и на других растениях семейства пасленовых (Solanaceae), таких, как картофель и петуния. Хотя некоторые растения и других семейств (например, подсолнечник, львиный зев и горчица) могут быть регенерированы из каллуса, данный метод не получил широкого распространения.
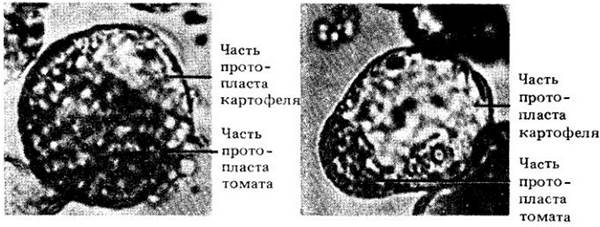
Слившиеся протопласты томата и картофеля. Часть протопласта, принадлежащую томату, можно идентифицировать по зеленым хлоропластам, которые присутствуют в изолированных протопластах мезофилла томата. Часть, принадлежащая картофелю, бесцветна, поскольку изолированные протопласты картофеля содержат только пропластиды. (Эксперименты G. Melchers. М. D. Sacristán и А. А. Holder. Профессор Melchers работает в Биологическом институте им. Макса Планка, Тюбинген, ФРГ.)
Теоретически любая растительная клетка, если она не лишена ядра или не заключена в твердую лигнифицированную оболочку, потенциально способна развиться в организм, из которого произошла, и, как говорят, обладает тотипотентностью. Совокупности сходных клеток образуют ткани; ткани и системы тканей образуют
органы, которые, располагаясь определенным образом в пространстве, формируют организм. Растения могут быть регенерированы in vitro многими путями: из эксплантатов органов (кончиков корня и верхушек побега, боковых и придаточных почек, примордиев листа, развивающихся зародышей, почечных чешуи и т. д.); эксплантатов тканей (сердцевины, первичной коры, эпидермы, флоэмы, нуцеллуса); клеток (паренхимы, колленхимы, одно- или двуядерных пыльцевых зерен) и протопластов. Приведенная здесь схема иллюстрирует несколько путей, которыми может осуществляться регенерация целого растения

Наиболее изящное использование культуры ткани для биотехнологии в настоящее время связано с регенерацией растения из протопластов — клеток, у которых ферментативным путем удалены клеточные оболочки. Разработаны два способа использования протопластов в генетике растений. Первый предполагает слияние протопластов двух различных растений с образованием гибридных клеток. Сначала при помощи данного метода был получен гибрид табака, затем и межвидовые гибриды петунии, моркови, картофеля и межродовые гибриды картофеля и томата — растений семейства пасленовых.
Во втором случае применяют Ti-плазмиды корончатых галлов, образуемых Agrobacterium tumefaciens, или какой- либо другой метод инъекции ДНК для введения в протопласты специфических генов. Далее культуру ткани используют для регенерации растений из отдельных протопластов. Это направление генной инженерии уже нашло применение для получения устойчивых к гербицидам растений табака и подсолнечника (см. гл. 30).
2. Соматический эмбриогенез. В эксперименте, проведенном Ф. Стьюардом из Корнеллского университета, культура ткани была использована для получения зародышей моркови из единичных клеток корня. Эта процедура, названная соматическим эмбриогенезом, поскольку в этом случае зародыш развивается из вегетативных (т. е. соматических) клеток, иллюстрирует поразительную тотипотентность растительных клеток. Тотипотентность — это неотъемлемая способность отдельной клетки обеспечивать генетическую программу, необходимую для развития целого организма. Соматический эмбриогенез открывает с этой точки зрения большие возможности для клонального размножения растений и уже успешно применяется для кукурузы, пшеницы и сорго.
3. Размножение побегов. Эта методика основывается на свойствах цитокинина усиливать рост побега и противодействовать ауксину в регуляции апикального доминирования. Верхушки растущих побегов помещают в культуральную среду с достаточно высокой концентрацией цитокинина не только для поддержания роста побега, но и для развития боковых почек в пазухах листьев. Таким образом можно размножить побег, культивируя его в течение нескольких недель in vitro. После рассаживания отдельные побеги могут быть использованы для дальнейшего размножения или укоренены после обработки ауксином. Для разработки этого метода особенно много сделал Тосио Мурасиге из Калифорнийского университета в Риверсайде. Метод обычно используют для размножения папоротников, орхидей и древесных растений семейства Ericaceae (семейство вересковых, включающее рододендроны, азалии, кальмии) и Rosaceae (семейство розовых, к которому относятся яблони, розы и земляника).