БОТАНИКА ТОМ 1 - КЛЕТОЧНАЯ БИОЛОГИЯ. АНАТОМИЯ. МОРФОЛОГИЯ - 2007
1. МОЛЕКУЛЯРНЫЕ ОСНОВЫ— СТРОИТЕЛЬНЫЕ «КИРПИЧИКИ» КЛЕТОК
1.2. Нуклеиновые кислоты
Нуклеиновые кислоты — это гетеро- полимерные молекулы, которые служат либо для хранения информации (дезоксирибонуклеиновая кислота, ДНК/DNA), либо для переноса и реализации информации (рибонуклеиновые кислоты, РНК/ RNA). Кроме того, определенные РНК обладают структурной функцией при построении рибосом (рибосомальная рРНК/ rRNA). У всех клеточных организмов — как про-, так и эукариот — двухцепочечная ДНК служит для хранения наследственной информации и ее передачи в ряду поколений путем матричного удвоения. Молекулы ДНК отличаются тем, что могут служить основой для образования новых молекул ДНК с точно такой же последовательностью оснований (репликация; см. 1.2.3). У всех организмов, кроме РНК- содержащих вирусов и вироидов (см. 1.2.5), только ДНК обладает такой функцией репликативного удвоения. Так как размножение, продолжение рода, наследование являются основными критериями жизни, репликативное удвоение ДНК стоит в центре всех жизненных процессов. Кроме того, молекулы ДНК способны служить матрицей для последовательностей РНК, а через РНК — также передавать информацию о последовательности аминокислот в белках. Благодаря этой генетической функции ДНК может проявляться наследственная информация: наследственные факторы (гены; от греч. genos — происхождение, род, наследство) реализуются как определенный фенотип (набор проявляемых внешне признаков организмов; от греч. phainein — делать видимым).
1.2.1. Строение нуклеиновых кислот
Нуклеиновые кислоты это полинуклеотиды, неразветвленные полимеры из мономеров — нуклеотидов. Один нуклеотид состоит из основания, связанного N-гли- козидной связью с сахаром, — нуклеозида — и от одного до трех остатков фосфорной кислоты (фосфата), так что различают нуклеозидмоно-, ди- и трифосфаты (рис. 1.3).
Рис. 1.3. Нуклеотиды состоят из трех структурных элементов — пиримидинового или пуринового азотистого основания сахара(пентозы)и фосфорной кислоты.
Азотистое основание связано с сахаром (N-β- гликозидной связью пиримидина или N9 пурина. Фосфорная кислота образует эфир с первичной спиртовой группой пентозы К α-остатку фосфорной кислоты может быть присоединено еще до двух ее остатков с помощью ангидридных связей. Гликозид из азотистого основания и рибозы называют нуклеозидом, а из основания и 2-дезоксирибозы — дезоксинуклеозидом (d-nucleosid) Таким образом, нуклеотиды являются нуклеозидмоно-, ди- или трифосфатами, как это показано внизу на примере аденозина и его нуклеотидов. В качестве сахара в рибонуклеиновой кислоте встречается рибоза, в дезоксирибонуклеиновой кислоте (ДНК = DNA)-2'-дезоксирибоза. Атомы углерода сахара обозначают 1', 2'...5"; С1 образует гликозидную связь. Для упрощения сложных структурных формул, например, в кольцах, обычно опускают стоящие при атомах углерода водородные атомы (сравните нижнюю формулу с расположенными выше). Этот прием часто используется и в других рисунках книги для наглядности
