МОРФОЛОГИЯ РАСТЕНИЙ - Т. А. Сауткина - 2012
ГЛАВА 7. ЦВЕТОК КАК ОСОБЫЙ РЕПРОДУКТИВНЫЙ ОРГАН ПОКРЫТОСЕМЕННЫХ РАСТЕНИЙ
7.7. Формирование пестика в онтогенезе. Характеристика процессов, происходящих в семяпочке
В процессе онтогенеза будущий пестик или формируется в виде бугорка меристематической ткани с последующим изменением формы и срастанием краев плодолистика, или закладывается в виде кольцевого валика. В ходе дальнейшего развития постепенно оформляется пестик, состоящий из завязи, стилодия и рыльца. Стилодиев может быть несколько, или они срастаются и образуют столбик. Иногда столбик отсутствует (мак — Рараvег) и рыльце оказывается сидячим. По анатомическому строению завязь пестика имеет явное сходство с листом. Снаружи она покрыта эпидермисом, под которым расположен мезофилл с хорошо развитой проводящей системой, полость завязи (семенная камера) также выстлана эпидермисом.
7.7.1. Заложение и развитие семяпочки
На определенном этапе развития на внутренней стенке завязи начинают формироваться семяпочки (семязачатки) (см. рис. 156).
Каждая семяпочка закладывается субэпидермально (под эпидермисом) в виде бугорка меристематической ткани. По мере увеличения меристематического бугорка начинает оформляться центральная часть семяпочки — нуцеллус, который и является макроспорангием (мегаспорангием). Нуцеллус может быть массивным или слабо выраженным. На ранних этапах развития семяпочки намечается ее поляризация: в ней четко выражены халазальный и микропилярный полюсы. Название каждый полюс получил в соответствии с наличием определенных частей семяпочки — халазы и микропиле, занимающих диаметрально противоположное положение.
Халаза семяпочки имеет большое значение как трофическая структура. В области халазы семяпочка прикрепляется к стенке завязи при помощи семяносца, или фуникулуса. К халазе подходит сосудисто-волокнистый пучок, который проходит через фуникулус, разветвляется в основании нуцеллуса и снабжает водой с растворенными в ней минеральными веществами клетки развивающейся семяпочки.
От халазы как боковые выросты начинают формироваться интегументы — покровы семяпочки (рис. 178). Количество интегументов у представителей различных семейств покрытосеменных различно. У свободнолепестных развивается два интегумента — наружный и внутренний. У спайнолепестных формируется один интегумент, у некоторых растений-паразитов интегументы вообще отсутствуют. Интегументы выполняют защитную функцию. По мере увеличения нуцеллуса интегументы обрастают его со всех сторон, но на верхушке семяпочки не срастаются и образуют микропиле (пыльцевход). В образовании микропиле чаще всего принимают участие оба интегумента, тогда внутренний интегумент образует эндостом, а наружный — экзостом. Иногда микропиле образуется каким-нибудь одним интегументом. В редких случаях у некоторых апомиктичных растений микропиле вообще отсутствует.
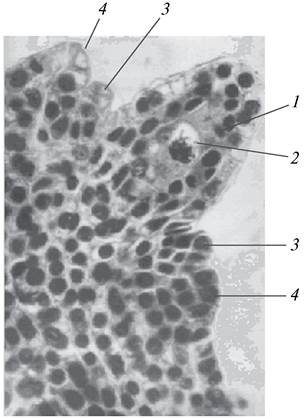
Рис. 178. Макроспороцит (мегаспороцит) бобов обыкновенных (Vicia faba) в нуцеллусе семяпочки: 1—нуцеллус; 2—макроспороцит; 3—формирующийся внутренний интегумент; 4—формирующийся наружный интегумент
7.7.2. Заложение женского археспория. Процесс макроспорогенеза (мегаспорогенеза)
На ранних этапах онтогенеза, когда интегументы достигают приблизительно половины высоты нуцеллуса, в его микропилярной части начинает субэпидермально выделяться одна (сем. Толстянковые — Crassulaceae, Ароидные — Aгасеае), реже несколько клеток (сем. Рясковые — Lemnaceae), которые становятся первичными археспориальными клетками (клетками женского археспория). Клетки первичного археспория крупнее остальных клеток нуцеллуса, имеют невакуолизированную цитоплазму и крупное ядро. У некоторых растений они делятся митотически и образуют кроющие клетки и клетки вторичного археспория (спорогенные клетки, материнские клетки макроспороцитов) (сем. Розоцветные), у других же (сем. Водокрасовые — Hydгоchагiасеае, Бальзаминовые — Ваl- saminaceae, Бобовые — Fabaceae) без деления становятся макроспороцитами (мегаспороцитами) (см. рис. 178). Различия в строении женского археспория тесно связано со строением нуцеллуса. Крассинуцеллятные семяпочки обычно имеют кроющие клетки, тенуинуцеллятные семяпочки их не имеют, хотя есть целый ряд исключений. Кроющие клетки продвигают мегаспороциты в глубь нуцеллуса. Придать разным типам женского археспория статус филогенетического признака не представляется возможным, так как одни и те же типы археспория встречаются у представителей как примитивных, так и высокоорганизованных семейств. Например, многоклеточный археспорий, который считается примитивным типом женского археспория, обнаружен у таких примитивных семейств, как Лютиковые, Розоцветные, а также у эволюционно продвинутых семейств — Сложноцветные, Зонтичные.
