БОТАНИКА ТОМ 3 - ЭВОЛЮЦИЯ И СИСТЕМАТИКА - 2007
11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
11.2. Бактерии, грибы, растения
Третий подотдел: папоротникообразные, птеридофиты (Рteridophytina)
Папоротникообразные, как и мхи, — это прежде всего растения, приспособившиеся к наземному образу жизни, однако их ткани и органы значительно более дифференцированы функционально.
В жизненном цикле современных папоротникообразных доминирует спорофит (рис. 11.128, Н). Он представляет собой самостоятельное зеленое растение, и у плаунов, хвощей и настоящих папоротников дифференцирован на стебель, листья и корни. Вымершие первопапоротники, напротив, были устроены чаще всего из исключительно однотипных, лишенных листьев вильчатых осей (теломов; см. рис. 11.130, А, В). У них. как и у современных псилотовых. не было настоящих корней.
Гаплоидный гаметофит у папоротникообразных называют заростком (рис. 11.128, А). Он обычно живет лишь несколько недель, достигает не более нескольких сантиметров в диаметре и по облику часто напоминает простой галломный печеночник. В типичном случае — отклонения от правила очень разнообразны — он представляет собой простой зеленый таллом, прикрепляющийся к почве одноклеточными нитевидными ризоидами. На нем в большом числе возникают антеридии и архегонии. Оплодотворение, как и у мхов, возможно только в воде, т. е. при увлажнении заростков.
После оплодотворения из зиготы развивается диплоидное поколение — спорофит (рис. 11.128, Н: 3, 4), который у папоротников, однако, выглядит совсем иначе и гораздо более развит, чем у мхов. Пожалуй, лишь начальные стадии его развития протекают сходно со мхами. Заросток у большинства видов вскоре отмирает (при подавлении оплодотворения он может прожить в течение нескольких лет), однако проросток спорофита вырастает в самостоятельное многолетнее растение с корнями, стеблем и листьями: собственно, папоротникообразное растение (см. рис. 11.128, Н: 4Б; 11.148, А; 11.150; 11.152, А). Спорофит папоротника, таким образом, — настоящее листостебельное растение.
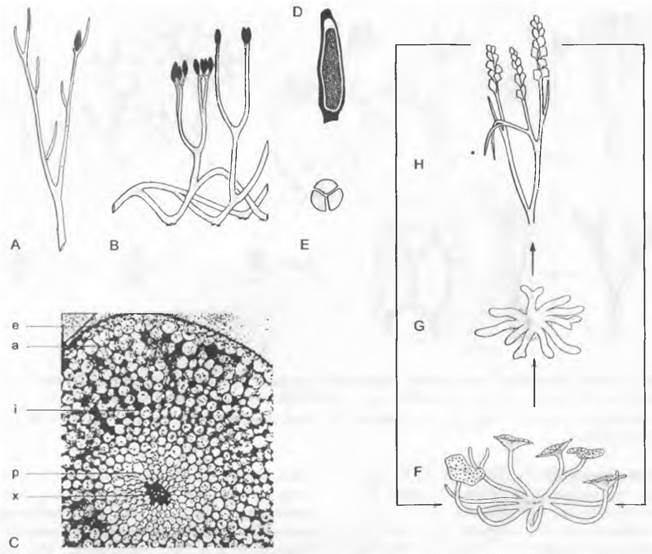
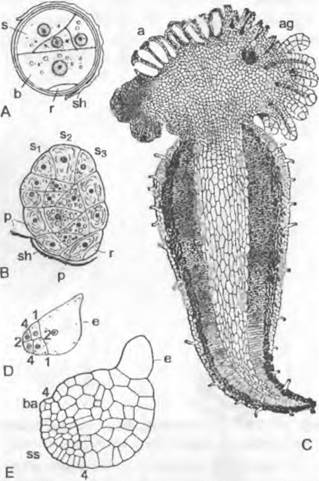
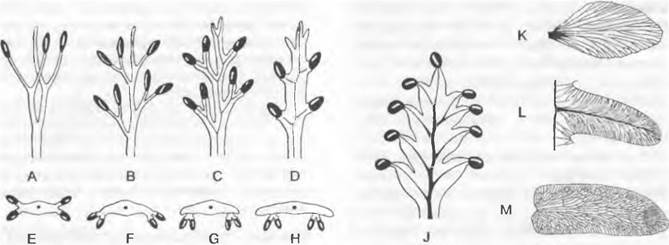
Рис. 11.128. Pteridophytina, Pteridopsida:
A, В — Dryoptens filix-mas: A — заросток (снизу) с архегониями (аг), антеридиями (аn) и ризоидами (г), В — заросток с молодым спорофитом (5х); С — G — развитие спорангия папоротника (С — Е — Asplénium, 300х; F, G — Polypodium, 200x ): С — первые деления возникающего из одной эпидермальной клетки зачатка, D — разделение на поверхностный слой — стенку спорангия (wa) и центральную клетку (as) — археспорий, которая уже отделила клетку тапетума (t), Е — археспорий, дифференци ровался на клетки тапетума и спорогенную ткань (sg), F — у клеток кольца стенки спорангия (wa) утолщены стенки; клетки тапетума лизировались, материнские клетки спор (sm) образовали тетрады спор (s), G — зрелый спорангий со спорами; Н — схема развития папоротника: светлые линии — гаплоидная фаза, темные линии — диплоидная фаза, 1 — спора, 2 — заросток с ♀ и ♂ гаметангиями, 3 — заросток с молодым спорофитом, 4 — спорофит (сильно уменьшен) с сорусами спорангиев, 5 — незрелый одиночный спорангий (сильно увеличен) из соруса, 6 — зрелый спорангий с тетрадами спор; 7 — споры (s); b — первый лист; w — корень; 1 — 5 на С, D — последовательно образующиеся стенки клеток; G — гаметофит; S — спорофит; R! — редукционное деление

Теломная теория. Органы, типичные для листостебельных растений, очевидно, возникли из лишенных листьев вильчатых теломов путем следующих основных процессов: перевершинивания, планации, срастания, редукции и изгибания (заворачивания).
Бокс 11.9. Сосудистые растения
Папоротникообразные и семенные растения имеют в стеблях, листьях и корнях настоящие проводящие пучки (а не только проводящие тяжи, см. ниже), которые служат для транспорта веществ (воды с минеральными веществами, продуктов ассимиляции). Поэтому их называют сосудистыми растениями (Тracheophyta) в отличие от мхов, у которых такие проводящие пучки отсутствуют1. Правда, у мхов с внутренней системой проведения воды уже встречаются проводящие тяжи, которые напоминают исходные типы проводящих пучков. Однако эти тяжи менее резко отграничены от окружающей ткани, а в клеточных стенках проводящих элементов не происходит отложения лигнина. Соответственно у проводящих воду элементов мхов нет утолщения стенок по типу трахеид и сосудов сосудистых растений. Совокупность проводящих пучков в стебле сосудистых растений называют стелой. В ходе все большего приспособления сосудистых растений к наземной жизни стелы становились все более развитыми в отношении проведения веществ и придания прочности стебелю (прото-, актино-, плекто-, поли-, сифоно-, эвстела, атактостела; см. Стелярная теория, бокс 4.3).
1 Сосудистые растения получили это название потому, что водопроводящие элементы у них трахеальные — трахеиды и сосуды, тогда как у мхов это гидроиды (или их нет совсем). — Примеч. ред.
Дифференциация и разделение функций исходно однородных теломов, из которых еще состояло тело первичных папоротников, на несущие основные и боковые оси должны были произойти в результате перевершинивания (рис. 11.129, А, В). При этом лидирующая основная ось растет более интенсивно, чем обгоняемые ею в росте сестринские оси, которые становятся ее боковыми придатками (В) и все больше берут на себя функцию ассимиляции. В ходе планации боковые оси стали ветвиться в горизонтальной плоскости (С, D). Путем конгенитального срастания эти теломы, расположенные уже в одной плоскости, превратились в плоские листовидные боковые органы (D, G). Так, по-видимому, появились крупные сложно-расчлененные листья с первоначально дихотомическим жилкованием (макро-, или мегафиллы). Располагающиеся в трехмерном объеме талломы могут срастаться между собой, в результате чего возникает более толстая паренхима- тическая ось, в которой теперь проходит уже не единственный проводящий пучок (протостела) (рис. 11.130, С), а два или более (рис. 11.129, Н — К). Благодаря этому прочность оси значительно повышается. Путем редукции (F, G), как можно полагать, могли возникнуть мелкие, более или менее шиловидные листья с одной жилкой (микрофиллы). Однако не вполне ясно, не имеем ли мы здесь дело с выростами осей, т. е. органами «sui generis», которые не могут быть выведены из теломов. Процесс изгибания можно проследить, например, на несущих спорангии осях хвощевидных (L, М) (см. рис. 11.141, F).