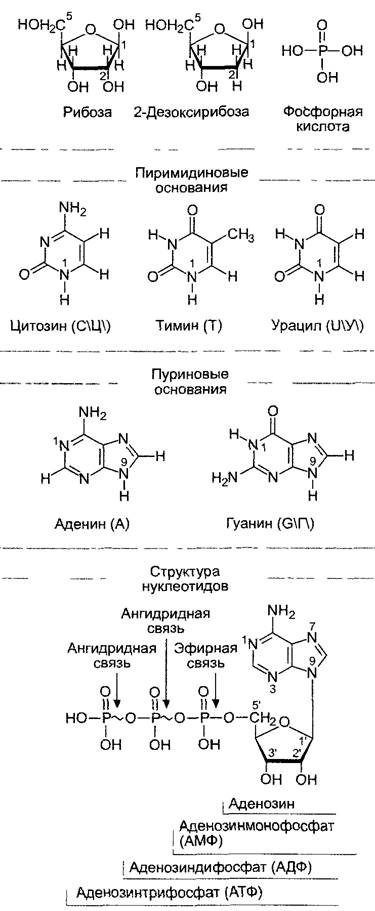
В качестве оснований в ДНК (DNA) встречаются пурины — аденин (А) и гуанин (Г, G), а также пиримидины — цитозин (Ц, С) и тимин (Т). В РНК (RNA) тимин заменен на урацил (У, U). Понятие «основание» (или азотистое основание) указывает на щелочную природу этих гетероциклических, азотсодержащих ароматических соединений. Связь с сахаром осуществляется через N1пиримидина или N9 пурина. В качестве сахара служат пентозы — рибоза в РНК и 2-дезоксирибоза в ДНК, в обоих случаях в ß-D-фуранозной форме (о номенклатуре сахаров см. 1.4.1). Нуклеозиды называют: аденозин, гуанозин, уридин и иитидин, если речь идет о РНК, и дезоксиаденозин, дезоксигуанозин, дезокситимидин и дезоксицитидин, когда в нуклеиновой кислоте (ДНК) содержится 2-дезоксирибоза. Благодаря образованию эфирной связи первичной гидроксильной группы у Cs-атома пентозы с фосфорной кислотой возникает нуклеозидмонофосфат. Одна или две других молекулы фосфорной кислоты могут подсоединиться к этой α-фосфатной группе так, что образуетсябогатый энергией ангидрид — нуклеозиддифосфат или нуклеозидтрифосфат. Нуклеозидтрифосфаты используются на первом этапе биосинтеза ДНК или РНК. Кроме того, эти соединения с высокой энергией переноса групп обладают и многими другими функциями в обмене веществ. Например, аденозинтрифосфат (АТФ, АТР) является важнейшим поставщиком энергии для многих ферментативных реакций (см. 6.1).
Через α -остаток фосфорной кислоты нуклеотид может быть ковалентно связан с пентозой второго нуклеотида с выделением воды, так что сначала получается динуклеотид. Из него может быть образован олиго- и, наконец, полинуклеотид. При этом в нуклеиновых кислотах образуются фосфодиэфирные мостики между 5' и 3' С-атомами соседних пентоз (чтобы различать атомы азотистых оснований и сахаров, С-атомы сахара нуклеозидов обозначают цифрами со штрихом). Как показано на рис. 1.4, тип нуклеиновой кислоты зависит от сахарофосфатного остова молекулы, который состоит из мономеров рибозы (или дезоксирибозы), связанных 5',3'-фосфодиэфириыми мостиками. На одном конце молекулы расположена свободная З'-ОН-группа (5'-конец нуклеиновой кислоты), на другом конце — свободная З'-ОН-группа (З'-конец). Азотистые основания связаны с этим сахарофосфатным остовом гликозидной связью.
Рис. 1.4. Короткие последовательности молекул ДНК и РНК.
