ФІЗІОЛОГІЯ ТА БІОХІМІЯ РОСЛИН
Опорний конспект лекцій
6. ДИХАННЯ РОСЛИН
Регуляція процесів дихання
Ендогенні механізми регуляції дихання у рослин
Регуляція процесів дихання здійснюється на різних рівнях.
Це субстратний контроль дихання (доступність, кількість і склад дихальних субстратів); регуляція активності ферментів (оксидоредуктаз); взаємозв’язок різних ланок дихання (дихальних циклів і ЕТЛ мітохондрій, оксидоредуктаз мітохондрій, цитоплазми й інших органоїдів); зміна фізико-хімічних умов у клітині, дія гормонів, геномний контроль і т.д.
Ефект Пастера
Рівень О2 в тканинах впливає на інтенсивність дихання і на величину витрати дихальних субстратів на синтетичні процеси.
Гальмування розкладу цукрів і більш ефективне їх використання при наявності кисню називають ефектом Пастера.
Механізм цього явища такий:
- велика кількість О2 → посилення дихання → збільшення кількості АТФ → зменшення активності фосфофруктокінази і сповільнення гліколізу;
- надлишок АТФ → посилення ресинтезу глюкози;
- збільшення кількості О2→ активація циклу Кребса, ПФШ → утворення проміжних продуктів, необхідних для синтезу.
У присутності О2 всі цикли (і синтетичні) активно працюють. А збільшення кількості АТФ у результаті дихання ще більше сприяє синтезу.
Дихальний контроль
У рослинному організмі існує механізм дихального контролю (акцепторний механізм дихання). Дихальним контролем називають залежність швидкості споживання кисню мітохондріями від концентрації АДФ.
Залежність інтенсивності дихального процесу (ІД) від відношення діючих мас АТФ-системи можна записати так:
![]()
причому, концентрація АДФ є обмежувальним фактором.
При зростанні функціональної активності клітин АТФ витрачається, а АДФ збільшується, а це, у свою чергу, призводить до зростання швидкості дихання, перенесення е- і збільшення інтенсивності окисного фосфорилювання.
Регуляція гліколізу
Інтенсивність гліколізу контролюється на декількох ділянках.
Захоплення глюкози в процес регулюється на рівні ферменту гексокінази за типом зворотного зв'язку: надлишок продукту реакції (глюкозо-6-фосфату) аллостерично пригнічує діяльність ферменту.
Друга ділянка регуляції швидкості гліколізу знаходиться на рівні фосфофруктокінази. Фермент алостерично інгібується високою концентрацією АТФ, а також цитратом і активується неорганічним фосфатом, АДФ та фруктозо-1,6-дифосфатом.
АТФ та ацетил-КоА зменшують активність піруваткінази та піруватдегідрогеназного комплексу; останній пригнічується ще й надлишком НАДН.
Регуляція циклу Кребса (ЦТК)
Використання клітинного ацетил-КоА залежить від енергетичного стану клітини. При незначній енергетичній потребі цикл Кребса гальмується й ацетил-КоА більше використовується на синтетичні процеси. Особливістю регуляції ЦТК є залежність всіх чотирьох дегідрогеназ від співвідношення [НАДН]/ [НАД+].
Активність цитратсинтази гальмується високою концентрацією АТФ і цитратом.
Ізоцитрат-дегідрогеназа інгібується НАДН і активується цитратом.
α-кетоглутаратдегідрогеназа пригнічується продуктом реакції - сукциніл-КоА й активується аденілатами (АДФ, АТФ, АМФ).
Окиснення сукцинату сукцинатдегідрогеназою гальмується оксалоацетатом і прискорюється АТФ, АДФ і відновленим убіхіноном (QH2). Малатдегідрогеназа інгібується оксалоацетатом, і, можливо, АТФ.
Регулюючу роль може відігравати альтернативний шлях транспортування е- в мітохондріях.