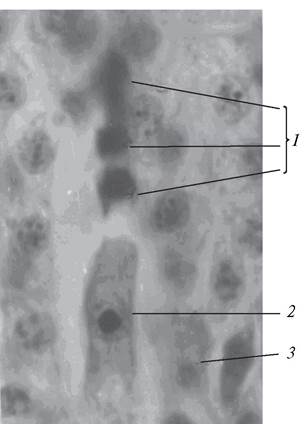
На определенном этапе развития в нуцеллусе семяпочки происходит процесс макроспорогенеза (мегаспорогенеза). При этом ядро макроспороцита претерпевает редукционное деление (мейоз) с последующим образованием клеточных перегородок, в результате чего формируется тетрада макроспор (мегаспор). Клеточные перегородки макроспор закладываются по симультанному типу. Макроспоры в тетраде обычно расположены линейно (рис. 179). Редко встречается иное расположение макроспор в тетраде. У представителей семейства Кипрейные (Onagraceae) довольно часто образуются Т- образные тетрады, у семейства Перечные (Piperaceae) — тетраэдрические.
Рис. 179. Тетрада макроспор (мегаспор): 1—три дегенерирующие макроспоры; 2—макроспора, из которой будет формироваться женский гаметофит (зародышевый мешок); 3—клетки нуцеллуса
Каждая макроспора одета каллозной оболочкой и представляет собой типичную гаплоидную клетку.
Процесс макроспорогенеза начинается позже микроспорогенеза, но протекает значительно быстрее, что затрудняет его изучение. Данные, касающиеся цитохимических и субмикроскопических особенностей археспориальных клеток и макроспор, фрагментарные.
7.7.3. Развитие женского гаметофита (зародышевого мешка). Строение типичного зародышевого мешка
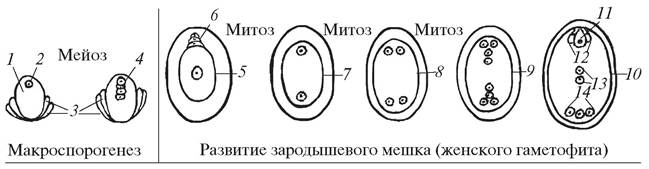
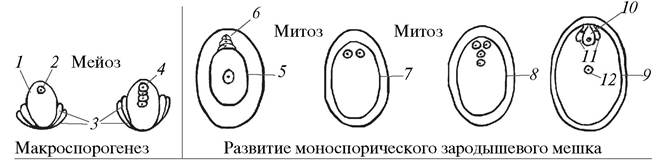
После макроспорогенеза макроспоры впадают в период кратковременно го покоя, а затем в нуцеллусе семяпочки начинается новый процесс — образование женского гаметофита (зародышевого мешка). У большинства покрытосеменных растений из четырех возникших макроспор только одна, чаще всего халазальная (ближе расположенная к халазальному полюсу), оказывается функционально способной, она может образовать женский гаметофит. Зародышевый мешок, развивающийся из одной мегаспоры, называется мо- носпорическим, или типичным зародышевым мешком, потому что он выявлен у большинства изученных покрытосеменных растений. Так как он впервые был описан у Polygonumdivaricatum — представителя семейства Гречишные (Polygonaceae), его называют также зародышевым мешком Polygonum-типа (рис. 180).