Тимин не встречается в РНК и заменен в ней урацилом. Направление синтеза и прочтения формул слева направо, от 5'- к З'-концу молекулы
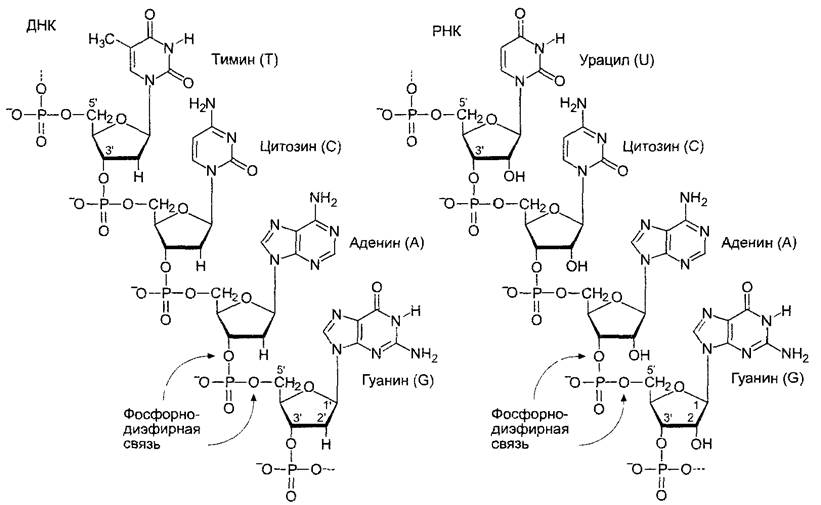
Эта первичная структура нуклеиновых кислот определяется характерной линейной последовательностью азотистых оснований, которая читается всегда в направлении 5' —» 3', что соответствует и направлению синтеза. Последовательность оснований (триплетный код; см. 7.3.1 1) несет информацию о структуре белка. Мерой величины молекулы нуклеиновой кислоты принято считать число пар оснований для ДНК (bр — от англ, base pairs, или п. н. — пар нуклеотидов ДНК) или число оснований для РНК (b — base, или н. — нуклеотидов РНК).
1.2.2. Структура дезоксирибонуклеиновой кислоты (ДНК, DNA)
Лишь у немногих фагов и вирусов ДНК встречается в форме однонитевой молекулы (ssDNA, англ, single-stranded DNA). У многих вирусов и фагов, а также в любых клетках ДНК представлена в виде двойной нити из двух спирально закрученных антипараллельных молекул (dsDNA, англ, double-stranded DNA). Эту структуру принято называть двойной спиралью ДНК. Сахарофосфатные цепочки при этом направлены наружу, а плоские гетероциклические кольца оснований располагаются почти перпендикулярно продольной оси двойной спирали внутри нее (рис. 1.5). Противолежащие основания обеих цепей находятся на одном уровне и образуют в области оси спирали водородные связи (см. 1.1). Это, разумеется, предполагает пространственное (стерическое) соответствие прилегающих друг к другу участков гетероциклов (рис. 1.6). Одно пуриновое основание (А или Г) всегда стоит напротив пиримидинового основания (Т или Ц), и комплементарны только пары оснований АТ и ГЦ.
Рис. 1.5. Модель двойной спирали ДНК Уотсона— Крика (В-форма): А—схема; В—полусферическая модель
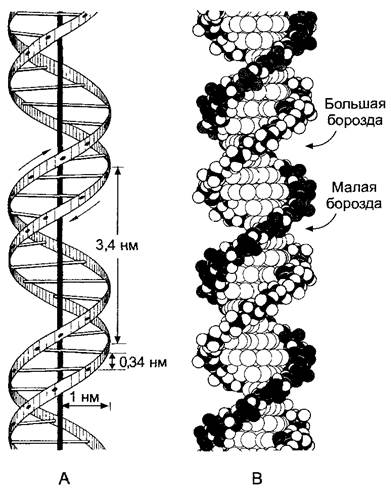
Рис. 1.6. Специфическое спаривание азотистых оснований путем образования водородных связей между двумя цепями ДНК.
Молекулярная структура оснований допускает соединение только АТ и ГЦ. В паре АТ образуются две, а в паре ГЦ — три водородных связи
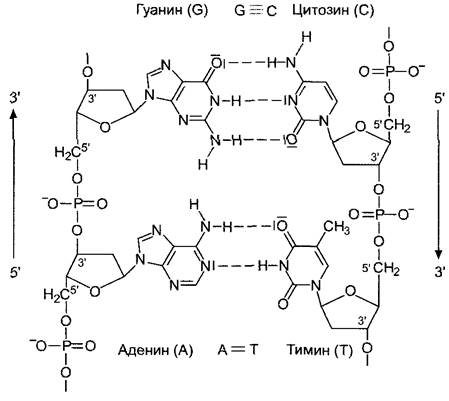
Соответственно комплементарны и последовательности оснований в обеих нитях двойной спирали ДНК, благодаря чему они прочно удерживаются одна против другой. Так, для последовательности оснований 5'- ГАТТАЦА-3' в противостоящей нити должна соответствовать З'-ЦТААТГТ-З'. Из принципа строения молекулы ДНК следует, что молярное соотношение пуриновых и пиримидиновых оснований в двойной спирали всегда равно 1:1, т.е. имеется столько же Ц, сколько Г, и столько А, сколько Т (т. е. А + Г - Т + Ц). Напротив, соотношение оснований (А + Т): (Г + Ц) может варьировать. И хотя для ДНК одного вида организмов это соотношение постоянно и даже является специфическим признаком вида, уже у родственных видов и рас оно отличается. У прокариот соотношение оснований (А + Т): (Г + Ц) колеблется в широких пределах (0,3 — 3,5), для эукариот оно близко к 1. От соотношения оснований (А + Т):(Г + Ц) зависит и температура плавления (Тm) ДНК. Под плавлением, или денатурацией, ДНК понимают, к примеру, вызванное нагревом расхождение обеих нитей ДНК. При этом разрываются водородные связи между противолежащими основаниями. ГЦ- пара с ее тремя водородными связями стабильнее, чем АТ-пара только с двумя водородными связями, поэтому последовательности ДНК, богатые парами АТ, разделяются при более низких температурах, нежели богатые ГЦ-парами.
