БОТАНИКА ТОМ 3 - ЭВОЛЮЦИЯ И СИСТЕМАТИКА - 2007
11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
11.2. Бактерии, грибы, растения
Второе подцарство: Мyxobionta
Отличающие этот таксон плазмодии возникают путем слияния миксофлагеллят или миксамеб или же развиваются из отдельных клеток без предшествующего полового процесса. В жизненном цикле встречаются подвижные стадии со жгутиками. Клеточные стенки, насколько они различимы на определенных стадиях развития, построены из галактозамина и целлюлозы.
Первый отдел: Myxomycota
I. Класс: миксомицеты (Myxomycètes). Вегетативная (= соматическая) фаза представляет собой диплоидный, многоядерный, не подразделенный на отдельные клетки настоящий плазмодий (см. рис. 2.9; 11.16), питающийся фаготрофно. Из плазмодиев развиваются плодовые тела, причем часть плазмы отвердевает и создает характерные структуры, тогда как другая часть, содержащая ядра, превращается в мейоспоры. Последние имеют клеточную стенку, состоящую не менее чем из двух слоев и, согласно новейшим исследованиям, не содержащую в своем составе ни целлюлозы, ни хитина, а лишь преимущественно полимерный галактозамин. В качестве запасного питательного вещества образуется гликоген. Плазмодии и особенно плодовые тела часто окрашены в яркие цвета. Химическая структура пигментов изучена лишь отчасти; она иная, чем у грибов.
Рис. 11.16. Myxomycota. Край плазмодия у Badhamia utricularis (2х)
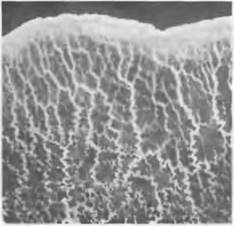
Жизненный цикл. Споры прорастают в воде или на влажном субстрате. Всхожесть их часто сохраняется очень долго. Так, удалось прорастить споры из гербарного образца, хранившегося свыше 70 лет. При этом споры высвобождают или одноядерные, голые амебоидные подвижные миксамебы. или миксофлагелляты, имеющие жгутики (рис. 11.17, А). Миксофлагелляты чаще всего двужгутиковые, причем один жгутик часто сильно редуцирован или полностью отсутствует. В последнем случае, по крайней мере, имеется второй, утративший свою функцию, блефаропласт. Подвижные клетки могут за счет утраты жгутиков превращаться в миксамебы. Последние размножаются, как и подвижные клетки, путем деления. Миксамебы или миксофлагелляты сливаются попарно друг с другом в амебозиготу (или имеющую жгутики планозиготу), в которой затем сливаются ядра (плазмогамия, затем кариогамия). Получающаяся диплоидная структура претерпевает многочисленные митотические ядерные деления и развивается в крупный многоядерный плазмодий (см. рис. 11.16). Такие плазмодии могут в свою очередь сливаться между собой. Митозы закрытые и происходят у всех ядер одного плазмодия синхронно. В плазмодиях плазма находится в интенсивном движении. Плазмодии развиваются при большой влажности воздуха в лесной почве, в соломе, среди травы, мхов или в гнилой древесине. Позже они изменяют форму и медленно выползают на поверхность субстрата. Клеточные стенки отсутствуют как на поверхности, так и внутри плазмодия. Его передняя часть (см. рис. 11.16) состоит из более плотной плазмы; ближе к задней части он часто выглядит как разделенный на сеть из отдельных тяжей. Диаметр плазмодия у некоторых видов может превышать 20 см (например, Fuligo, Brefeldid).