Рис. 180. Образование макроспор и развитие типичного зародышевого мешка Polygonum - типа: 1—нуцеллус семяпочки (макроспорангий); 2—макроспороцит; 3— интегументы семяпочки; 4—тетрада макроспор; 5—разрастающаяся халазальная макроспора (одноядерный зародышевый мешок); 6—дегенерирующие микропилярные макроспоры; 7—двуядерный зародышевый мешок; 8—четырехъядерный зародышевый мешок; 9—молодой восьмиядерный зародышевый мешок; 10—зрелый зародышевый мешок; 11—яйцеклетка; 12—синергиды; 13—полярные ядра; 14—антиподы
Развитие зародышевого мешка происходит благодаря серии митотических делений ядра мегаспоры и его производных. Зародышевый мешок развивается как ценоцитная структура, т. е. при развитии зародышевого мешка образование дочерних ядер не сопровождается образованием клеточных перегородок между ними.
Началом образования типичного зародышевого мешка является разрастание халазальной макроспоры, при этом три другие макроспоры начинают сжиматься и впоследствии дегенерируют. Вследствие разрастания макроспоры, прилегающие к ней клетки нуцеллуса, также разрушаются, и их содержи мое используется на питание развивающегося женского гаметофита.
Зародышевый мешок проходит в своем развитии ряд стадий, и каждая из них носит название, отражающее число ядер в развивающемся зародышевом мешке и степень их дифференцировки.
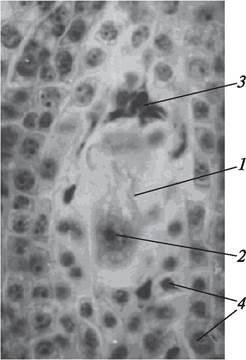
Начало образования, когда в развивающемся зародышевом мешке (фактически в разрастающейся макроспоре) имеется только одно ядро, называет - ся стадией одноядерного зародышевого мешка (рис. 181). На этой стадии ядро занимает центральное положение, окружено слоем цитоплазмы, который соединяется с цитоплазмой, расположенной на периферии.
Рис. 181. Одноядерный зародышевый мешок бобов обыкновенных (Vicia faba): 1—халазальная макроспора, из которой развивается зародышевый мешок; 2—ядро разрастающейся макроспоры; 3—дегенерирующие микропилярные макроспоры; 4—клетки нуцеллуса
Первое митотическое деление ядра макроспоры завершается образованием двух ядер, расположенных в центре развивающейся макроспоры. Эта стадия называется стадией молодого двуядерного зародышевого мешка. Вследствие образования крупной вакуоли между возникшими ядрами они расходятся к противоположным полюсам, и зародышевый мешок становится двуядерным (рис. 182). После того как на халазальном и микропилярном полюсах оказывается по одному ядру, деление ядер в развивающемся зародышевом мешке происходит строго синхронно.
Рис. 182. Этапы развития зародышевого мешка Polygonum-muna: А—«молодой» двуядерный зародышевый мешок; Б—двуядерный зародышевый мешок: 1—ядра в экваториальной зоне развивающегося зародышевого мешка; 2—клетки нуцеллуса; 3—разрушающиеся клетки нуцеллуса, прилегающие к развивающемуся зародышевому мешку; 4—ядро на микропилярном полюсе; 5—ядро на халазальном полюсе
Следующее, второе по счету, митотическое деление завершается образованием на каждом полюсе двух ядер, и зародышевый мешок становится четырехъядерным. И, наконец, наступает третий, последний, митоз, после которого на каждом полюсе зародышевого мешка оказывается по четыре ядра. Вокруг каждой группы из четырех ядер концентрируется цитоплазма, обе группы соединены между собой тяжами цитоплазмы, которые расположены по периферии. Зародышевый мешок, в котором имеется восемь недифференцированных ядер, называется молодым восьмиядерным зародышевым мешком. Он представляет собой довольно крупную полость в тканях нуцеллуса, ограниченную его разрушающимися клетками. Имеет ли зародышевый мешок собственную оболочку — не ясно. Одни исследователи указывают на наличие вокруг зародышевого мешка цитоплазматической оболочки, по мнению других, она отсутствует и зрелый зародышевый мешок ограничен только прилегающими клетками нуцеллуса.