Последовательность оснований в цепях молекулы ДНК называют ее первичной структурой, а спиральную структуру — вторичной структурой. Связывающие ДНК белки часто узнают вторичную структуру в определенных местах двойной спирали ДНК (см. 7.2.2.3). Представленная на рис. 1.5 модель двойной спирали, предложенная в 1953 г. Д. Уотсоном и Ф. Криком на основе данных рентгеноструктурного анализа М.Уилкинса и Р.Франклин, демонстрирует преобладающую В-форму ДНК —
правозакрученную спираль. Как тогда полагали, двойная спираль ДНК имеет диаметр около 2 нм (20 А). В одном полном обороте спирали (3,4 нм в направлении оси) помещается 10 спаренных оснований, т.е. каждое основание поворачивается на 36° относительно соседнего. Более точные измерения, произведенные позже, показали, что на самом деле на один виток В-ДНК, приходится 10,5 пар оснований на протяжении 3,6 нм. Наряду с В-формой, существует также правозакрученная A-форма ДНК, которая отличается конформацией сахара. При специальных условиях может существовать левозакрученная 2-форма ДНК. Она представляет собой цепь чередующихся пуриновых и пиримидиновых оснований. Z- ДНК, как полагают, могла бы принимать участие в генной регуляции. Двойная спираль ДНК подвижна, т.е. может слегка изгибаться с минимальным радиусом изгиба около 5 нм (например, в нуклеосомах) (см. рис. 2.21). В клетках двойная спираль ДНК находится не в беспорядочном состоянии, а образует третичную структуру, которая может при участии многочисленных белков создавать очень компактную суперструктуру хромосом эукариотических клеток (см. 2.2.3.2).
1.2.3. Репликация ДНК
Обе нити двойной спирали ДНК из-за комплементарности оснований находятся в позитивном/негативном отношении друг с другом. Таким образом, самой структурой предопределена возможность идентичного удвоения, репликации наследственного материала. Нити расходятся, и на каждой заново образуется комплементарная ей вторая нить спирали (рис. 1.7). Эта модель полуконсервативной репликации была многократно подтверждена экспериментально. Было показано, что все хромосомы эукариот реплицируются полуконсервативно. Далее установили, что нереплицированные хромосомы содержат только одну двойную спираль ДНК (одноцепочечная модель; см. 2.2.3.2), а реплицированные (после S-фазы) — две двойные спирали.