Рис. 11.129. Схема пяти элементарных процессов, которые согласно теломной теории привели к появлению побега: А, В — перевершинивание; С, D — планация; Е — G — редукция; Н — К — срастание (также D, N); L — N — изгибание

Развитие зародыша. У современных папоротникообразных вскоре после первых клеточных делений оплодотворенной яйцеклетки, как правило, возникают кроме гаустории (ножки) апексы корня, стебля1 и листа, которые у растущего и еще связанного с заростком зародыша (см. рис. 11.128, В; 11.155) развиваются соответственно в первичный корень, первичный стебель2 и первый лист (семядолю). Наличие корня характерно для большинства папоротникообразных. Противоположный «полюсу побега» коней оси проростка можно было бы назвать «корневым полюсом»; однако только у семенных растений из него развивается главный корень (см. рис. 3.1), тогда как у папоротникообразных первый корень возникает сбоку на оси как эндогенное придаточное образование3 (см. рис. 11.155, В: w). Проросток папоротникообразных, таким образом, не биполярный, как у семенных растений, а униполярный4. Зародышевый корень (см. рис. 11.128, В: w) быстро отмирает, но в дальнейшем возникают многочисленные придаточные корни на побегах (первичная гоморизия, см. 4.4.1).
1 Правильно: апекс побега. — Примеч. ред.
2 Первичный побег. — Примеч. ред.
3 Главный корень семенных растений образуется под подвеском и, строго говоря, эндогенный. — Примеч. ред.
4 Зародыш подавляющего большинства папоротникообразных такой же биполярный, как и у семенных растений, только искривлен соответственно своему боковому положению относительно питающего его заростка. — Примеч. ред.
Рис. 11.130. Psilophytopsida:
А — Е — Rhyniales: А — Rhyniagwynne-vaughanii, реконструкция (1/4х), В —Aglaophyton (Rhynia) major, реконструкция (1/5х), С — Е — Rhynia: С — поперечный срез телома, показывающий протостелу (50х), D — спорангий, продольный шлиф (2х), Е — тетрада спор (100х); F — Н — Zosterophyllales: Zosterophyllum rhenanum: F — гаметофит (= Sciadophyton),G, H — спорофит: G — молодой, связанный с гаметофитом, Н — взрослый со спорангиями; а — внешняя кора; е — эпидерма; i — внутренняя кора; р — флоэма; х — ксилема
Три основных органа у большинства папоротникообразных нарастают благодаря деятельности апикальных инициалей (см. 3.1.1.1; рис. 3.2, А; 3.6, А). Обильно олиственный побег ветвится вильчато или латерально (но никогда из пазух листьев!). Корни имеют корневой чехлик (см. рис. 3.6, А); боковые корни возникают не из перицикла, а из самого внутреннего слоя первичной коры (см. 4.4.2.2). Листья (по крайней мере у наиболее высокоразвитых папоротников) анатомически в основном подобны листьям семенных растений. Эпидерма надземных частей, как правило, покрыта кутикулой (важная предпосылка для наземного образа жизни на некотором расстоянии от уровня земли!) и имеет устьица (см. 3.2.2.1), однако эпидермальные клетки чаще всего еще содержат хлоропласты. В стеблях, корнях и листьях проходят хорошо развитые, состоящие из флоэмы и ксилемы проводящие пучки, которые в типичном виде в эволюции впервые появляются у папоротникообразных Водопроводящие элементы у них — одревесневшие трахеиды; очень редко (например, у Рteridium) встречаются также сосуды (см. рис. 11.149: t). Особых механических элементов в проводящих пучках еще нет, тем не менее проводящие воду элементы часто укреплены кольчатыми или иными утолщениями своей оболочки (см. рис. 11.149). Преобладают концентрические проводящие пучки (и притом с ксилемой, окруженной флоэмой) в числе одного или нескольких, однако встречаются и другие типы пучков. У папоротникообразных можно проследить общий ряд эволюции проводящих пучков (см. бокс 4.2, рис. А). Одревеснение трахеид повышает их пропускную способность, а заодно и укрепляет побег. Поэтому папоротникообразные в противоположность мхам смогли развиться в обильно расчлененные, иногда даже древовидные растения. Клеточные стенки механической ткани, расположенной вне проводящих пучков, как правило, также содержат лигнин. Наличие корней обеспечивает интенсивное поглощение воды и позволяет образовать более крупные листья, которые производят продукты ассимиляции. Проведение питательных веществ осуществляют вытянутые продольно ситовидные клетки (см. 3.2.4.1). Вторичное утолщение благодаря деятельности камбия у современных папоротникообразных свойственно лишь отдельным представителям, да и то выражено слабо, однако оно было характерно для некоторых вымерших групп папоротникообразных.
Спорангии с мейоспорами (см. рис. 11.128, G, Н: 6) образуются на листьях, лишь в самых примитивных классах — на недифференцированных теломах. Строение спорангиев разное. Листья, несущие спорангии, называют спорофиллами. Они часто устроены более просто, чем ассимилирующие листья (трофофиллы), и располагаются по нескольку в стробиле: такие собрания спорофиллов можно называть «цветками»1. Для лучшего рассеивания спор спорофиллы обычно довольно высоко приподняты над субстратом.
1 Так уже не делают с рубежа XIX — XX вв. — Примеч. ред.
Спорангии содержат археспорий со спорогенной тканью (см. рис. 11.128, Н: 5, Е: sg); ее клетки округляются, отделяются друг от друга и становятся материнскими клетками спор (чаше всего 16), каждая из которых после мейоза дает 4 гаплоидные мейоспоры, часто расположенные в виде тетраэдра.
Вокруг спорогенной ткани находятся, часто в несколько слоев, клетки, примыкающие к стенке спорангия. Они служат для питания спор и в совокупности представляют тапетум (Е: t). Клетки секреторного тапетума выделяют содержимое через стенки. Стенки клеток плазмодиального тапетума лизируются, и высвободившиеся протопласты объединяются в периплазмодий. Последний внедряется затем между молодыми спорами из распадающихся тетрад, питает их, участвует в формировании стенки спор (периспория) и при этом расходуется (F, G).
Споры. Оболочка споры состоит из внутреннего эндоспория и стойкого внешнего экзоспория, на котором откладывается периспорий в виде орнамента различного облика. Споры, окраска которых бывает от буроватой до желтой, почти всегда лишены хлорофилла.
У большинства папоротникообразных (более примитивных) все споры однотипные. При прорастании из них развиваются заростки, на которых чаще всего возникают как антеридии, так и архегонии. Однако у более эволюционно продвинутых форм заростки бывают двудомными2. Это разделение полов в некоторых группах папоротникообразных привело к по явлению двух форм мейоспор: богатых запасными веществами мегаспор (= макроспор), которые возникают в мегаспорангиях (= макроспорангиях) и прорастают в относительно крупные женские заростки, и микроспор, которые образуются в микроспорангиях и дают более мелкие мужские заростки (см. рис. 11.137). По этому признаку следует различать равноспоровые (изоспорические) и разноспоровые (гетероспорические) группы — различие, которое неоднократно независимо возникало в разных эволюционных линиях папоротникообразных.
2 Точнее, раздельнополыми. — Примеч. ред.
1. Класс: псилофиты (Рsilophytopsida)
Вымершие к настоящему времени Рsilophytopsida — исходная группа папоротникообразных. Их вегетативное тело было построено из теломов, которые у представителей примитивных семейств были голые, а у продвинутых покрыты выростами. Спорангии располагались на концах дихотомически разветвленных осей или сбоку на главной или боковых ветвях. Все роды были изоспорическими. Настоящих корней еще не было; вместо них имелись ризомоиды, покрытые ризоидами. В осях были развиты только простые прото- или актиностелы.
Псилофиты были древнейшими наземными растениями, имевшими проводящие пучки и устьица. Они появились на рубеже силура—девона (т. е. примерно 400 млн лет назад), быстро достигли значительного разнообразия форм и вымерли уже к началу верхнего девона1.
1 Отдельные представители существовали еще в конце верхнего девона. — Примеч. ред.
1. Порядок: Rhyniales. Риниевые (морфологически наиболее примитивные представители) имели вегетативное тело в виде голых вильчатых теломов, часто еще с терминальными спорангиями.
К роду Соoksonia, встречавшемуся с верхнего силура до нижнего девона, относят древнейших из найденных к настоящему времени наземных растений. Их дихотомически разветвленные теломы высотой около 10 см с проходившей внутри протостелой несли спорангии, ширина которых превосходила длину.
