ФІЗІОЛОГІЯ ТА БІОХІМІЯ РОСЛИН
Опорний конспект лекцій
7. МІНЕРАЛЬНЕ ЖИВЛЕННЯ РОСЛИН
Азотне живлення
Надходження азоту в рослини і його перетворення
Молекулярний азот (N2) складає близько 80% атмосферного повітря. Однак у такому вигляді він недоступний для вищих рослин. Вони можуть засвоювати лише його окисли, або аміак, яких дуже мало в атмосфері. Тому основним джерелом азоту для рослин служать його мінеральні сполуки з ґрунту. Рослини мають здатність до автотрофного засвоєння не лише вуглецю, але й азоту.
Автотрофна асиміляція азоту
Азот знаходиться в ґрунті у вигляді різних сполук. Значна їх частина входить до складу гумусу (продукт неповного розкладу органічних залишків), який засвоюється рослиною лише після мінералізації ґрунтовою мікрофлорою на більш доступні сполуки, перш за все - на аміак. У газоподібній формі аміак легко засвоюється рослинами, але в такому вигляді його у ґрунті мало. Звичайно аміак розчиняється в ґрунтовій воді і, вступаючи у взаємодію з різними кислотами, дає амонійні солі або окиснюється бактеріями-нітрифікаторами і перетворюється в азотисту і азотну кислоти, які утворюють із катіонами нітрити й нітрати. Амонійні солі та нітрати - водорозчинні і є головною формою ґрунтового азоту, доступного рослинам.
Рослини для свого розвитку потребують значної кількості азоту.
Амонійний азот поступає до коренів рослини у відновленій формі і тому зразу ж бере участь в утворенні амінокислот та білків.
У нітратах азот знаходиться в окисненому стані. Тому, перш ніж залучитися до обміну речовин, він повинен відновитися до аміаку.
Перетворення нітратів у аміак у клітинах кореня називається їх хімічним відновленням. Це відбувається в результаті двох послідовних реакцій, що відбуваються з участю нітрат- і нітритредуктаз, які містять молібден і залізо. Спочатку нітрати відновлюються нітратредуктазою в нітрити, які згодом відновлюються нітритредуктазою до аміаку:
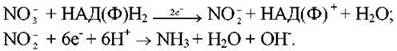
Необхідною умовою протікання даних реакцій є наявність відновника (донора електронів) і енергії. Джерелом е служить НАДН, а енергії - АТФ, які утворюються в процесі дихання кореня. Для активного відновлення нітратів у корені завжди повинен бути резерв вуглеводів, що використовується як субстрат дихання й джерело кетокислот, здатних зв’язувати аміак. У протилежному випадку аміак скупчується й гальмує реакцію відновлення нітратів.
За відсутності хоча б однієї з цих умов або при надходженні до коренів великої кількості нітратів, вони тут не метаболізуються, а піднімаються з транспіраційним потоком по стеблах до листків і там відновлюються до аміаку з участю НАДФН2 і АТФ, які утворюються в результаті фотосинтезу. Тому даний процес називається фотохімічним відновленням нітратів.
Аміак, який синтезується в результаті відновлення нітратів, шляхом прямого амінування приєднується до первинних кетокислот (ПВК, ЩОК, α-КГК) і дає відповідні первинні амінокислоти(аланін, аспарагінову та глютамінову кислоти). Ці реакції відновного амінування каталізуються специфічними дегідрогеназами (аланін-, аспартат-, глутаматдегідрогенази) і протікають з участю НАД(Ф)Н2. Первинні амінокислоти в результаті перебудови їх вуглеводневих скелетів перетворюються на інші амінокислоти і разом із ними використовуються у синтезі первинного білка. Весь цей процес являє собою анаболічну гілку азотного обміну. Більшість амінокислот використовується для синтезу необхідних для клітини білків, а вже непотрібні поліпептиди розпадаються до амінокислот. Вони можуть повторно використовуватися на синтез білків чи брати участь у реакціях переамінування, а можуть дезамінуватися з утворенням кетокислот і аміаку. Цей процес являє собою катаболічну гілку азотного обміну у рослині.