Рис. 1.7. Доказательство полуконсервативной репликации ДНК в эксперименте Мезельсона—Сталя
В присутствии тяжелого изотопа азота 15N в среде, на которой выращивали клетки бактерии Escherichia coli, образуется содержащая 15N ДНК с удельной плотностью 1,724 г см 3(определяется путем центрифугирования в градиенте плотности CsCI). Если затем клетки с наличием 15N и 14N растут синхронно, то после завершения первого и второго циклов репликации молекул ДНК соотношение получившихся плотностей (см числа на рисунке сверху) оказывается 1 1 Расщепление в конце второго цикла на два вида ДНК — средней плотности (одна из нитей не содержит тяжелого изотопа 15N) или меньшей плотности (обе нити не содержат 15N) подтверждает правильность модели полуконсервативной репликации ДНК

В действительности, конечно, процесс репликации куда сложнее, чем показано на рис. 1.7. С одной стороны, разделение цепей молекулы ДНК с образованием репликационной вилки из-за спирального закручивания цепей ДНК вызывает быстрое вращение вокруг ее оси — до 300 об/с. Хаотическому образованию узлов и разрывов двойной спирали при этом препятствуют особые релаксационные ферменты (топоизомеразы I): они вносят цепочечные разрывы и затем ликвидируют их. Благодаря этому временно возникают точки свободного вращения, в которых удается выровнять торсионные напряжения и избежать опасных разрывов ДНК, не вовлекая во вращение соседние части молекулы.
С другой стороны, обе нити двойной спирали антипараллельны, так что в месте репликационной вилки удлиняться должны как 3'-, так и 5'-концы. Однако ДНК- полимеразы (и РНК-полимеразы) из-за особого механизма катализируемых реакций способны удлинять цепочки исключительно с 3''-конца. Фактически постепенно удлиняется только нить со стороны 3'- конца (главная цепь; англ, leading strand), а на обратной цепи (англ, lagging strand) вторая цепь синтезируется кусочками — прерывисто — и уже затем с помощью лигаз эти новые отрезки связываются ковалентными связями (полунепрерывная репликация). Лигазы — ферменты, которые могут связывать ковалентно свободные 3'- концы со свободными 5'-концами. Они играют важную роль в реакциях репарации (восстановления) поврежденных участков цепей ДНК (рис. 1.8), но участвуют также и в репликации. У организмов с дефектными лигазами часть последовательностей в обратной цепи остаются не связанными, и могут возникать изолированные фрагменты Оказаки, названные так в честь исследователя, открывшего это явление.
Рис. 1.8. Репарация двойной цепи ДНК после индуцированного УФ образования тимидиновых димеров
1—определение дефектного участка, разделение нитей ДНК и вырезание дефектного участка. В этом принимают участие многие белки, среди которых у эукариот есть и общий транскрипционный фактор — ТFLIН (см 7.2 2 2), работающий также и при синтезе мРНК Это объясняет, почему повреждения ДНК в транскрибируемой области (= переписываемой в РНК) восстанавливаются быстрее, чем в участке не транскрибируемого гена, 2—заполнение поврежденного участка в направлении от свободного З'-конца с помощью ДНК-полимеразы, 3—связывание свободного З'-конца с 5'-фосфатом исходной цепи с помощью ДНК-лигазы вновь создает полноценную цепь ДНК
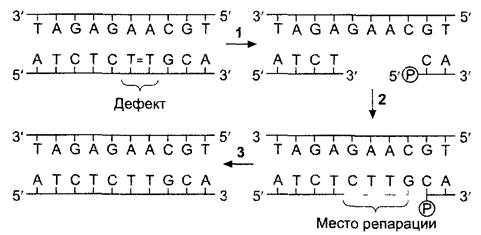
ДНК-полимеразы в отличие от РНК- полимераз могут удлинять только уже имеющиеся З'-концы. Поэтому им требуется — кроме матрицы в виде одноцепочечной ДНК — также и праймер (от англ, primer — затравка, пусковое устройство), чтобы запустить синтез ДНК. В качестве праймера на обратной цепи образуются на равных расстояниях — соответствующих длине фрагмента Оказаки — с помощью особой РНК-полимеразы (праймазы) короткие последовательности РНК, на З'-концах которых затем вступают в работу ДНК- полимеразы. В дальнейшем праймеры разрушаются, соответствующие бреши в последовательностях закрываются с помощью репарационных полимераз и лигаз.