Несмотря на то что ядра в зародышевом мешке образуются в результате митоза и, казалось бы, должны обладать одинаковыми свойствами, они на самом деле оказываются разнокачественными, о чем свидетельствует не только их морфология, но и функциональные особенности. Предполагают, что формированию такой разнокачественности способствует характер обменных процессов ядер микропилярного и халазального полюсов с тканями материнского организма.
После завершения митотических делений в полости молодого зародышевого мешка начинается процесс дифференцировки ядер (см. рис. 180). По одному ядру с каждого полюса мигрирует в центральную (экваториальную) зону зародышевого мешка и располагаются они в общей цитоплазме. Эти ядра называются полярными. Термин «полярные ядра» не несет функциональной нагрузки, но он показывает, откуда переместились в центр, в экваториальную зону, эти ядра.
Вокруг трех ядер микропилярного полюса начинают обособляться участки цитоплазмы, в результате чего формируются три клетки, составляющие яйцевой аппарат зародышевого мешка. В состав яйцевого аппарата входит яйцеклетка и две синергиды. Эти клетки хорошо различаются по морфологическим признакам. Яйцеклетка имеет грушевидную форму, ядро занимает апикальную часть яйцеклетки, а в базальной части находится крупная вакуоля. Синергиды расположены по обе стороны яйцеклетки, они имеют вытянутую форму. В отличие от яйцеклетки ядро синергид расположено в базальной части клетки, а небольшая вакуоль — в апикальной. Вопрос об оболочках клеток яйцевого аппарата до настоящего времени не решен.
Вокруг трех ядер, оставшихся на халазальном полюсе, также обособляется цитоплазма и образуются три клетки. Они называются антиподами и характеризуют особенность расположения этой группы — напротив яйцевого аппарата.
Клетки зародышевого мешка имеют четкую функциональную дифференцировку. Яйцеклетка — единственная женская гамета. Синергиды выполняют хемотропическую функцию, они «привлекают» пыльцевую трубку и ориентируют ее положение после вхождения в зародышевый мешок. Иногда синергиды могут образовывать микропилярные гаустории, и тогда они могут выполнять трофическую функцию, так как способствуют поступлению питательных веществ к яйцеклетке. Антиподы — это трофические клетки, которые с помощью гаусторий поставляют питательные вещества из халазальной части семяпочки в зародышевый мешок до того момента, пока не сформируется эндосперм. Полярные ядра после оплодотворения дают начало особой питательной ткани — эндосперму. У одних растений полярные ядра остаются свободными вплоть до слияния со спермием, у других они сливаются до оплодотворения, и образуется центральное ядро зародышевого мешка. Некоторые эмбриологи считают, что вокруг него также обособляется цитоплазма и возникает центральная клетка зародышевого мешка.
Таким образом, женский гаметофит покрытосеменных растений, как и мужской, сильно редуцирован, образуется в тканях материнского организма, никогда его не покидает и развивается за счет питательных веществ материнского растения — спорофита. Сильную редукцию женского гаметофита следует рассматривать как адаптивный процесс, который сокращает время развития полового поколения и ускоряет развитие половых клеток.
Зародышевый мешок, в котором произошла дифференцировка клеток, считается зрелым, он готов к процессу оплодотворения.
7.7.4. Типы развития зародышевых мешков
Приблизительно у 80 % изученных покрытосеменных растений зародышевый мешок развивается по нормальному типу, т. е. представлен зародышевым мешком Роlygonum-munа (см. рис. 180). В то же время у представителей различных семейств покрытосеменных выявлены так называемые зародышевые мешки отклоняющихся типов.
Представление о типах развития зародышевого мешка было окончательно сформулировано профессором И. Д. Романовым. При характеристике типа развития зародышевого мешка учитывается не только число мегаспор, дающих начало зародышевому мешку, но указывается также число делений, которые происходят при его образовании, число полюсов и количество ядер в зародышевом мешке.
Таким образом, зародышевый мешок нормального типа (Polygonum-типа) является моноспорическим, трехмитозным, двухполюсным, восьмиядерным.