Аміак, який надійшов до рослини чи утворився при біохімічних перетвореннях - токсичний для організму. Але він не виводиться у зовнішнє середовище, як у тварин, а зв’язується аспарагіновою чи глютаміновою кислотами з утворенням відповідних амідів - аспарагіну й глютаміну. Даний процес відбувається з участю синтетаз і вимагає енергії.
Аміди являють собою своєрідне депо азоту, де дефіцитний для рослини аміак резервується і знешкоджується. Аміногрупи первинних амінокислот і їх амідів за допомогою амінотрансфераз зв’язуються з різними кетокислотами. У результаті утворюються вторинні амінокислоти. Ферментативне перенесення аміногруп з амінокислот і амідів на кетокислоти без проміжного утворення аміаку отримав назву переамінування. В реакціях переамінування зв’язуються анаболічна та катаболічна гілки азотного обміну, початковою і кінцевою ланкою якого є аміак.
Крім автотрофної асиміляції N, існує гетеротрофна, коли деякі види рослин пристосувалися задовольняти свої потреби в азоті шляхом засвоєння азотовмісних сполук інших організмів. Це - рослини-паразити, напівпаразити, сапрофіти (поглинають гниючі органічні залишки), комахоїдні рослини.
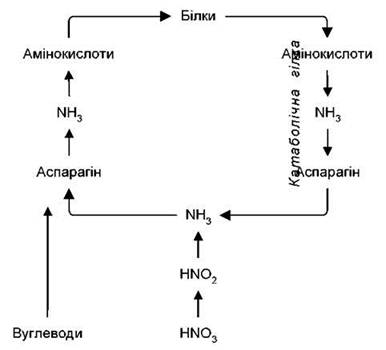
Рис. 19. Перетворення азотистих речовин у рослині.
Фіксація молекулярного азоту
Молекулярний азот можуть зв’язувати тільки деякі бактерії. Вони відновлюють його в доступну для інших організмів аміачну форму. Тому їх називають азотфіксаторами. Розрізняють вільноживучі та симбіотичні азотфіксувальні бактерії.
Вільноживучі азотфіксатори заселяють водойми та ґрунти і становлять велику групу мікроорганізмів різних видів. До них належать бактерії родів Clostridium і Azotobacter, фотосинтезуючі бактерії та мікобактерії, актиноміцети, синьо-зелені водорості (або ціанобактерії) та лишайники.
Усі вони за певних екологічних умов збагачують ґрунт зв’язаними формами азоту. Так, основним постачальником його на рисових полях і у водах Світового океану є синьо-зелені водорості. Первинний ґрунт збагачується азотом у результаті діяльності лишайників. На підзолистих і торф’яних ґрунтах головним азотонакопичувачем служать мікобактерії, оскільки діяльність інших азотфіксаторів тут пригнічується високою кислотністю.
Механізм біологічної фіксації поки що повністю не розкритий. Встановлено, що цей процес здійснюється за допомогою нітрогенази - особливого ферментного комплексу, який містить дві білкові фракції. Одна з них складається з азофередоксину, що містить негемінне залізо (4FeS), друга — із молібденофередоксину, до якої, крім негемінного заліза у вигляді кластерів FeS, входять два атоми Мо.
Джерелом необхідного для даного процесу відновника молекулярного азоту служить відновлений фередоксин (постачає ē), а джерелом енергії - АТФ, яка утворюється при бродінні (в анаеробних азотфіксаторів), або диханні (в аеробних). Активатором даної реакції є іони Mg2+.
У загальних рисах процес зв’язування молекулярного азоту відбувається так. Спочатку азофередоксин з участю АТФ і електронів змінює конфігурацію і відновлюється, а потім передає електрони на молібденофередоксин, який уже безпосередньо взаємодіє з молекулярним азотом і відновлює його до аміаку.
Продуктивність зв’язування молекулярного азоту вільноживучими азотфіксаторами становить декілька десятків кілограмів азоту на гектар.
Азотфіксатори-симбіотики являють собою численну групу так званих бульбочкових бактерій (рід Rhisobium), що розвиваються на коренях бобових, а також багатьох деревних та кущових (чагарникових) порід. До групи симбіотичних азотфіксаторів також належать деякі актиноміцети і ціанобактерії. У вільному стані вони не фіксують азот; цю властивість отримують, лише вступивши у симбіоз із вищими рослинами. В даний час відомо близько 190 видів дерев та кущів різних родин, із якими зазначені мікроорганізми вступають у симбіотичні відносини. Вони проникають до клітин кореня через кореневі волоски за допомогою особливого слизистого тяжу, який полегшує просування до глибших шарів тканин.