Молекулярная структура репликационной вилки в настоящее время представляется такой, как схематически показано на рис. 1.9. В принципе эта модель относится к репликации ДНК у прокариот, а также в митохондриях, пластидах и клеточных ядрах эукариот, где всегда встречается как раз двухцепочечная ДНК. Однако в то время как сравнительно короткие кольцевые ДНК органелл и прокариот (см. 7.2.1) имеют только одну стартовую точку репликации — точку инициации репликации (origin), — от которой в противоположных направлениях от репликационной вилки процесс идет вдоль всего кольца ДНК, в линейных ДНК хромосом эукариот длиной от сантиметра до дециметра имеется много стартовых точек репликации; иначе сложный процесс удвоения хромосомы продолжался бы недели и месяцы, несмотря на высокую эффективность полимераз, которые при копировании наследственного материала должны работать с очень большой точностью. Синтезированная с одной точки инициации репликации последовательность нуклеотидов называется репликоном. Кольцевые ДНК бактерий и клеточных органелл эукариот являются монорепликонными1, тогда как линейные ДНК эукариотических хромосом полирепликонны. Другие отличия в удвоении эукариотических хромосом по сравнению с бактериальной репликацией (рис. 1.9) относятся к ДНК- полимеразе: вместо прокариотической ДНК-полимеразы III работающая у эукариот ДНК полимераза а обладает собственной праймазной активностью, что позволяет синтезировать РНК-праймеры как на «главной», так и на «обратной» цепи. ДНК полимераза а, однако, не способна к синтезу длинных участков ДНК и заменяется главным репликационным ферментом (ДНК полимеразой δ), если праймер был удлинен примерно на 30 нуклеотидов.
1 Вопрос о числе репликонов в митохондриальном геноме до сих пор не решен окончательно. Возможно, в крупных митохондриальных геномах со сложной многокольцевой структурой есть несколько репликонов. — Примеч. ред.
Рис. 1.9. Репликация ДНК у Escherichia соli; репликационная вилка, движущаяся в направлении стрелки (по А. Kornberg, из Н. Kletnig и U. Maier): А—раскручивание двойной спирали ДНК с помощью специфических ферментов геликаз с временной стабилизацией отдельных цепей особым белком (SSB). На продолжающемся вперед (на рисунке вниз) конце нити сразу начинается непрерывный синтез новой комплементарной [антипараллельной] цепи (конец стрелки: растущий З'-кoнец) с помощью ДНК- полимеразы III. На обратной цепи (вверху) полимераза, напротив, работает в противоположном направлении (однако также 5' → З'); она удлиняет З'-конец РНК-праймеров, которые в свою очередь синтезируются праймазами (особыми ферментами праймосом) на равных расстояниях (прерывистая репликация). РНК-праймер наконец перестраивается, бреши заполняются благодаря репаративному синтезу (ДНК- полимераза I) и сохраняющиеся разрывы одиночной цепи связываются ковалентно с помощью лигазы. В—гипотетическая модель «реплисомы», в которой все ферменты и белковые факторы аппарата репликации объединены в комплекс. Антипараллельность исходных родительских цепей ДНК локально сохраняется благодаря образованию петель на обратной цепи
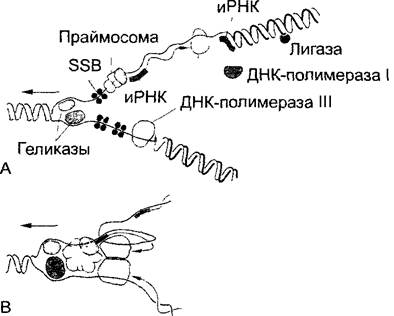
Синтез ДНК идет примерно в 5 раз быстрее, чем синтез РНК во время транскрипции (см. 7.2.2.2). Так как во время репликации переписываются гены, предполагают, что ДНК-полимеразы приостанавливают свою работу, пока не завершится синтез молекулы РНК.
1.2.4. Рибонуклеиновые кислоты (РНК, RNA)
В отличие от ДНК, которая в виде одной нити встречается лишь у некоторых вирусов и фагов, а во всех иных случаях представлена двойной нитью, молекулы РНК чаше бывают однонитевыми. За счет внутримолекулярных спариваний азотистых оснований (рис. 1.10, А, С) образуются стабилизированные вторичные структуры и, часто в результате ассоциации с белками, третичные структуры. Поэтому имеется несколько вариантов структуры РНК и рибонуклеолротеиновых комплексов (РНП, RNP). Исключением являются лишь транспортные РНК (тРНК, tRNA), которые никогда не связываются с белками. Единственная цепь РНК легко разрушается ферментами нуклеазами, поэтому внутримолекулярные спаривания оснований и ассоциация с белками повышают устойчивость молекулы. К тому же вторичные и третичные структуры важны для функционирования молекул РНК. Итак, как мы уже упоминали, имеются следующие отличия от ДНК:
✵ наличие рибозы вместо 2-дезоксирибозы;
✵ наличие урацила вместо тимина;
✵ значительно меньшая величина молекул РНК по сравнению с ДНК.