Rhynia (включая Aglaophyton1 ранее относимый к Rhynia) — типовой род семейства Rhyniaсеае, представленный в нижнем девоне Шотландии двумя видами (см. рис. 11.130, А, В) — достигал 1 — 2 м в высоту2. Спорофит, по крайней мере, у одного вида (В) отрастал от ползучих осей3 («корневищ») — горизонтальных до дуговидно изогнутых, местами покрытых ризоидами без поперечных перегородок. Вертикальные, округлые в сечении, вильчато разветвленные теломы были безлистными.
1 Aglaophyton настолько своеобразен, что не может быть включен не только в Rhynia, но и в порядок Rhyniales. Возможно, что его вообще нельзя включать в Tracheophyta. — Примеч. ред.
2 Это сильно завышенная оценка. — Примеч. ред.
3 Эти оси были частью спорофита. — Примеч. ред.
Оси имели кутикулу и устьица еще весьма простого строения (см 3.2.2.1) и, очевидно, были органами ассимиляции. Таким образом, виды Rhynia были надземными растениями и образовывали заросли типа тростниковых. Проводящий пучок состоял из гидроидов у Aglaophyton (Rhynia) major и Homeophyton или из трахеид с очень простыми кольчатыми и. спиральными утолщениями стенок у Rhynia gwynne-vaughanii. Протостела (см. бокс 4.3, рис. А; рис. 11.130, С) у некоторых видов уже содержала метаксилему; типичных ситовидных клеток с ситовидными полями во внешней ткани пучка — флоэме еше не было1. Не было и вторичного утолщения. Относительно крупные спорангии, имевшие форму от цилиндрической до булавовидной, находились на концах или по бокам осей2. Они имели стенку из нескольких слоев клеток и вскрывались продольной щелью. В них формировалось множество тетрад изоспор (рис. 11.130, D, Е).
1 Флоэма не сохраняется в ископаемом состоянии, поэтому неправомерно отрицать ее наличие у Rhyniaceae. — Примеч. ред.
2 У Rhyniaceae спорангии всегда были терминальными. — Примеч. ред.
У Rhynia gwynne-vaughanii вследствие перевершинивания были боковые ответвления; перевершиниванию отчасти подвергались и спорангии (см. рис. 11.130, А).
У внешне сходного с родом Rhynia рода Ногneophyton1 продолговатые спорангии, располагавшиеся вместе плотными группами по 2 — 4, напоминают по своему строению спорогонии Sphagnum: спорогенная ткань, как колокол, облегала колонку, состоявшую из продольно вытянутых клеток. Спорангии вскрывались верхушечной порой.
1 Homeophyton существенно отличается как от Rhynia, так и от Sphagnum. — Примеч. ред.
Из-за отсутствия трахеид в последнее время Aglaophyton (Rhynia) major и Horneophyton больше не рассматривают как сосудистые растения.
2. Порядок: Zosterophyllales (-сеае)1. Распространенные в нижнем девоне по всему земному шару виды этого порядка также имели вегетативное тело в виде голых вильчатых ветвей, но их латеральные спорангии, вскрывавшиеся пре- формированной поперечной щелью, были чаще всего сгруппированы в колосовидные собрания (см. рис. 11.130, Н). Их рассматривают как предковую группу плаунов, с которыми часто и объединяют.
1 Zosterophyllales вместе с плауновидными, по-видимому, представляют эволюционную линию, параллельную всем остальным сосудистым растениям, и нет оснований включать его в Psilophytopsida. — Примеч. ред.
Удалось показать, что гаметофит некоторых родов (например, Zosterophyllum) представлял собой маленькое, звездчато разветвленное растение (см. рис. 11.130, F), которое на дуговидно изгибающихся вверх ножках образовывало зонтиковидные гаметангиофоры с архегониями в центре и антеридиями на периферии. Возникавший в результате оплодотворения молодой спорофит отделялся от гаметангиофора и развивался затем в самостоятельное растение (см. рис. 11.130, G, H). Соответственно неправильно рассматривать «корневища» Rhyniaceae как гаметофиты, которые на протяжении всей жизни связаны со спорофитом подобно гаметофитам мхов.
Таким образом, эти древние папоротникообразные имели гетероморфную смену поколений, однако эти поколения были развиты еще примерно одинаково. Отсюда эволюция могла идти как к мхам (преобладание гаметофита), так и к более продвинутым папоротникообразным (преобладание спорофита)2.
2 Это явный анахронизм. Мохообразные, по современным данным, к водорослям ближе, чем сосудистые растения. Поэтому их следует считать либо сестринской группой всех сосудистых растений, либо даже предками сосудистых растений. — Примеч. ред.
Как и у некоторых водорослей (например, Halycystis-Derbesia из Chlorophyta), разные поколения зостерофилловых также сначала описывали под разными родовыми названиями, пока только в последнее время не удалось установить их принадлежность к одному и тому же жизненному циклу. Так, название Taeniocrada соответствует стерильному, a Sciadophyton — фертильному гаметофиту, а спорофитное поколение носит название Zosterophyllum (см. рис. 11.130, F). Lyonophyton с ползучими, круглыми в сечении осями и проводящим пучком, содержащим гидроиды. а также с вертикальными, блюдцевидно расширенными вверху гаметангиофорами предположительно представлял собой гаметофитное поколение Aglaophyton major (см. рис. 11.130, В). У него даже удалось выявить структуру антеридиев со сперматогенными клетками.
Рис. 11.131. Psilophytopsida: А — Psilophyton, спороносная ветвь (3/4х); В — Lycopodiopsida, Asteroxylon mackiei, поперечный шлиф через актиностелу: темным показана ксилема, светлым флоэма (10х); С — то же, реконструкция (1/Зх)

3. Порядок: Trimerophytales (-сеае). Виды этого порядка, более высокоорганизованны по сравнению с представителями предыдущего порядка, встречались от нижнего до среднего девона. На распростертых (симподиально-ветвящихся) основных осях здесь образовывались дихотомически или тройчатоветвяшиеся боковые оси с расположенными на их концах спорангиями, уже собранными в группы. Этот порядок рассматривают как предковую группу «Progymnospermae» (см. рис. 11.288).
Dawsonites в противоположность Trimerophyton умел обращенные назад спорангии на искривленных осях. Psilophyton ornatum нес на осях мелкие выросты (рис. 11.131, А) и представляет собой, таким образом, переход к Lycopodiopsida1.
1 Как структурные особенности, так и время существования Psilophyton исключают его из числа предков Lycopodiopsida. Авторы изложили давно устаревшие представления о происхождении плауновидных. — Примеч. ред.
Psilophytopsida, будучи первыми наземными растениями, представляют исходную группу для других, филогенетически более продвинутых папоротникообразных1, а возможно, даже и определенных голосеменных.
1 См. предыдущее примечание. — Примеч. ред.
2. Класс: плауны (Lycopodiopsida)
Спорофит плауновидных, часто вильчато разветвленный, имеет простые, нерасчлененные, мелкие листья (= микрофиллия), обычно расположенные по спирали. Спорангии находятся, кроме немногих ископаемых форм, поодиночке адаксиально или у основания листьев (спорофиллов); чаще всего они располагаются в верхушечных стробилах. Следуя теломной теории, можно полагать, что расположение спорангиев на листьях возникло согласно схеме, приведенной на рис. 11. 133. Наряду с изоспорией широко распространена гетероспория. Сперматозоиды у немногих представителей многожгутиковые (Isoetes), но чаше всего двухжгутиковые — этим плауновидные отличаются от всех остальных папоротникообразных. Предполагают, что Lycopodiopsida происходят от просто устроенных псилофитов, имевших боковые выросты на осях и верхушечные стробилы спорофиллов (Zosterophyllales)2.
2 У Zosterophyllales не было спорофиллов. — Примеч. ред.
1. Порядок: Аsteroxylales (-сеае). Ветви были покрыты выростами в виде игл или шипов, рыхло либо более или менее плотно расположенных, они придавали растению сходство с современными плаунами.
В стеблях Asteroxylon mackiei, встречающегося совместно с Rhynia в нижнем девоне Шотландии, проходила стела, имевшая на срезе звездчатую форму (актиностела — см. бокс 4.3, рис. А; рис. 11.131, В). Лучи звезды представляли собой ответвляющиеся боковые тяжи, шедшие к месту прикрепления боковых выростов, в которых, однако, проводящего пучка не было. Ксилема стелы состояла из кольчатых и спиральных трахеид. Спорангии сидели непосредственно на стебле или в пазухах его выростов.