Образование плодовых тел начинается при определенных, еще недостаточно изученных экзогенных условиях (исчерпание субстрата, свет, температура, pH); возможно, решающую роль играют также эндогенные факторы. Перед этим меняется поведение плазмодия: он выползает из влажного субстрата к свету и превращается при сильной потере воды в многочисленные спорокарпы (спорангии, см. рис. 11.17, D). Эти плодовые тела имеют внешнюю, часто содержащую известь стенку — перидий, а нередко также и ножку, которая может продолжаться внутрь спорокарпа как колонка, а кроме того, нередко и систему нитей— капиллиций. Структуры формируются из затвердевшей безъядерной плазмы, которая не идет на образование спор. Возникновение капиллиция, очевидно, начинается с отложения строительного материала в особых пузырьках. Содержащая ядра плазма путем разделения образует одноядерные, сначала диплоидные споры. В результате последующего мейоза в каждой споре возникает 4 гаплоидных ядра, из которых три ядра исчезают и остается только одно. При созревании плодового тела перидий разрывается, и споры выбрасываются из сплетения капиллиция. У некоторых видов капиллиций, подобно элатерам печеночников, за счет гигроскопических изменений формы способствует освобождению спорокарпа. В жизненном цикле гаплоидны миксофлагелляты и не копулировавшие миксамебы, диплоидными являются плазмодии, плодовые тела и молодые споры, а зрелые споры опять гаплоидны. Таким образом, диплоидная фаза в ходе развития преобладает.
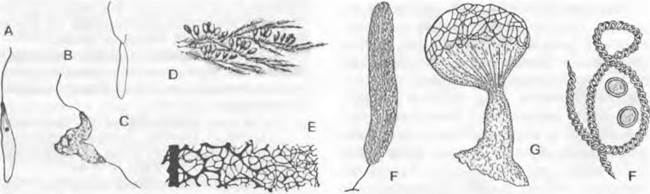
Рис. 11.17. Myxomycota: А — С — миксофлагелляты: А (и В) — короткий жгутик не изображен, В — копуляция (1 500х), С — показаны длинный и короткий жгутик; D — Leocarpus fragilis,многочисленные плодовые тела на мхе (нат. величина); Е — Comatricha typhoides, часть капиллиция (180х) F — Stemonitis fusca, плодовое тело (5х); G — Cribraria rufa, плодовое тело (З0х); Н — Trichia varia; нить капиллиция и споры (З00х)
Бокс 11.2. Типы организации слизевиков
Организмы обоих подцарств — Аcrasiobionta и Мyxobionta — называются слизевиками. Слизевики отличаются вегетативным телом, представляющим собой лишенную клеточной стенки, многоядерную амебоидную подвижную плазменную массу — плазмодий. Он возникает;
✵ как агрегационный плазмодий; миксамебы сползаются в плазменные скопления, не теряя своей индивидуальности;
✵ как настоящий плазмодий: миксамебы или миксофлагеллаты должны слиться между собой, образуя диплоидный многоядерный плазмодий.
Еще одна возможность— возникновение плазмодия бесполым путем из одной-единственной клетки с делением ядер (но без клеточного деления).
Имеющиеся в жизненном цикле плазмодии различны, таким образом, по своей природе и аналогичны прокариотическим плазмодиям миксобактерий (Мухоbacteriales). Размножение осуществляется спорами, которые возникают в особых плодовых телах (см. рис. 11.17, G), если это не паразитические формы, обитающие внутри хозяина. У подвижных стадий имеется два гладких жгутика чаще всего неравной длины, реже один жгутик редуцирован. Между разными отделами слизевиков близкого эволюционного родства нет. Очень своеобразные по своей форме и образу жизни слизевики имеют много общих признаков с представителями выделяемой зоологами группы простейших, с которыми они близкородственны, но отличаются образованием плодовых тел и спор. Сходство состоит, в частности, в следующем;
✵ гетеротрофность — большинство форм питается, как и животные, фаготрофно, поглощая целые твердые частицы;
✵ наличие амебоидных стадий, входящих в жизненный цикл;
✵ отсутствие клеточных стенок, по крайней мере, в вегетативных фазах развития.
Это сходство с простейшими побудило многих исследователей к тому, чтобы относить слизевиков под названием Муcetozoa к царству животных.
Питание плазмодиев или предшествующих им одноклеточных стадий осуществляется в природе, вероятно, всегда за счет поглощения различных микроорганизмов, таких, как бактерии, простейшие, споры, дрожжевые клетки, гифы грибов и т. д. Частицы пищи заключаются в пищеварительные вакуоли и ферментативно перевариваются; непереваренные остатки через некоторое время выбрасываются. В культуре большинство видов может поддерживаться только тогда, когда им скармливают живые микроорганизмы (например, бактерии). Некоторые виды могут также поддерживаться в чисто сапротрофных условиях на питательных средах определенного состава.