Очевидно, ці бактерії виділяють якісь речовини типу гормонів, які стимулюють поділ клітин кореня й утворення потовщень (бульбочок). У клітинах бульбочок бактерії змінюються фізіологічно й морфологічно, отримують додаткову мембрану і перетворюються в так звані бактероїди.
1967 року Бергерсен із допомогою ізотопу азоту 15N встановив, що азотфіксацію здійснюють власне клітини бактероїдів. Важливу роль у процесі зв’язування азоту відіграють клітини кореневих волосків. Вони містять рожевий пігмент легоглобін, який, подібно до гемоглобіну, зв’язує кисень і постачає ним бактероїди, що залягають у глибині бульбочок.
Механізм біологічної фіксації молекулярного азоту бактеріями-симбіотиками принципово не відрізняється від механізму зв’язування азоту вільноживучими азотфіксаторами. Цей процес здійснюється аналогічним ферментним комплексом і вимагає затрати великої кількості енергії.
Бульбочкові бактерії мають багато рас, кожна з яких живе на коренях тільки одного чи декількох видів рослин. Від характеру раси залежить продуктивність азотфіксації. Найбільш активними азотонакопичувачами є симбіонти люцерни (дають приблизно 500-600 кг зв’язаного азоту на 1 га за рік), конюшини (300 кг/га), люпину (160 кг/га). Менш ефективні симбіонти гороху, квасолі, бобів (5060 кг/га). Бульбочкові бактерії деревних порід зв’язують близько 100 кг азоту на 1 га за рік. У цілому продуктивність азотфіксації бульбочкових бактерій набагато вища, ніж у вільноживучих азотфіксаторів. Це пояснюють тим, що в результаті симбіозу з вищими рослинами бульбочкові бактерії краще забезпечуються субстратами дихання та іншими необхідними метаболітами.
Продуктивність азотфіксації залежить від характеру взаємовідносин між партнерами по симбіозу та від активності штаму. Ефективні штами на перших стадіях проникнення й утворення бульбочок виявляють щодо вищої рослини явний паразитизм. Вони використовують її метаболіти, не фіксуючи азот. У рослин з’являються ознаки пригнічення життєдіяльності. Однак із розростанням бульбочок і формуванням бактероїдів виникає та збільшується азотфіксувальна здатність бактерій і взаємовідносини партнерів набувають взаємовигідного характеру. До моменту цвітіння однолітніх бобових постачання бульбочок поживними речовинами знижується, бактероїди та клітини бульбочок лізують і фіксація атмосферного азоту припиняється.
Бульбочки багаторічних трав фіксують азот протягом декількох років, але до кінця кожного періоду вегетації частина їхніх клітин із бактероїдами деградує, тому інтенсивність азотфіксації також знижується.
При інфікуванні рослин неефективним штамом у бульбочках утворюється дуже мало легоглобіну і продуктивність азотфіксації практично дорівнює нулю. В таких випадках бактерії лише пригнічують ріст і розвиток вищої рослини. Тому в сільськогосподарській практиці насіння бобових культур спеціально інфікують бактеріальним препаратом нітрагіном, який містить високоефективний штам бульбочкових бактерій.
Біологічна фіксація молекулярного азоту відіграє в житті екосистеми виключно важливу роль. Велике значення азотфіксаторів у природних біогеоценозах, де різноманітні втрати азоту (вимивання солей, денітрифікація і т.д.) поповнюються головним чином за рахунок їхньої діяльності і лише частково - у результаті розкладу тваринних і рослинних рештків.
В агробіоценозах утрати азоту ще більші, оскільки тут значна його частина забирається разом із врожаєм. Наприклад, при врожаї картоплі 250-300 ц/га із ґрунту виноситься приблизно 200 кг азоту. Тому бобові є важливою складовою частиною будь-якої сівозміни. Однак значну частину втрат азоту доводиться поповнювати шляхом внесення органічних і мінеральних добрив.