2. Порядок: Ргоtolepidodendrales. Вымершие к настоящему времени представители этого порядка развивали спорангии уже на листьях или в непосредственной близости от них. Несмотря на рыхлое расположение листьев, по облику они напоминают современные плауны (Lycopodiales). Представители обоих важнейших семейств — Dre- panophycaceae1 и Protolepidodendraceae — найдены в нижне- и среднедевонских отложениях. Родственные связи Drepanophycus (рис. 11.132, А) неясны (связь с Zosterophyllaceae); спорангии рас - полагались не на листьях, а между ними на коротких ножках, снабженных проводящим пучком. Листья Protolepidodendron (В) были на конце еще вильчато разделенными; спорангии находились на верхней стороне листовидных образований (спорофиллов; отчасти также вильчато разветвленных боковых осей).
1 Drepanophyусасeае, по-видимому, занимают промежуточное положение между Аsteroxylales и Lycopodiales, и нет оснований включать их в Protolepidodendrales. — Примеч. ред.
Рис. 11.132. Lycopodiopsida, Protolepidodendrales: A — Drepanophycus spinaeformis, нижний девон (1/4x); B — Protolepidodendron scharyanum, средний девон (1/4x)
Рис. 11.133. Переход (A — D) от верхушечного расположения спорангиев у Psilophytopsida (А) к эпифильному их расположению у Lycopodiopsida (D)
3. Порядок: Lycopodiales. Представителей этого преимущественно современного порядка чаще объединяют в единственное семейство (Lycopodiaceae). Это травянистые вечнозеленые растения (400 видов; 9 из них встречаются в Германии), густо покрытые более или менее игловидными листьями. Вторичное утолщение стеблей, как и у выше приведенных порядков, отсутствует.
У плауна (Lycopodium) (см. рис. 11.134) вильчато ветвящийся побег из-за перевершинивания одного из разветвлений кажется моноподиальным2 (см. 4.2.5). Побеги длинные, ползучие2. От нижней стороны побегов отходят дихотомически разветвленные корни, имеющие несколько инициальных клеток в апексе. Листья, мелкие, шиловидные, по существу расположенные по спирали2, имеют неветвящуюся среднюю жилку, а в остальном сходны с листьями Asteroxylales.
2 Только у некоторых видов. — Примеч. ред.
Мезофилл L. clavalum однородный; лишь у немногих видов возможно различить палисадную и губчатую паренхиму. Эпидермальные клетки листа без хлоропластов. Как и всегда при дихотомии, ветвление не связано с листьями.
Проводящая система стебля представляет собой производную от актиностелы обильно расчлененную плектостелу (см. бокс 4.3, рис. А) с ситовидными клетками во флоэме, несущими ситовидные поля на продольных стенках, но еще не имеющими ситовидных пластинок. Плектостела окружена снаружи влагалищем из неодревесневших клеток, внешний слой которых содержит крахмал; далее следует одно- или двухслойная эндодерма с лигнином в тонких клеточных стенках. Как и у всех папоротникообразных, у плаунов эндодерма является самым внутренним слоем первичной коры. Внешняя кора состоит из сильно одревесневших склеренхимных клеток (рис. 11.134, L).
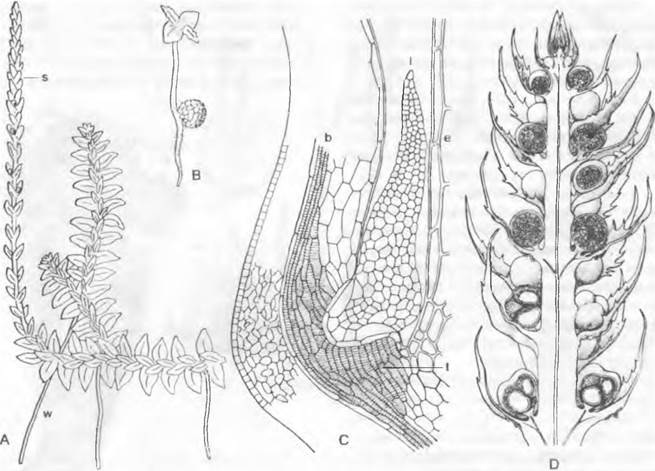
Рис. 11.134. Lycopodiopsida, Lycopodiales, Lycopodium clavatum:
A — антеридий, еще не вскрывшийся, продольный разрез (75х); В — сперматозоиды (400х); С — молодой, еще не вскрывшийся архегоний; D — готовый к оплодотворению, вскрывшийся архегоний (75х); Е — взрослый заросток (2х); F — заросток с молодым спорофитом (3/4х); G — спорофилл в колосовидных стробилах; Н — спорофилл с вскрывшимся спорангием (8х); J, К — споры в двух позициях (400х); L — поперечный срез стебля (100х); b — основание листа с полостью h; е — эпидермис; еп — эндодерма; h — полость; р — флоэма; г — первичная кора; s — крахмалоносное влагалище; х — ксилема
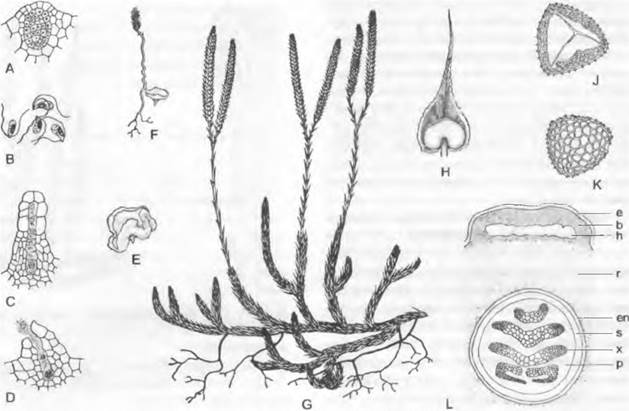
Часть ветвей обладает отрицательным геотропизмом. Их спорофиллы часто располагаются в плотных колосовидных стробилах над менее олиственным участком побега (рис. 11.134, G); при их образовании расходуется апикальная меристема, так что стробил завершает побег. Спорофиллы (Н) имеют вид широких чешуек и несут у основания на верхней стороне по одному крупному уплощенному почковидному спорангию, из которого высеиваются многочисленные мейоспоры — все одинакового размера (изоспоры) (1, К). С краев спорофиллов свисают вниз кожистые лопасти, каждая из которых как «индузий» прикрывает ближайший ниже расположенный спорангий.
Стенка спорангия состоит из нескольких слоев клеток, к которым изнутри примыкает секреторный тампетум. Спорангий вскрывается двумя створками, щель проходит по его вершине по линии, различимой по особому строению клеток. Споры остаются до созревания объединенными в тетрады; их многослойный экзоспорий покрыт сетью из выступающих ребрышек1(см. рис. 11.134, J, К). Споры прорастают в природе только через 6 — 7 лет и за счет находящегося в них запаса питательных веществ образуют сначала 5-клеточный предросток (рис. 11.135, А), который после периода покоя развивается дальше только тогда, когда в его нижние клетки внедрятся гифы гриба, образуя подобие микоризы (В: р).
1 Это не особый покров, а скульптура самого экзоспория. — Примеч. ред.
Заростки (см. рис. 11.134, Е, F; 11.135) живут в земле и представляют собой гетеротрофные беловатые клубневидные тельца величиной до 2 см с бугорчатыми лопастями и одноклеточными ризоидами, служащими для поглощения воды. В их питании, несомненно, большую роль играют микоризные грибы, живущие в клетках их периферических клеточных слоев (рис. 11.135, В, С). В естественных условиях половая зрелость наступает только через 12 — 15 лет, а общая продолжительность жизни заростков может составлять примерно 20 лет. Однако в искусственной чистой безбактериальной культуре все развитие занимает несколько месяцев. У некоторых видов заростки выступают верхней зеленеющей на свету частью над поверхностью земли. Заростки однодомные и несут многочисленные половые органы, преимущественно в верхней части (см. рис. 11.134, А — D; 11.135, С: а, аg). Антеридии (а) несколько погружены в ткань. Они многоклеточные, и каждая клетка, за исключением клеток стенки, дает начало одному овальному сперматозоиду, несущему у своей вершины только два жгутика (см. рис. 11.134, В). Архегоний(см. рис. 11.134, С. D; 11.135, С: аg), также погруженные, часто имеют многочисленные шейковые канальцевые клетки (до 20, однако может происходить их редукция вплоть до одной-единственной); самые верхние клетки стенки при вскрывании сбрасываются. Из оплодотворенной яйцеклетки после ряда клеточных делений возникает зародыш, подвесок (рис. 11.135, D, Е: е) которого проталкивает его в ткань заростка. Развитие гаустории, всасывающей из заростка питательные вещества, и первых листьев (bа), остающихся чешуевидными, показано на рис. 11.135, Е. Первый корень возникает как придаточный на побеге.