Зародышевые мешки отклоняющихся типов существенно отличаются от нормального типа. В зависимости от числа макроспор, образующих зародышевые мешки, они бывают моноспорическими (развиваются из одной мегаспоры), биспорическими (образуются из диады мегаспор) и тетраспорическими (формируются из четырехъядерного ценоцита, т. е. из своеобразной «тетрады мегаспор», в которой не произошло образование клеточных перегородок). Развитие зародышевых мешков отклоняющихся типов происходит за одно (одномитозные зародышевые мешки) или два (двухмитозные зародышевые мешки) митотических деления. Количество ядер в зародышевых мешках отклоняющихся типов варьирует от 4 до 16. В некоторых зародышевых мешках количество ядер может сокращаться за счет полиплоидизации, в таких зародышевых мешках часть ядер являются гаплоидными, а часть — триплоидными.
В зависимости от того, как в онтогенезе распределяются ядра в полости зародышевых мешков отклоняющихся типов, последние могут быть однополюсными, двухполюсными и четырехполюсными.
Из моноспорических зародышевых мешков отклоняющегося типа своеобразно формируется зародышевый мешок Oenothera-тиnа (рис. 183). Этот моноспорический, двухмитозный, однополюсный зародышевый мешок впервые был обнаружен В. Гофмейстером у представителей семейства Кипрейные(Onagraceae). Развитие зародышевого мешка происходит из микропилярной макроспоры. В нем всего четыре ядра: три ядра образуют яйцевой аппарат, а одно ядро находится в центре и является полярным. До настоящего времени этот тип развития не выявлен ни у одного представителя других семейств, поэтому он имеет большое систематическое значение. На основании изучения типа развития зародышевого мешка род Trapa L. — рогульник, который ранее входил в семейство Onagraceae, был исключен из него и переведен в семейство Trapaceae — Рогульниковые. У рогульника зародышевый мешок формируется по Polygonum-типу.
Рис. 183. Образование макроспор и особенности развития зародышевого мешка Oenothera-типа: 1—нуцеллус семяпочки (макроспорангий); 2—макроспороцит; 3— интегументы семяпочки; 4—тетрада макроспор; 5—разрастающаяся халазальная макроспора (одноядерный зародышевый мешок); 6—дегенерирующие халазальные макроспоры; 7—двуядерный зародышевый мешок; 8—четырехъядерный зародышевый мешок; 9—зрелый четырехъядерный зародышевый мешок; 10—яйцеклетка; 11— синергиды; 12—полярное ядро
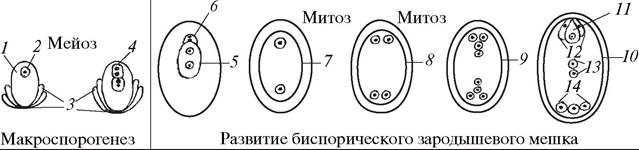
Биспорические зародышевые мешки образуются из диады макроспор и могут быть одномитозными, однополюсными, четырехъядерными (Podostemum- mun) или двухмитозными, двухполюсными, восьмиядерными (Allium-mun) (рис. 184).
Рис. 184. Образование макроспор и особенности развития зародышевого мешка Аllium-типа: 1—нуцеллус семяпочки (макроспорангий); 2—макроспороцит; 3— интегументы семяпочки; 4—микропилярная и халазальная диады макроспор; 5—разрастающаяся халазальная диада макроспор (молодой двуядерный зародышевый мешок); 6—дегенерирующая микропилярная диада макроспор; 7—двуядерный зародышевый мешок; 8—четырехъядерный зародышевый мешок; 9—восьмиядерный зародышевый мешок; 10—зрелый зародышевый мешок; 11—яйцеклетка; 12—синергиды; 13—полярные ядра; 14—антиподы
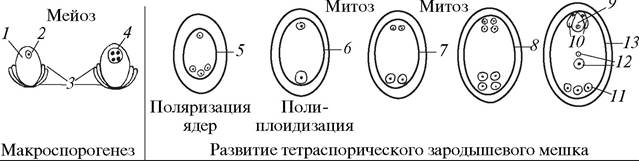
Среди тетраспорических зародышевых мешков описано не менее 10 типов. Они чрезвычайно разнообразны по строению и встречаются у представителей различных семейств.