В условиях культуры плазмодии некоторых видов могут захватывать и накапливать внутри себя одноклеточные зеленые водоросли, не переваривая их. Становясь таким образом зелеными, плазмодии приобретают фотосинтетическую активность. Хотя зеленые водоросли и попадают в плодовые тела, в естественных условиях такой симбиоз не может поддерживаться из поколения в поколение.
Систематика. Подразделение миксомицетов основывается прежде всего на различиях во внешнем виде плодовых тел. В более примитивных группах отсутствует капиллиций. Эволюция должна была идти также от сидячих плодовых тел к плодовым телам на ножках и от одиночных плодовых тел к комплексным. Примерно 500 известных видов подразделяются на следующие порядки.
1. Порядок: Сеratiomyxales. Спорообразование осуществляется экзогенно. На поверхности имеющего вид колонки плодового тела отчленяются на ножках споры (вероятно, односпоровые спорокарпы). Каждая спора дает при прорастании плазматический вырост с 4 гаплоидными ядрами, из которых после митоза образуется 8 гаплоидных подвижных клеток. В порядок входит только один род Сеratiomyxa с одним видом, представленным многочисленными формами и встречающимся по всему земному шару на гнилой древесине.
Во всех последующих порядках (2 — 6) споры образуются эндогенно внутри плодовых тел.
Плодовые тела у следующих 4 порядков (2 — 5) формируются из полукруглых выпуклостей плазмодия, которые сначала соединены с ним и друг с другом плазматическими тяжами, однако позже становятся изолированными. Прилегающий к субстрату, составляющий основу слой
плазмодия («гипоталлус») или вообще не сохраняется при созревании плодовых тел, или от него остается только слизистая кожица.
2. Порядок: Liceales. В противоположность дальнейшим порядкам (3 — 6) здесь отсутствуют капиллиций и колонка (например, Lycogala, Cribraria. рис. 11.17, G).
3. Порядок: Echinosteliales. Имеется колонка.
4. Порядок: Trichiales. Колонка отсутствует. Капиллиций состоит из нитей со свободными концами, каку Trichia' (рис. 11.17, Н). У всех трех предыдущих порядков (2 — 4) споровая масса светлых тонов.
5. Порядок: Physarales. Споровая масса черная или разных тонов — от темно-фиолетовых до ржавых. На перидии и часто также на капиллинии видны белые отложения извести (например, Didymium). Сюда относятся также Leocarpus (рис. 11.17, D); Badhamia (рис. 11.16); Fuligo séptica с комплексным плодовым телом (эталием).
6. Порядок: Stemonitales. Плодовые тела, имеющие размер от 0,5 до 1 — 2 см, развиваются на «гипоталлусе». Внутри они дифференцированы на колонку, капиллиций и споры. У Stemonitis капиллиций образует по периферии замкнутую сеть, у Comatricha нити имеют свободные концы (рис. 11.7, Е). Lamproderma отличается перидием с металлическим блеском, у Brefeldia плоское, крупное плодовое тело составлено из многих спорокарпов (эталий).
Практическое значение миксомицетов невелико. Большую популярность как тест-обьект при физиологических и биохимических исследованиях получил Physarum (Physarales).
В следующем классе плазмодии возникают без предшествующего полового процесса, непосредственно из одиночных клеток (также и без агрегации плазмодиев).
II. Класс: протостелиомицеты (Protosteliomycetes). Многоядерные сетчатые плазмодии образуются из клеток со жгутиками или без них. На тонких ножках экзогенно отчленяются от одной до четырех спор.