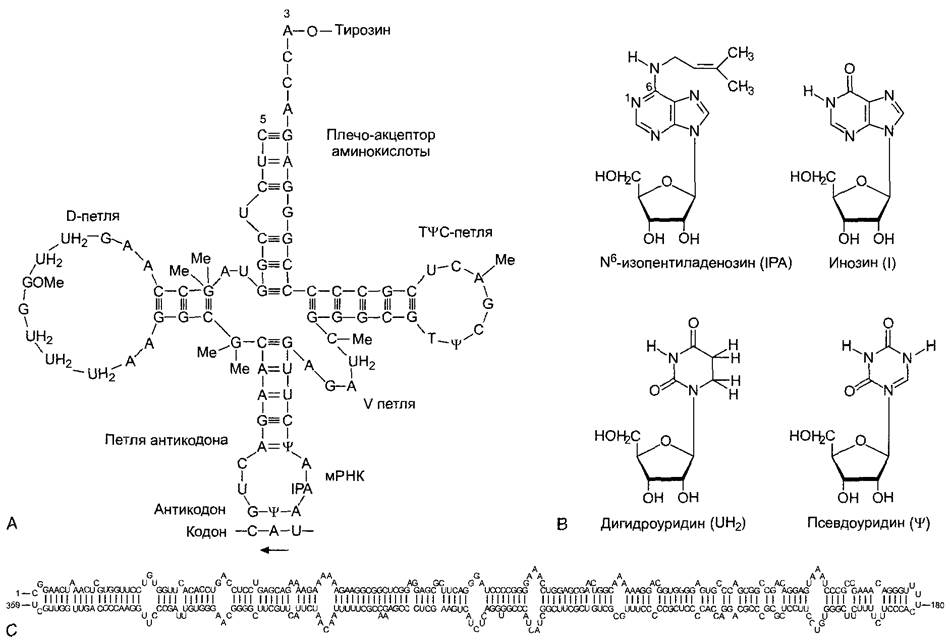
Рис. 1.10. Рибонуклеиновые кислоты
А—структура «клеверного листа» в молекуле тРНК, на примере переносящей аминокислоту тирозин (tRNATyr) дрожжей Структура стабилизируется благодаря особой конформации молекулы,свернутой таким образом, что внутри каждой из трех петель возникают водородные связи между парными азотистыми основаниями, а в наружных участках петель находятся неспаренные основания Многие основания тРНК модифицированы, некоторые важные модификации представлены на В Кроме того, имеются метилированные основания Триплет оснований, так называемый антикодон, связывается с комплементарным ему триплетом оснований мРНК — кодоном Последовательностью триплетов в мРНК определяется (благодаря взаимодействию кодон—антикодон) последовательность аминокислот в синтезируемом белке (генетический код, см 7 3 1 1, синтез белков, см 7 3 12) D-петля названа так по встречающемуся здесь вместо урацила дигидроурацилу (UH2), Т![]() С-петля названа по всегда присутствующей последовательности оснований 5'-Т-
С-петля названа по всегда присутствующей последовательности оснований 5'-Т-![]() - С-3' Разные тРНК имеют V-петлю варьирующей величины (от Variable) Петля, принимающая аминокислоту, и петля антикодона важны для опознавания соответствующей тРНК и принадлежащей ей аминокислоты с помощью аминоацил-тРНК-синтетаз Петля антикодона в то же время располагает антикодон так, что в рибосоме он может вступать в комплементарную связь с кодоном мРНК Для связывания тРНК с рибосомой особое значение, как предполагают, имеют петли Т
- С-3' Разные тРНК имеют V-петлю варьирующей величины (от Variable) Петля, принимающая аминокислоту, и петля антикодона важны для опознавания соответствующей тРНК и принадлежащей ей аминокислоты с помощью аминоацил-тРНК-синтетаз Петля антикодона в то же время располагает антикодон так, что в рибосоме он может вступать в комплементарную связь с кодоном мРНК Для связывания тРНК с рибосомой особое значение, как предполагают, имеют петли Т![]() С и D С — пример замкнутой в кольцо молекулы РНК — вироид, вызывающий веретеновидность клубней картофеля (PSTV, Potato Spindle Tuber Viroid) Он состоит из 359 нуклеотидов, замкнутых в кольцо ковалентной связью Его структура стабилизирована благодаря внутримолекулярным связям азотистых оснований, которые для наглядности показаны одинарными линиями
С и D С — пример замкнутой в кольцо молекулы РНК — вироид, вызывающий веретеновидность клубней картофеля (PSTV, Potato Spindle Tuber Viroid) Он состоит из 359 нуклеотидов, замкнутых в кольцо ковалентной связью Его структура стабилизирована благодаря внутримолекулярным связям азотистых оснований, которые для наглядности показаны одинарными линиями