Своеобразным тетраспорическим двухмитозным, двухполюсным, восьмиядерным зародышевым мешком является Fritillaria-mun (рис. 185). В зародышевом мешке Fritillaria-munа на первом этапе развития происходит неравная поляризация: три ядра четырехъядерного ценоцита отходят к халазальному полюсу, а одно — к микропилярному. В дальнейшем три халазальных ядра сливаются, наблюдается явление полиплоидизации, и образуется одно триплоидное ядро. Таким образом, в результате двух последовательных митотических делений в зародышевом мешке формируется восемь ядер: четыре гаплоидных и четыре триплоидных. На микропилярном полюсе три гаплоидных ядра «одеваются» оболочками, в результате чего образуется яйцеклетка и две синергиды, а на халазальном полюсе образуется три антиподы с триплоидными ядрами. Два полярных ядра различаются по степени плоидности — одно ядро гаплоидное, второе — триплоидное.
Рис. 185. Образование макроспор и особенности развития зародышевого мешка Fritillaria-munа: 1—нуцеллус семяпочки (макроспорангий); 2—макроспороцит; 3— интегументы семяпочки; 4—четырехъядерный ценоцит; 5—неравномерная поляризация ядер в развивающемся зародышевом мешке; 6—двуядерный зародышевый мешок с гаплоидным ядром на микропилярном полюсе и триплоидным ядром на халазальном полюсе; 7—четырехъядерный зародышевый мешок; 8—восьмиядерный зародышевый мешок; 9—яйцеклетка; 10—синергиды; 11—антиподы; 12—полярные ядра; 13—зрелый зародышевый мешок
В результате расхождения ядер четырехъядерного ценоцита к микропилярному, халазальному и двум латеральным (боковым) полюсам образуются четырехполюсные тетраспорические зародышевые мешки. Они могут быть одномитозными 8-ядерными (Рlumbagо-mun) (рис. 186) или двухмитозными 16-ядерными (Реnаеа-mun) (рис. 187).
Рис. 186. Образование макроспор и особенности развития зародышевого мешка Plumbago-munа: 1—нуцеллус семяпочки (макроспорангий); 2—макроспороцит; 3— интегументы семяпочки; 4—четырехъядерный ценоцит (четырехъядерная макроспора); 5—развивающийся четырехполюсный зародышевый мешок; 6—восьмиядерный четырехполюсный зародышевый мешок; 7—зрелый зародышевый мешок; 8—яйцеклетка; 9—полярные ядра
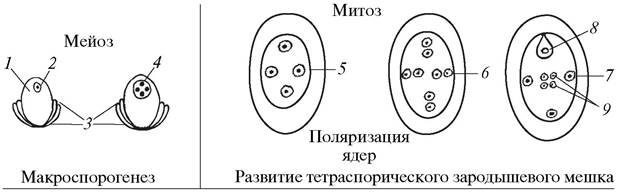
Рис. 187. Образование макроспор и особенности развития зародышевого мешка Penaea-muna: 1—нуцеллус семяпочки (макроспорангий); 2—макроспороцит; 3— интегументы семяпочки; 4—четырехъядерный ценоцит (четырехъядерная макроспора); 5—развивающийся четырехполюсный зародышевый мешок; 6—восьмиядерный четырехполюсный зародышевый мешок; 7—шестнадцатиядерный четырехполюсный зародышевый мешок; 8—зрелый шестнадцатиядерный зародышевый мешок; 9— яйцеклетка; 10—синергиды; 11—антиподы; 12—полярные ядра; 13—клетки на латеральных полюсах 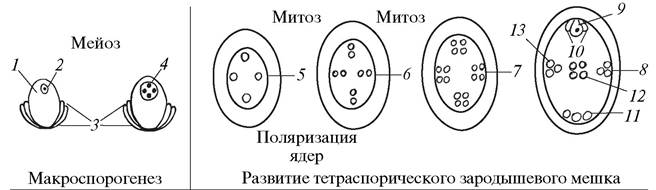
Большинство исследователей считают наиболее примитивным и исходным типом развития зародышевого мешка Polygonum-mun. Сокращение числа митотических делений при формировании зародышевого мешка — одно из главных направлений эволюции женского гаметофита покрытосеменных растений. Это способствует более быстрому развитию женского гаметофита и вполне вероятно имеет адаптивное значение. Однако, по мнению эмбриологов растений, «довольно многочисленные аномалии от нормального типа развития и строения зародышевых мешков у цветковых растений требуют, по-видимому, более широких и детальных исследований с целью выяснения их конкретно-адаптивного онтогенетического или экологического смысла».