Второй отдел: плазмодиофоромицеты (Plasmodiophoromycota)
Этот отдел отличается от всех ранее рассмотренных слизевиков наличием хитиновых клеточных стенок, а также одной особенностью ядерного деления: в метафазе хроматиновые массы располагаются перпендикулярно к обеим сторонам крупного, несколько вытянутого ядрышка, так что возникает крестообразная фигура внутри ядерной мембраны. Остается под вопросом, правомерно ли рассматривать Plasmodiopho- romycota как производные миксомицетов, перешедшие к эндопаразитизму. Правда, у этих двух групп имеются сходные зооспоры с двумя жгутиками неравной длины. В цикле развития участвуют гаплоидные и диплоидные плазмодии; у миксомицетов они всег да диплоидные, у протостелиомицетов — гаплоидные.
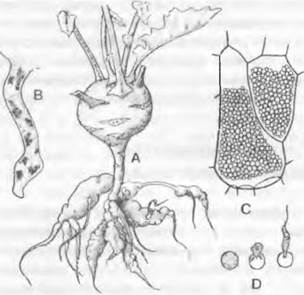
Известный представитель единственного класса Plasmodiophoromycetes — Plasmodiophom brassicae — возбудитель килы капусты1 (рис. 11.18).
1 Более корректно называть эту болезнь «кила крестоцветных», поскольку поражается не только капуста. — Примеч. пер.
Жизненный цикл. Перезимовавшие покоящиеся споры (гипноспоры) паразита прорастают весной в почве двужгутиковыми гаплоидными зооспорами, которые после утраты жгутиков внедряются, как амебоиды, в корневые волоски молодых растений капусты. Здесь каждая внедрившаяся амеба образует многоядерный гаплоидный плазмодий (рис. 11.18, В). Он может распадаться на многоядерные частицы, которые в свою очередь (после разрушения стенок в тканях хозяина) перемещаются дальше от клетки к клетке и таким образом быстро расширяют очаг инфекции. Позже, после распада плазмодия на сначала одноядерные, потом многоядерные фрагменты, возникают многоядерные гаметангии. Последние распадаются на двужгутиковые гаметы, число которых соответствует числу ядер. После разрушения ткани хозяина они высвобождаются и копулируют между собой в почве.
Диплоидные планозиготы после отбрасывания жгутиков снова внедряются в корни уже развившегося к тому времени растения капусты (теперь уже не исключительно через корневые волоски), где они разрастаются в многоядерные протопласты без клеточных стенок — диплоидные плазмодии. Растение-хозяин отвечает на это образованием опухолей (рис. 11.18. А). Затем после мейоза в клетках растения-хозяина возникают покоящиеся толстостенные гаплоидные мейо- споры (покоящиеся споры, гипноспоры), которые зимуют вместе с пораженным растением и весной, после сгнивания ткани опухоли, снова попадают в почву.
Рис. 11.18. Plasmodiophoromycota, Plasmodio- phora brassicae: A — кила на корнях растения кольраби (1/Зх); В — плазмодии в корневом волоске (З00х); С — клетки коры корня со спорами (520х); D — прорастание спор (1 240х)
Зооспоры и гаметы несут на переднем конце два очень различающихся по длине жгутика без мастигонем. Смена гаплоидного и диплоидного плазмодиев соответствует смене генераций, однако места кариогамии и мейоза в жизненном цикле еще окончательно не выяснены.
Виды некоторых родственных родов (например, Polymyxa) — паразиты разных наземных и водных растений, которые образуют похожие разрастания органов (60 облигатно эндопаразитических видов, поражающих сосудистые растения, водоросли и грибы).
Обзор слизевиков. Слизевики (Acrasiobionta и Myxobionta; около 600 видов) находятся в основании эволюционного развития гетеротрофных эукариот. Протостелиомицеты и миксомицеты возникли, скорее всего, от бесцветных жгутиковых, а акразиомицеты — от каких-то амеб. Эти группы из-за особенностей морфологии и цикла развития занимают очень обособленное положение («Mycetozoa»). Что касается плазмодиофоромицетов, то остается неясным, возник ли у них плазмодий исходно или только вторично в связи с эндосимбиотическим образом жизни. Во втором случае следовало бы присоединить этот класс к рассматриваемым далее грибам и там сближать с группами, характеризующимися сходными признаками. Правда, там нигде не встречаются подвижные стадии с двумя гладкими жгутиками неравной длины, как у миксомицетов, протостелиомицетов и плазмодиофоромицетов.