БОТАНИКА ТОМ 1 - КЛЕТОЧНАЯ БИОЛОГИЯ. АНАТОМИЯ. МОРФОЛОГИЯ - 2007
5. ПРИНЦИПЫ СТРОЕНИЯ ТАЛЛОМНЫХ РАСТЕНИЙ (ТАЛЛОФИТОВ)
5.4. Мицелий грибов
Вегетативные тела большинства грибов (и, следовательно, всех лишайников) построены из разделенных на клетки нитей, называемых гифами (от греч hyphe — ткань из ниток) Гифы растут исключительно апикально, только на верхушке гифы длиной менее 20 мкм, клеточная оболочка настолько мягкая и пластичная за счет постоянного встраивания в нее нового материала, что способна растягиваться под тургорным давлением. Грибные гифы могут расти очень быстро до 1 мм/ч. Быстрый рост и частые ветвления в субстрате (почве, разлагающейся листве, стволах деревьев и т. п, а также в жидкостях) способствуют возникновению рыхлого сплетения нитей с огромной общей поверхностью — это мицелий (греч myzen — сосать, mykes — гриб, рис 5.11). Гифы не имеют кутикулы, поэтому мицелий легко высыхает, но зато хорошо приспособлен к осмотрофному поглощению растворенных веществ. Многие высшие растения используют это, вступая в симбиоз с грибами и образуя микоризу (см 9 2 3). Впрочем, по тем же причинам грибы легко аккумулируют и ядовитые тяжелые металлы (например, кадмий) и радионуклиды, что следует принимать во внимание при современном уровне загрязнения окружающей среды.
Рис. 5.11. Мицелии грибов (СЭМ-фотографии: А—L. Schreiber; В—V. Kern): А—рыхлый мицелий на поверхности однолетней хвоинки пихты (62х); В—плектенхима зимнего грибаFlammulina velutipes (Basidiomycota) из семейства рядовок
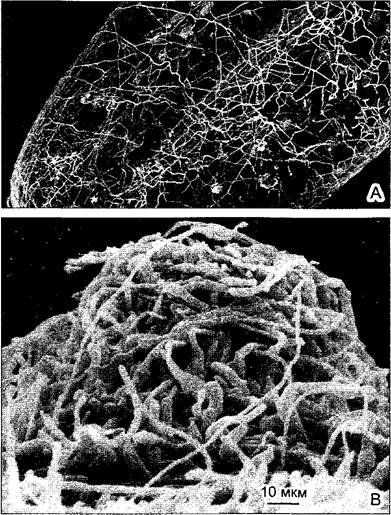
В то время как гифы низших грибов не разделены перегородками на клетки и полиэнергидны, т.е. находятся на сифональной ступени организации, гифы аскомицетов и базидиомицетов поделены на клетки поперечными перегородками (септами), так что имеют клеточное строение (см. рис. 11.43). В септах, правда, имеются центральные поры диаметром от 50 до 500 нм, так что и у высших грибов цитоплазма непрерывна во всей бластеме. При формировании недолговечных плодовых тел — именно их принято называть грибами — мицелий уплотняется в типичную плектенхиму (см. рис. 5.11, В). При этом в типичных случаях ткани, такие как проводящие, механические и т.п., не дифференцируются, но образующий споры гимений нередко сильно отличается от остальной плектенхимы. Однако в долго живущих плодовых телах трутовиков есть механические толстостенные, длинные и обычно разветвленные скелетные гифы, оплетенные также жесткими, но короткими связующими гифами (см. рис. 11.58).
5.5. Формы организации печеночников и лиственных мхов
Большинство мхов — наземные обитатели влажных биотопов. Особенно пышная моховая растительность встречается в дождевых и туманных горных лесах, в тенистых местах и у водотоков, а также на низинных и верховых болотах. Наличие мхов в определенном смысле указывает на повышенную влажность. Они нуждаются в капельножидкой воде для оплодотворения, так как мужские гаметы (сперматозоиды, образующиеся в антеридиях) имеют жгутики и вплавь добираются до расположенных в архегониях яйцеклеток. Кроме того, одноклеточные или многоклеточные ризоиды лить на несколько миллиметров внедряются в почву, так что не могут достичь глубоких водоносных ее слоев. (Корней у мхов нет.). Наконец, и способность проведения воды в надземные органы весьма ограниченна.
Рис. 5.12. Вильчатое ветвление талломов бурых водорослей (А—D) и печеночников (Е, F) (А—по Н.Schenk; В —D—по de Wildeman; Е—по W. Kingmüeller; F—по L. Кпу).