Разнообразие структуры определяет и функциональное многообразие различных видов РНК (табл. 1.2). Большинство коротко живущих молекул информационной, или матричной, РНК (иРНК, мРНК) (messenger-RNA) представляют собой переписанные участки генов (см. 7.2.2). Они служат матрицей в процессе трансляции на рибосоме при последовательном присоединении к ней стабильных транспортных РНК, несущих определенные аминокислоты, и, таким образом, определяют последовательность аминокислот в белке. Также стабильные рибосомальные РНК принимают участие в построении рибосом. Кроме того, в белковых комплексах встречаются малые цитоплазматические РНК (мцРНК, scRNA; англ, small cytoplasmic), например, как составная часть сигнального комплекса SRP1, который играет определенную роль в трансляции (см. 7.3.1.4). Малые ядерные РНК (мяРНК, snRNA; англ, small nuclear) участвуют в процессинге транскрибированных мРНК итРНК (см. 1.2.2.2).
1 SRP — signal recognition particle (англ.) — частица, распознающая сигнал. — Примеч. ред.
Таблица 1.2. Приблизительные размеры и функции 3 видов РНК по сравнению с ДНК
Нуклеиновая кислота |
Размер, н. п. |
Функция |
ДНК |
Свыше 100 млн |
Хранение информации (гены) |
мРНК |
От нескольких сот до 10 000 и более |
Переписанный участок гена переносит к рибосомам информацию для синтеза белка |
рРНК |
4 вида (в клетках эукариот): около 120, 150, 1 700, 3 500 |
Обеспечение структуры и функции рибосом |
тРНК |
80-90 |
Переносят аминокислоты к рибосомам |
1.2.5. Вирусы, фаги, вироиды
Вирусы — облигатные паразиты эукариот, фаги — такие же паразиты прокариот. Речь идет о частицах, которые слишком просты, чтобы размножаться самостоятельно, и используют для этого метаболическую активность живых клеток. Их нуклеиновые кислоты — двухцепочечные или одноцепочечные ДНК — несут генетическую информацию; они могут мутировать и при подходящих обстоятельствах также и рекомбинировать. Именно фаги сыграли важную роль как модельные организмы в развитии современной генетики.
У вирусов и фагов нуклеиновые кислоты образуют комплекс с белками. Молекулы белков обычно образуют высоко симметричную структуру оболочки, называемую капсида (см. рис. 1.17). Она выполняет структурную и защитную функции и важна при инфицировании новых клеток хозяина. У «сложных» вирусов имеется дополнительно рыхлая мембранная оболочка. Она образована клеточными мембранами последней клетки-хозяина, но включает также и специфические вирусные гликопротеины.
Вироиды не имеют таких оболочек, а являются очень маленькими, содержащими всего 250 — 370 оснований, кольцевыми или палочковидными свободными молекулами РНК, которые могут действовать как возбудители болезней растений (см. рис. 1.10, С). Веретеновидность клубней картофеля и болезнь каданг-каданг кокосовых пальм вызываются именно вироидами. Вироиды попадают в новое растение-хозяин через поврежденные клетки и нарушают их метаболизм, вмешиваясь в процесс образования молекул РНК хозяина.