У Lycopodium спорофиллы собраны в стробилы, завершающие короткие боковые ветви; у Huperzia на вертикальных, вильчато разветвленных побегах в течение года последовательно образуются трофофиллы и спорофиллы. Diphasium имеет стробилы из спорофиллов, как у Lycopodium, но побеги у него плоские, дорсовент- ральные, с чешуевидными листьями.
Рис. 11.135. Lycopodiales:
А, В — Lycopodium, L. annotinum, развитие заростка: А — 5-клеточный бесцветный предросток с оболочкой споры (sh), ризоидальной клеткой (г), базальной клеткой (b), апикальной клеткой (s) (580х); В — молодой заросток, в нижних клетках которого обитает эндофитный гриб (р); апикальная клетка разделилась на три клетки апикальной меристемы (s1, s2, s3) (470х); С — Е — Diphasium, D. complanatum: С — зрелый заросток с антеридиями (а), архегониями (ад) и клетками, содержащими гифы гриба (показаны интенсивно темным) (24х); D, Е — развитие зародыша: D — зародыш, претерпевший первые деления клеток; базальная стенка 1 отделяет зачаток подвеска (е) от зачатка тела зародыша; перпендикулярные ей стенки 2 и 3 (последняя в плоскости среза), а также параллельная ей стенка 4 дают расположенные в два этажа группы из 4 клеток, из которых клетки, расположенные между стенками 1 и 4, образуют гаусторию, а самая нижняя — побег, Е — средняя стадия развития (112х); bа — верхушка первого листа; ss — верхушка побега
Виды Lycopodites из верхнего девона были уже очень сходны с современными представителями этого семейства. Таким образом, облик плауна сохранился неизменным на протяжении более 300 млн лет.
Если пока мы имели дело с изоспориой, то последующие порядки эволюционировали в сторону гетероспории. В пазухах листьев их представителей находится небольшой вырост —язычок(lígula) (рис. 11.136. С).
4. Порядок: Selaginellales. По облику травянистые Selaginella, или плаунки, несколько напоминают мхи1. Однако по строению своих генеративных органов и анатомическим признакам они однозначно являются папоротникообразными — соответственно настоящими сосудистыми растениями. В средних широтах они представлены несколькими видами, а в тропиках, напротив, обитает более 700 видов.
1 Отсюда их немецкое название Moosfarne (дословно «мхи-папоротники»). — Примеч. пер.
Облик. У видов Selaginella имеются отчасти лежачие, отчасти вертикальные, обильно вильчато разветвленные побеги; некоторые образуют дерновины, другие имеют лазающие побеги, поднимающиеся по кустарникам на несколько метров.
Стебель несет мелкие чешуевидные листья. Они расположены спирально или чаще всего в 4 ряда на дорсовентральных побегах: 2 ряда мелких так называемых спинных листьев и 2 ряда более крупных брюшных листьев (11.136, А; см. рис. 4.67, В; анизофиллия). Листья имеют только одну неразветвленную среднюю жилку и лишь изредка наряду с губчатой паренхимой обладают и палисадной паренхимой; у некоторых видов клетки мезофилла содержат только по одному крупному бокаловидному хлоропласту. Листья селагинелл несут на верхней стороне у основания возникающую из эпидермы мелкую кожистую бесхлорофилльную чешуйку — язычок (см. рис. 11.136, С), который служит органом поглощения воды: он очень быстро всасывает дождевую воду, стекающую по стеблю, и у некоторых видов связан трахеидами с проводящим пучком.
Рис. 11.136. Lycopodiopsida, Selaginellales, Selaginella: А — S. helvética, растение со стробилом (s) спорофиллов (2х); В — S. kraussiana, мегаспора с проростком растения (10х); С — S. lyallii, продольный срез через основание листа (b) (250х); D — S. se- laginoides, продольный разрез через стробил с мегаспорангиями (внизу) и микроспорангиями (вверху); у срезанных в медиальной плоскости спорофиллов над местом прикрепления спорангия виден язычок (I) (6х); е — эпидерма стебля; t — трахеиды; w — ризофор
В местах ветвления побегов у многих видов экзогенно возникают цилиндрические, вытянутые, растущие вниз, вильчато разветвленные бесцветные и безлистные оси — ризофоры(рис. 11.136, А: w), на свободных концах которых эндогенно возникают пучки корней. В определенных условиях ризофоры могут образовывать листья, как типичные побеги. Строение стелы варьирует от центральной протостелы и дистелы до сифоностелы. Вторичного утолщения нет. Крайне редко встречаются сосуды с лестничными утолщениями стенок. Эндодерма стебля (например, у S, kraussiana) состоит из трубковидных отделенных друг от друга клеток с поясками Каспари (трабекул).
Selaginellaceae (единственное семейство) отличается гетероспорией и очень сильноредуцированными заростками.
Расположенные на концах побегов стробилы спорофиллов (рис. 11.136, А, D) бывают простыми или разветвленными, четырехгранными с радиальной симметрией или дорсовентральными. Каждый спорофилл несет только один спорангии, отходящий из его пазухи1. Спорангии содержат крупные мегаспоры или мелкие микроспоры: они развиваются всегда в разных спорангиях — соответственно в мега- или микроспорангиях (рис. 11.137, А, В). Однако оба типа спорангиев встречаются в одном и том же стробиле (рис. 11.136, D). Таким образом, определение пола осуществляется уже в диплофазе модификационным путем (дипломодификационное определение пола). В мегаспорангиях все заложившиеся материнские клетки спор отмирают, кроме одной, которая после редукционного деления дает начало 4 крупным мегаспорам (♀) с бугристой поверхностью оболочки (рис. 11.137, А). В микроспорангиях возникают— также после редукционного деления — многочисленные мелкие микроспоры (♂) (рис. 11.137, В).
1 Спорангий располагается на самом спорофилле, а не в его пазухе. — Примеч. ред.
Стенка спорангия состоит из 3 слоев клеток (средний в зрелом спорангии очень узкий). Внутренний слой — тапетум (рис. 11.137, А: В питает споры, не разрушаясь при этом (секреторный тапетум). Спорангии раскрываются посредством когезионного механизма по заранее детерминированной линии, причем споры активно выбрасываются.
Гаметофит. Микроспоры начинают прорастать уже внутри спорангия. Спора при этом делится сначала на маленькую линзовидную клетку (рис. 11.137, С: р) и крупную клетку, которая последовательно делится на 8 стерильных покровных клеток и 2 или 4 центральные клетки (С). Эти клетки представляют собой мужской заросток, который так и остается внутри споры. Только маленькую линзовидную клетку можно рассматривать как вегетативную, утратившую свою функцию ризоидальную клетку; остальные клетки считают одним-единственным антеридием, из которых центральная клетка, окруженная покровными клетками (рис. 11.137, С — F: а), после дальнейших делений дает начало большому числу округляющихся сперматид (D — F: s). Затем покровные клетки вместе со своими стенками расплываются в слизистую массу, в которую погружена группа сперматид (F, G). Маленькая проталлиальная клетка (F: р), наоборот, сохраняется. Мужской заросток до этой стадии еще целиком находится в оболочке микроспоры; наконец, она разрывается, и ♂ гаметы, возникшие из сперматид, высвобождаются в виде слабоизогнутых, булавовидных сперматозоидов, несущих на переднем конце два длинных жгутика (Н).
Рис. 11.137. Lycopodiopsida, Selaginellales:
А, В — Selaginella inaequalitolia: А — мегаспорангий с одной-единственной тетрадой мегаспор и недоразвитыми материнскими клетками спор (70х), В — микроспорангий с тетрадами микроспор; С — G — S. stolonifera (640х); прорастание микроспоры, последовательные стадии; проталлиальную клетку (р) можно рассматривать как ризоидальную, С, D, F — сбоку, Е — со спинной стороны; на G протал- лиальная клетка не видна, клетки стенки, расплывшиеся; Н — S. cuspidate, сперматозоиды (780х); J, К — S. martensir. J — проросшая мегаспора, заросток с 3 ризоидальными буграми и несколькими архегониями с поверхности (112х); К— продольный разрез: 2 архегония с развивающимися зародышами, подвесок (е), гаустория (h) ризофор (w), зародышевые листья (к) с язычком (150х); L — схема развития Selaginella; светлые линии: гаплоидная фаза; темные линии или затушеванное черным — диплоидная фаза; 1 — мейоспоры; 2 — то же, после образования заростка; 3 — мегаспора и заросток с проросшим спорофитом; 4 — спорофит (S); 5 — спорангии; 6 — мейоспоры после высеивания из спорангиев; а — клетки стенки антеридия, s — сперматогенная клетка; t — тапетум; G — гаметофит; R! — редукционное деление

Нe столь сильноредуцированный женский заросток образуется в мегаспоре (J). Его развитие несколько неодинаковое у разных видов. Ядро споры делится на много дочерних ядер. Они располагаются в постенной плазме у вершины споры. Клеточные стенки сначала образуются здесь, а позднее возникают и ниже. Так в большинстве случаев вся спора сверху вниз последовательно заполняется крупными проталлиальными клетками. Одновременно в том же направлении начинаются дальнейшие деления этих клеток с образованием мелкоклеточной ткани. В верхней части заростка закладываются немногочисленные архегонии.