Регуляція ПФШ і гліоксилатного циклу
ПФШ окиснення регулюється концією НАДФ+, оскільки містить дві НАДФ- залежні дегідрогенази (глюкозо-6-фосфат- та 6-фосфоглюконат-дегідрогенази). Він регулюється також рівнем синтетичних процесів у клітині, які використовують НАДФН.
Нестача неорганічного фосфату пригнічує гліколіз та активує ПФШ. 6-фосфоглюконова кислота, еритрозо-4-фосфат інгібують гліколіз і сприяють ПФШ.
Активність гліоксилатного циклу знижується при зростанні концентрації оксалоацетату і фосфоенолпірувату.
Екологічні та онтогенетичні аспекти дихання
Вплив факторів зовнішнього середовища
Концентрація кисню. Процес дихання пов'язаний із споживанням кисню. Однак зниження парціального тиску кисню з 21% до 9% практично не впливає на інтенсивність дихання. При 5% вмісті кисню його поглинання у молодих рослин дещо знижується, а виділення СО2 змінюється мало.
Очевидно, у процесі еволюції дихальні системи рослин формувалися в низькокисневих умовах, про що свідчить висока спорідненість цитохромоксидази до кисню. Крім того, це дає можливість рослинам володіти широкою адаптацією до низьких концентрацій О2 (елемент надійності). Така особливість рослин є важливою, бо газовий склад всередині тканин сильно відрізняється від атмосферного. Вміст О2 в паренхімі протягом доби може коливатися від 7 до 17%. При пониженні концентрації О2 до 1-2% дихальний коефіцієнт різко зростає і дихання переходить в анаеробний процес бродіння.
Надлишок О2 виникає лише локально. Він пов'язаний з посиленням у клітині вільнорадикальних реакцій, що може викликати суттєві порушення обміну речовин.
Диоксид вуглецю. Підвищення концентрації СО2, як кінцевого продукту дихання, спричинює зниження інтенсивності процесу. Гальмуються реакції декарбоксилювання й активність сукцинатдегідрогенази, зменшується ДК. При цьому спостерігається закислення тканин - ацидоз, що призводить до шкідливих наслідків. Збільшення концентрації СО2 викликає закривання продихів, гальмування проростання насіння.
Температура. Дихання, як ферментативний процес, залежить від температури. В інтервалі температур від 0 °С до 20 °С Qio дихання ≈ 2-3. При вищих температурах Q10 понижується. Одна з причин - зниження розчинності О2 в рідинах при зростанні температури.
Дихання у зимуючих рослин спостерігається при дуже низьких температурах (-20 °С ÷ -25 °С). Температурний оптимум для видів помірних широт знаходиться у межах 35-40 °С. Максимальні температури (45 ÷ 55 °С) визначаються здатністю білків до денатурації. Важлива і тривалість температурного стресу.
Водний режим. Зміна оводненості тканин рослин віддзеркалюється в динаміці інтенсивності дихання. Швидка тимчасова втрата води посилює дихання - реакція на подразнення. Тривалий водний дефіцит викликає поступове зниження дихання, причому це відбувається повільніше, ніж падіння інтенсивності фотосинтезу. Дихання повітряно-сухого насіння (10-11% Н2О) дуже незначне. При зростанні вологості насіння до 15% дихання зростає у 4-5 разів, а при дальшому зростанні оводненості до 35% дихання збільшується в 1000 разів і більше.
Різке зростання дихання при набряканні насіння супроводжується виділенням тепла, що може призвести до перегріву (»самозгорання») при зберіганні.
Зміна інтенсивності дихання при втраті води залежить також від віку та фізіологічного стану рослини.
Мінеральні речовини. Розчин солей звичайно посилює дихання коренів проростків - ефект «сольового дихання». Вважається, що дія солей на дихання побічна. Деякі катіони металів (магній, калій) підвищують активність окисних ферментів, а їх нестача зменшує дихання. Катіони важких металів пригнічують дихання.