В то время как у бурой водоросли Dictyota dichotoma (А, 0,5х) имеется настоящая дихотомия — деление верхушечной клетки (В—D, 250х), у печеночников (например, у Riccia rhenana, Е, 2,5х) ветвление таллома начинается благодаря новообразованию дополнительной двухсторонней верхушечной клетки S2рядом с уже имевшейся S1 (F, 370х)
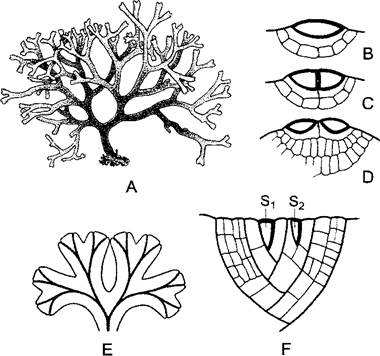
Рис. 5.13. Верхушка побега лиственного мха Fontmalis antipyretica (А—по H. Leitgeb; В—по О. Stocker): А—продольный срез (120х); В—вид сверху; трехсторонняя верхушечная клетка закрашена. Каждый образованный ею сегмент делится периклинальной стенкой на внутреннюю и наружную (коровую) клетку. Последняя образует ткань коры и лист. Боковые веточки возникают ниже листа путем образования новой трехсторонней верхушечной клетки z. У Fontinalis, погруженного обитателя текучих вод, листья расположены тремя продольными рядами. У большинства остальных лиственных мхов листья слегка асимметричны, что приводит к очередному их расположению
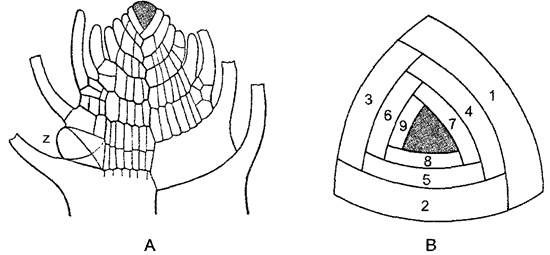
У печеночников талломы находятся на высоком уровне дифференцированности. У многих из них многослойные тканевые талломы плоские, они растут с помощью двухсторонних верхушечных клеток (рис. 5.12, Е, F). Вильчатое ветвление их происходит не путем равного деления самой верхушечной клетки (как, например, у бурых водорослей, — настоящая дихотомия), а путем превращения клетки, отчлененной от исходной верхушечной клетки, в новую, вторую верхушечную клетку. У так называемых слоевищных печеночников, например, у маршанции (Marchantía, см. рис. 11.111), талломы лежат на почве. Антеридио- и архегониофоры возвышаются на вертикальных ножках более 1 см длиной над талломом. У многих листостебельных печеночников вместо плоского лопастного таллома образуются стелющиеся по поверхности почвы побеги с листьями толщиной в один слой клеток без средней жилки. (Стебель и листья печеночников не тождественны куда более сложным стеблям и листьям кормофитов.). Еще более продвинута организация лиственных мхов. Их побеги растут благодаря трехсторонней верхушечной клетке1 (рис. 5.13), их стебли имеют сложную тканевую структуру с центральным проводящим пучком (центральный тяж, рис. 5.14, 1), ответвления которого часто продолжаются в листья как средняя жилка. Строение листьев может быть усложнено вследствие закономерной смены равных и неравных делений клеток (сфагнум, рис. 5.15; 11.120); возможно также увеличение поверхности листьев при образовании на их верхней стороне продольных гребней из клеток, особенно богатых хлоропластами (см. рис. 11.122, А).
1 Такая же клетка находится на верхушке побега у листостебельных печеночников. — Примеч. ред.
Рис. 5.14. Поперечный срез стебля лиственного мха Mnium undulatum(90х) (по Е. Strasburger): а—покровная ткань; I—центральный проводящий пучок; р—коровая паренхима; rh—ризоиды

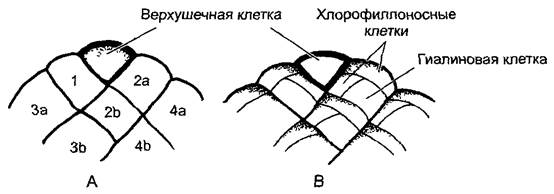
Рис. 5.15. Деления клеток в листке торфяного мха Sphagnum (В—по Е. Вunning): А—двухсторонняя верхушечная клетка отделяет налево и направо сегменты (1—4), которые затем снова делятся на одинаковые ромбические клетки (2а, 2b, За, Зb...); В—после прекращения делений верхушечной клетки каждая ромбическая клетка делится двумя неравными делениями на две хлорофиллоносные и одну гиалиновую клетку (150х). Гиалиновые клетки отмирают после того, как их стенки укрепляются спиральными утолщениями и обретают большое отверстие наружу; гиалиновые клетки служат для впитывания воды (см. рис 11.120, G, Н)
Лиственные мхи, близкие по своей организации кормофитам, в ходе развития уникальным образом проходят разные типы организации таллофитов. С прорастания споры начинается рост на трихальной ступени организации и образуется протонема — сплетение из однорядных разветвленных клеточных нитей (гаплонем). На этом нитчатом талломе вырастают образующие половые клетки гаплоидные гаметофиты в виде побегов. На них возвышаются развивающиеся из зигот диплоидные спорофиты, на которых (не на листьях!) находятся простые устьица.