Стенка мегаспоры разрывается трехлучевой щелью (рис. 11.137, J); состоящий из мелких клеток бесцветный заросток немного выставляется наружу и образует на 3 бугорках своей ткани ризоиды, которые служат для поглощения воды. Затем происходит оплодотворение одного или нескольких архегониев1. Первой же перегородкой зигота делится на обращенный к шейке архегония подвесок (К: е) и собственно зародыш, который для освобождения из заростка должен изогнуться наружу (К); сначала он остается заключенным в женском заростке, который в свою очередь находится в мегаспоре.
1 Происходит оплодотворение не архегониев, а находящихся в них яйцеклеток. — Примеч. ред.
Большинство видов Selaginella живет на почве во влажных тропических лесах. Лишь немногие виды приспособились к сухим местообитаниям, как центральноамериканская S. lepidophylla, побеги которой, расположенные розеткой, при высыхании скручиваются (ложная «иерихонская роза»). Травянистые виды Selaginellites из карбона были уже разноспоровыми. Примерно 300 млн лет назад они выглядели как современные виды Selaginella.
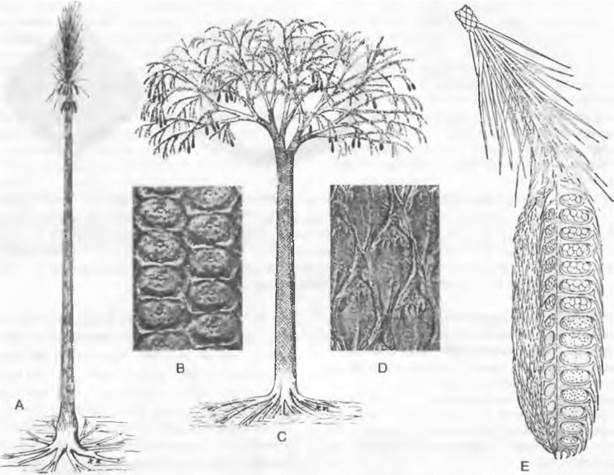
5. Порядок: Lepidodendrales (лепидофиты). Эти «древовидные плауны» (рис. 11.138) высотой до 40 м и диаметром ствола до 5 м достигли наибольшего расцвета в карбоне (см. рис. 11.163) и представляют существенный компонент каменного угля. Их линейные, спирально расположенные листья (типа микрофиллов, которые, однако, достигали до 1 м в длину) имели устьица в двух продольных желобках на нижней стороне. После их опадения на поверхности ствола оставались характерные рубцы и листовые подушки (рис. 11.138, В, D). Деревья прикреплялись к почве тянувшимися по ее поверхности (сырая почва!) повторно вильчато разветвленными ризофорами, обладавшими, как и ствол, вторичным утолщением (А, С); от них экзогенно отрастало множество относительно слабых корней своеобразного строения (так называемые аппендиксы), которые позже опадали, оставляя многочисленные рубцы на ризофоре, почему последние были названы стигмариями.
Рис. 11.138. Lycopodiopsida, Lepidodendrales: А, В — Sigillaria: А — реконструкция (1/80х), В — листовые подушки (2,5х); С — Е — Lepidodendron: С — реконструкция (1/200х), D — листовые подушки (натуральная величина), Е — стробил из спорофиллов (натуральная величина)
Листья имели простой, изредка вильчато разветвленный проводящий пучок, но в них еще не было палисадной ткани. Пятна, различимые попарно (рис. 11.138, В) или по две пары (D) на листовом рубце рядом с пятном от проводящего пучка, представляют собой места выхода тяжей рыхлой ткани, служивших для проведения воздуха и проходивших параллельно листовым следам в первичной коре. Стволы имели сифоностелу (рис. 11.139, А); тонкостенная флоэма была еще слабо дифференцирована. Кольцо малоактивного камбия образовывало в ходе вторичного утолщения новые ткани. При этом лестничные трахеиды (с несколько нетипичными утолщениями) вторичной древесины имели одинаковый диаметр. Вторичная древесина с уже имевшимися у ряда форм однорядными лучами напоминала древесину современных хвойных (но поры трахеид были не точечные, а лестничные, и, кроме того, как почти у всех растений из карбона Северного полушария, не было годовых колец). Однако вся вторичная древесина, очевидно, не играла заметной роли ни в обеспечении механической прочности деревьев, ни в проведении воды. В стволах также была меристема, соответствующая феллогену; она особенно интенсивно откладывала производные внутрь, так что образовывалась чрезвычайно мощная по сравнению с древесиной кора (у Lepidodendron до 99 % площади поперечного сечения (!), почему виды этого рода назвали «коровыми деревьями»; см. рис. 11.139, А). Эта кора состояла главным образом из механической ткани; кроме того, она посредством язычков, сохранявшихся длительное время после отпадения листьев, по-видимому, участвовала в поглощении воды.
Стволы Sigillariaceae (рис. 11.138, А) были покрыты продольными радами более или менее шестиугольных листовых подушек (В) (при вторичном росте в толщину они увеличивались вследствие дилатации). Их простые листья, имевшие длину до 10 м и ширину до 10 см, располагались пучком на конце неразветвленных или лишь слабо вильчато разветвленных стволов. В нижней части кроны на очень коротких боковых ветвях свисали крупные стробилы спорофиллов.
У Lepidodendгасеае (рис. 11.138, С) листья, располагавшиеся по спирали, достигавшие нескольких дециметров в длину, сидели на ромбических листовых подушках (D). Их стволы были обильно дихотомически разветвлены и несли на ветвях длиной до 34 м терминальные стробилы спорофиллов, внешне сходные с шишками хвойных (С, Е). Очень многочисленные, чешуевидные, черепицеобразно-расположенные спорофиллы закрывали спорангии, защищая их. Лепидодендроны были почти исключительно гетероспоровыми, некоторые из них имели в мегаспорангии только одну-единственную мегаспору до 6 мм и более; у некоторых представителей (Lepidostrobus major) мегаспора частично срасталась со стенкой спорангия, так что развитие заростка должно было происходить внутри спорангия. Заростки были сходны с заростками Selaginellaceae (рис. 11.139, В).
Рис. 11.139. Lycopodiopsida, Lepidodendrales: А — Lepidodendron, поперечное сечение ствола (схема), В — Bothmstrobus mundus, продольный шлиф мегаспоры с заростком (35х); С — Lepidocarpon lomaxi, продольный шлиф мегаспорангия (8х); ар — наружная перидерма; аг — наружная первичная кора; b — листовая подушка; h — обвертка (мегаспорофилл). ip— внутренняя перидерма; ir — внутренняя первичная кора; m — сердцевина: р — флоэма; pt — заросток; s — стенка споры; sp — стенка спорангия; х — ксилема
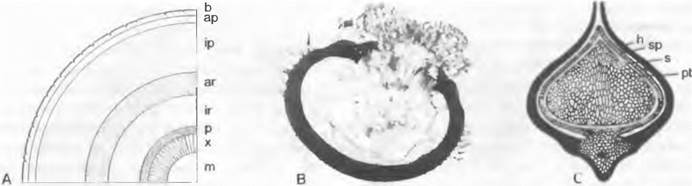
Большой интерес представляют некоторые каменноугольные формы (травянистые Miadesmia, Selaginellales, древовидные Lepidocarpon, Lepidodendrales) со структурами, напоминающими семя; их объединяют, несмотря на отсутствие близкого родства, под названием «Lepidospermae».
Мегаспорофилл располагался у этих «семенных» плаунов как обвертка вокруг спорангия (рис. 11.139, С: h); эта обвертка наверху была открытой и могла пропускать переносимые по воздуху микроспоры, обеспечивавшие оплодотворение (пока еще не известно, каким образом) заростка (pt), находившегося внутри единственной мегаспоры. Весь этот комплекс оставался на материнском растении и развивался здесь в семя1, в образовании покрова которого кроме стенки мегаспорангия участвовала также обвертка (мегаспорофилл). Мегаспорофиллы были сгруппированы в шишковидный стробил, сходный с таковым у современных голосеменных2.
1 Эта структура не гомологична семенам семенных растений. — Примеч. ред.
2 Шишка голосеменных только чисто внешне напоминает стробилы спорофиллов, ее организация принципиально иная. — Примеч. ред.