Світло. Дію світла на дихання зелених органів рослин вивчено недостатньо. Освітленість, при якій інтенсивність фотосинтезу = інтенсивності дихання, називають компенсаційним пунктом. Дихання не припиняється на світлі, в нормі його інтенсивність і розпад органічних речовин повинні бути значно меншими, ніж інтенсивність фотосинтезу.
Дихання (поглинання кисню і виділення вуглекислого газу) незелених тканин активізується світлом короткохвильової частини спектру - ультрафіолетом (380 нм), синіми й зеленими променями (400-500 нм)
Пошкодження. Механічні впливи викликають короткочасне посилення поглинання кисню. При цьому натискання і згинання впливають слабо, а зрізання й порізи - дуже сильно. В останньому випадку різні субстрати з'єднуються з оксидазами, а також активізуються процеси відновлення.
Зміна інтенсивності дихання в онтогенезі
Молоді органи й тканини рослин, які знаходяться в стані активного росту, мають найвищу інтенсивність дихання. Поки молодий листок росте й розвивається, інтенсивність дихання зростає. Потім дихання знижується до рівня половини максимального і лише перед пожовтінням спостерігається короткочасний підйом. Цвітіння й плодоношення супроводжується посиленням дихання квітів та плодів. Перед повним дозріванням соковитих плодів спостерігається тимчасове посилення дихання - клімактеричний підйом дихання. Перед цим у тканинах зростає утворення етилену. Клімактеричним явищам запобігають, зберігаючи плоди у холодних приміщеннях, насичених СО2 і азотом.
Клітинне дихання - це сукупність біохімічних реакцій, у ході яких відбувається окиснення вуглеводів, ліпідів і амінокислот до вуглекислого газу і води.
Вивільнена енергія запасається в хімічних зв’язках молекул АТФ.
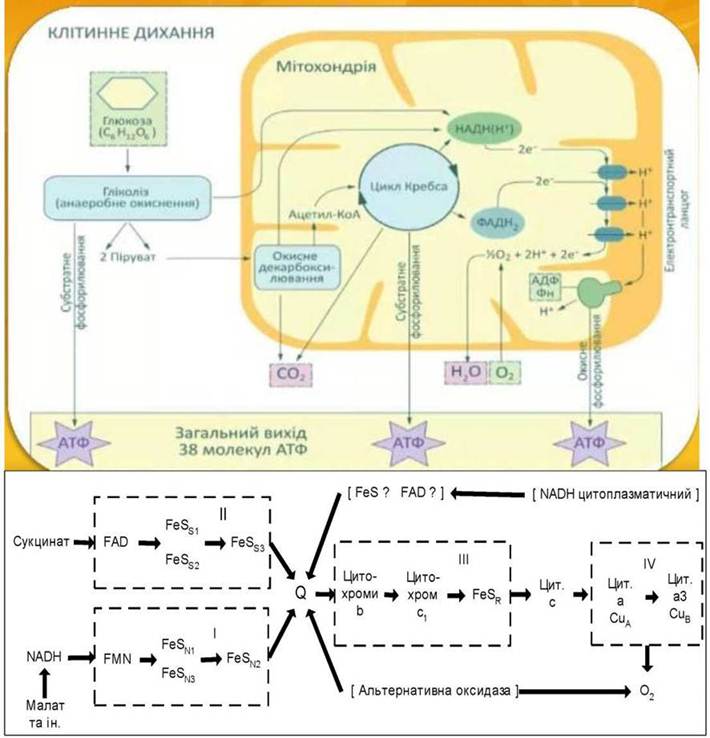
Рис. 16. Електронно-транспортний ланцюг дихання рослинних мітохондрій.