6. Порядок: Isoetales. Порядок в настоящее время представлен семейством Isoeta- сеае с двумя родами. Примерно 60 видов рода полушник (Isoetes) (рис. 11.140) представляют собой подводные или живущие на влажной почве многолетние травы с клубневидной, укороченной, изредка дихотомически разветвленной осью, которая может достигать очень большого возраста.
От 2 — 3 продольных борозд внизу оси отходят ряды дихотомически разветвленных корней, а от верхней части оси — собранные в розетку длинные шиловидные листья (у некоторых видов до 1 м длиной!).
Пронизанные 4 воздушными каналами листья имеют на верхней стороне своего расширенного основания продольное углубление (ямка, «fovea»). Большинство листьев представляет собой спорофиллы, несущие по одному спорангию в ямке; только самые внутренние листья розетки стерильны, но по форме они не отличаются от остальных. За ямкой имеется язычок в виде кожистой треугольной структуры с погруженным основанием (рис. 11.140, В, С).
Рис. 11.140. Lycopodiopsida, Isoetales:
А — С — Isoétes lacustris: А — целое растение (1/2х), В — базальный участок листа с язычком (li) и ямкой (f) (2х), С — его продольный разрез (4х); D — М — I. seíacea; развитие мужского заростка и образование сперматозоидов (500х); N — I. malinverniana, сперматозоид (1 100х); О — Q — женский заросток: О — Stylites andícola, заросток во вскрывшейся оболочке споры с архегониями (аг), правый с брюшной канальцевой клеткой (b) и яйцеклеткой (о) (60х); Р, Q — Isoetes echinospora, развитие архегония из одной поверхностной клетки (250х); е — экзина, внутри нее интина; f — ямка; h — клетки стенки шейки; hk — шейковая канальцевая клетка; m — микроспоры; р — проталлиальная клетка; s — сперматогенные клетки; t — трабекулы; w — клетки стенки; z — центральная клетка; дает брюшную канальцевую клетку (b)1
1 При делении она образует брюшную канальцевую клетку и яйцеклетку. — Примеч. ред.
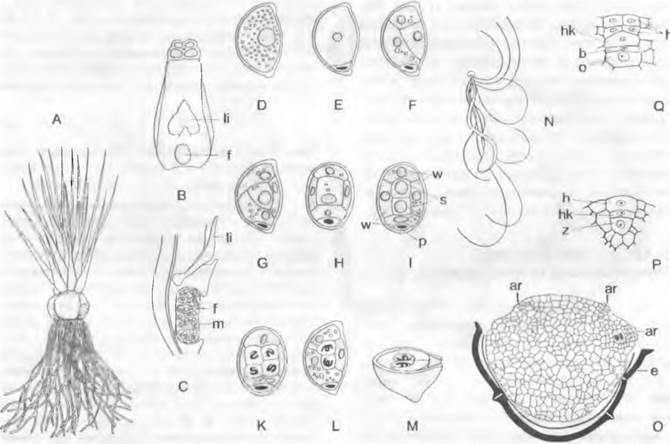
На внешних листьях розетки образуются мегаспорангии с многочисленными мегаспорами, на расположенных внутри более молодых листьях — микроспорангии с очень многочисленными микроспорами в каждом. Стенки спорангиев выстланы изнутри секреторным тапетумом. Заростки крайне редуцированы и образуются соответственно в микро- (♂) или мегаспорах (♀). Мужские заростки на ранних стадиях развития весьма сходны с находящимися еще в оболочке споры заростками Lycopodium (см. рис. 11.135, А). В остальном они подобны мужским заросткам Selaginella, однако образуют лишь 4 спирально закрученных сперматозоида, которые несут на переднем конце пучок жгутиков. Женский заросток (рис. 11.140, О; показан у близкого рода Srylites) по строению сходен с заростком Seloginella и заполняет всю мегаспору. Он развивает небольшое число архегониев в том месте, где разрывается оболочка споры. У зародыша, развивающегося в женском заростке, находящемся в мегаспоре, нет подвеска.
Эволюция. У Srylites (2 вида из Перу) — второго рода порядка, эволюционно, вероятно, более древнего, — стебли, покрытые листовыми рубцами, крупнее (15 см); имеется только одна продольная борозда с корнями; более выраженная тенденция к дихотомическому ветвлению. Особи вымерших представителей Isoetatales из семейств Рlеutometасеае и Nathorsianaceae были значительно крупнее, чем особи ныне живущих видов. Это в меньшей мере относится к нижнемеловой Nathorstiana и более явно — к Pleuromea из пестрого песчаника, неветвящиеся стволы которой толщиной примерно с человеческую руку (с короткими листьями и верхушечными гетероспоровыми стробилами) достигали высоты 2 м. Современные полушники находятся в конце эволюционного ряда, который берет начало от сигиллярий (Lepidodendrales), имевших относительно длинные листья и слабоветвившиеся или вообще неветвившиеся стволы, однако значительно более крупные, и идет дальше к Pleuromeia, Nathorstiana, а также современным видам Stylites, в котором происходило прогрессирующее укорочение стебля.
Из изложенного выше следует, что современные Lycopodiopsida часто можно связать с вымершими формами. Последние были представлены в карбоне многочисленными древовидными родами; они были значительно более развиты, чем в настоящее время (см. рис. 11.163), и в некоторых эволюционных линиях (Lepidospermae) достигли уровня организации семенных растений. Вследствие того, что климат в конце палеозоя становился все суше, древовидные плауны с их несовершенными структурами поглощения и транспорта воды вымерли или же были вытеснены появившимися формами с более совершенными проводящими системами (например, Соrdaitidae) (см. рис. 11.163). Травянистые плауны и селагинеллы, напротив, оказались столь хорошо приспособленными, что сохранялись неизменными на протяжении примерно 300 млн лет вплоть до настоящего времени («персистентные типы»). Правда, в современных ландшафтах они больше не играют никакой роли, тогда как древовидные плауны вместе с каламитами и некоторыми древовидными папоротниками доминировали в каменноугольных лесах, определяя их облик (см. 11.3.2.2; рис. 11.295).
3. Класс: хвощевидные (Еquisetopsida) (= Sphenopsida)
Этот класс отличается от предыдущего (II) по ряду признаков: спорофиллы четко отличаются от трофофиллов; у современных видов спорангии сидят по нескольку штук на зонтиковидных спорангиофорах, но никогда — в пазухах листьев; тапетум плазмодиальный (у Lycopodiopsida — секреторный тапетум); побег четко расчленен на узлы с мутовчато расположенными листьями и междоузлия.
Общие признаки хвощей: мелкие по сравнению со стеблем листья — так называемые микрофиллы, которые в противоположность остальным папоротникообразным расположены в мутовках. Стебель, чаще всего мутовчато разветвленный, четко дифференцирован на узлы и длинные междоузлия (см. рис. 11.143, А, В). Спорофиллы1, отличающиеся от ассимилирующих листьев, чаще всего имеют форму щитков с центральной ножкой, с нижней стороны которых свисает большое число спорангиев, и собраны в шишковидные верхушечные стробилы.
1 Листовая природа спорангиофоров хвощей не доказана. — Примеч. ред.
Полную противоположность морфологическому единообразию современных хвощей (рис. 11.141, Е, К) представляет богатство форм ископаемых Equisetopsida.