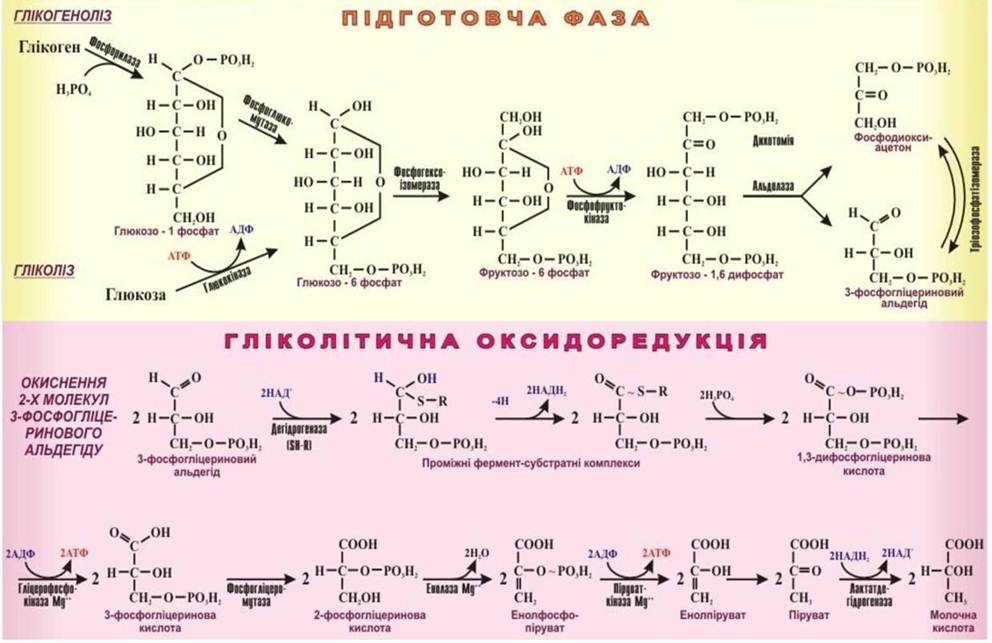
Рис. 17. Анаеробне розщеплення вуглеводів.
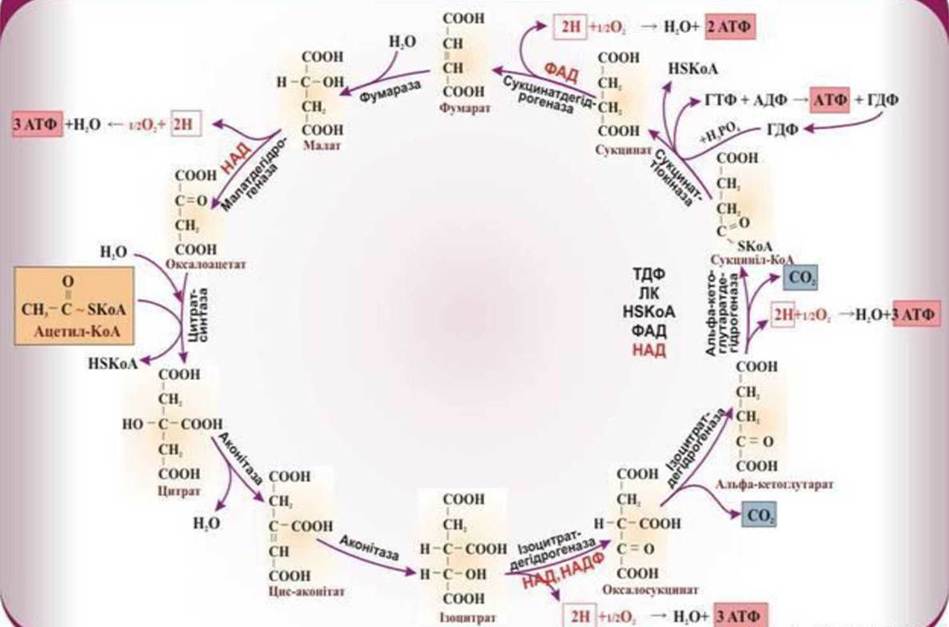
Рис. 18. Цикл трикарбонових кислот Кребса.