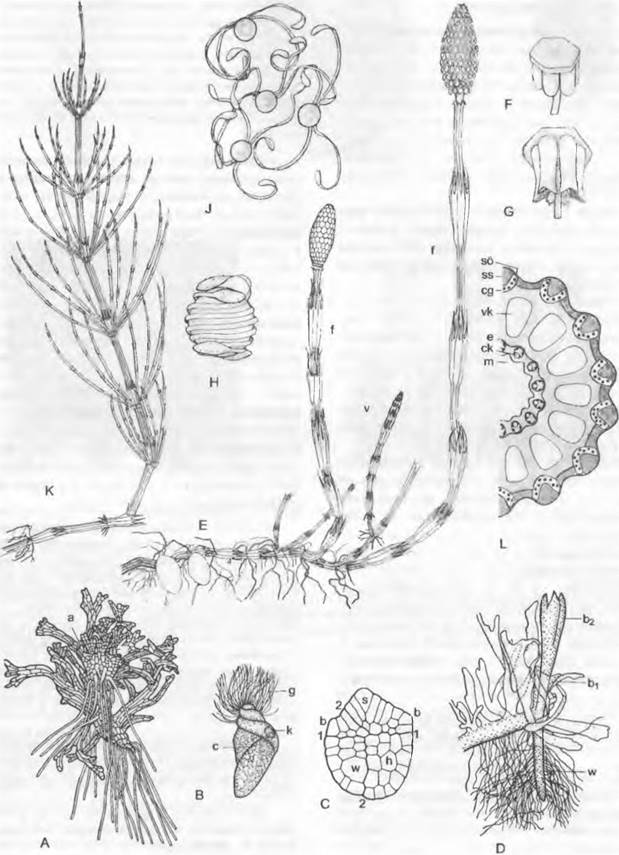
Рис. 11.141. Equisetopsida, Equisetaceae Equisetum:
A — ♀ заросток с архегониями с нижней стороны (17x); В — сперматозоид (1 250x); С — зародыш;1,2 — стенки квадрантов; из верхней половины, расположенной над базальной перегородкой 1, развиваются стебель1(s) и первая мутовка листьев (b), из нижней половины — корень (w) и гаустория (h) (165х); D — ♀ заросток с проростком (показан как более темный) сбоку, имеющим мутовки листьев и корень; Е — L — Equisetum arvense: Е — спороносные побеги (f), возникающие из почек подземного корневища, и начинающий расти бесплодный (вегетативный) побег (v) (1/2х); F и G — спорофиллы со спорангиями, на G вскрывшимися (6х); Н — спора с двумя спиральными лентами (гаптерами) периспория (360х); J— споры с развернувшимися в сухом состоянии гаптерами, при меньшем увеличении, чем на Н (100х); К— бесплодный, вегетативный побег (1/2х); L — поперечный срез стебля, с черной ксилемой и каринальными каналами (ск) в пучках, со склеренхимными тяжами (ss) в гребнях и ложбинках (16х); а — архегоний; b1, b2 — первые мутовки листьев; с — цитоплазма; cg — хлорофиллоносная ткань; е — эндодерма; g — жгутики; к — клеточное ядро; m — лизигенная сердцевинная полость; so — ряд устьиц; vk — валлекулярный канал
1 На самом деле апекс побега. — Примеч. ред.
1. Порядок: Sphenophyllales (клинолистные).
Представители этого порядка полностью вымерли. Их ископаемые остатки из палеозоя (от верхнего девона до перми) отличаются (чаше всего шестичленными) мутовками листьев, на конце вильчатых или клиновидно расширяющихся, с вильчато ветвящимися жилками (см. рис. 11.142, А). Это были травянистые растения, достигавшие длины примерно 1 м, возможно, лазающие. По облику они напоминали современные виды Galium. В тонких членистых, слаборазветвленных
стеблях проходил один триархный проводящий пучок; было вторичное утолщение (сетчатые трахеиды и трахеиды с окаймленными порами) (В). Четко выраженные стробилы были у некоторых видов равноспоровыми, а у некоторых— предположительно разноспоровыми1.
1 Равно- и разноспоровыми были не стробилы, а сами растения. — Примеч. ред.
2. Порядок: Equisetales (хвощeвые). Представители порядка составляют распространенную с конца девона и до настоящего времени основную группу класса и отличаются наличием в стебле сердцевинной полости, которая окружена венцом коллатеральных проводящих пучков2, к которому у древовидных палеозойских представителей снаружи примыкает вторичная древесина3.
2 Тяжи проводящих тканей в стеблях у Equisetales не тождественны коллатеральным проводящим пучкам. — Примеч. ред.
3 Вторичная древесина находилась снаружи от первичной ксилемы, но внутри от флоэмы, поэтому неверно утверждать, что она снаружи примыкала к кольцу пучков. — Примеч. ред.
Хвощи (Equisetасеае) в настоящее время представлены только одним родом — Equisetum, все виды которого (32) сходны между собой в основных чертах строения и развития.
Форма. От подземного ползучего корневища, находящегося в почве часто на значительной глубине, отрастают вертикальные надземные побеги с одной апикальной клеткой (см. 3.1.1.1; рис. 3.2, А, В), живущие чаще всего лишь один год. Они либо остаются простыми, либо ветвятся, образуя мутовки осей второго, третьего и т. д. порядков (см. рис. 11.141).
Ребристые оси состоят из удлиненных междоузлий. На узлах, отделенных друг от друга этими междоузлиями, расположены мутовки (см. 4.2.2) микрофиллов в форме заостренных зубцов с одним проводящим пучком каждый, которые срастаются основаниями во влагалище, охватывающее стебель (см. рис. 11.141, Е). Основания междоузлий, где они интеркалярно нарастают, одеты этими влагалищами. В узлах находится по одному кольцу внутренней ксилемы и флоэмы снаружи от нее (сифоностела). В междоузлиях проводящая система представлена отдельными тяжами, расположенными в паренхиме (эвcтела1; см. бокс 4.3, рис. А).
1 Это не эвстела, а артростела. так как ее отдельные тяжи не являются пучками листовых следов. — Примеч. ред.
В области узла находятся состоящие из протоксилемы тяжи листовых следов, расположенные друг за другом под зачатками боковых ветвей и над ними. Расположенные над зачатками тяжи выходят в листья только в области расположенного выше узла. Проводящие пучки вместе со своими тяжами листовых следов в смежных междоузлиях чередуются (рис. 11.142, Е) Боковые ветви прорывают влагалища между листьями.
Рис. 11.142. А, В — Sphenophytales, Sphenophyllum: А — S. cuneifolium, участок побега с вильчатыми и неразделенными листьями (1/Зх); В — S. plurífoliatum, поперечный шлиф стебля; внутри треугольная первичная ксилема с тремя протоксилемными группами, окруженная вторичной ксилемой (7х); С — Е — Eqisetales, Calamitaceae: С — Calamites са- rinatus,реконструкция (1/200х); D, Е — Arthropitys communis: D — поперечный шлиф через участок древесины (10х), Е — тангенциальный шлиф через молодой побег (10х); b — листовой след; с — ка- ринальный канал; i — внутриузловой канал; m — сердцевина; ms — сердцевинный луч; sx — вторичная ксилема; х — ксилема

При малой величине листовых пластинок, которые вскоре теряют хлорофилл, функция ассимиляции переходит на зеленые стебли. Коллатеральные (см. 3.2.4.3) проводящие пучкиочень бедны ксилемой. Самые старые части ксилемы вскоре исчезают, и на их местах образуются крупные полости, которые на поперечном срезе стебля выглядят как кольцо так называемых каринальных полостей (см. рис. 11.141, L). В расширенной сердцевине возникает большое воздухоносное межклеточное пространство (сердцевинная полость), а в первичной коре — кольцо из так называемых валлекулярных (ложбиночных) полостей (под ложбинками на поверхности побега). С внутренней стороны первичная кора чаще всего отграничена одно- или двуслойной эндодермой с поясками Каспари на антиклинальных стенках клеток.
Наружные стенки клеток эпидермы стебля у хвощей более или менее сильно пропитаны кремниевой кислотой (поэтому раньше их использовали для чистки металлической посуды). В бороздках между ребрышками расположены устьица, всегда по два в продольных рядах Им присущи следующие уникальные признаки: замыкающие клетки полностью покрыты околоустьичными клетками; при повышении тургора замыкающие клетки округляются, причем движение посредством утолщений смежных стенок передается на околоустьичные клетки, и устьица открываются.
Спорангии образуются на своеобразных спорангиофорах. Последние располагаются несколькими чередующимися мутовками, из-за сильно укороченных междоузлий собранными на концах побегов в шишковидные стробилы (см. рис. 11.141, Е). Сами спорангиофоры («спорофиллы») имеют форму столика на одной ножке, на нижней стороне которого располагается 5 — 10 мешковидных спорангиев (F, G); их снабжают концентрические проводящие пучки.
Спорогенная ткань в более молодом спорангии окружена многослойной стенкой. Ее самый внутренний слой (плазмодиальный тапетум) в результате растворения клеточных стенок образует периплазмодий, который внедряется между округляющимися спорами и расходуется на образование их оболочек1. Таким образом, ко времени созревания спор сохраняются только два внешних слоя клеток стенки спорангия; стенки клеток эпидермы имеют утолщения в форме колец и спиралей. Спорангии вскрываются продольной трещиной на внутренней стороне за счет сил сцепления между оболочками клеток стенки спорангия и водой, объем которой в этих клетках уменьшается вследствие испарения.
1 Плазмодиальный тапетум выделяет вещества, используемые при формировании оболочек спор, но некорректно утверждать, что он «расходуется» на образование этих оболочек. — Примеч. ред.
Вскрывшийся спорангий современных видов Equisetum высвобождает многочисленные зеленые споры со своеобразно устроенной оболочкой. На собственно споровой оболочке, состоящей из эндоспория и экзоспория, периплазмодий откладывает многослойный периспорий. Его самый внешний слой состоит из двух узких параллельных друг другу лент (гаптеры; см. рис. 11.141, Н, J), на концах, расширенных в виде лопатки. Во влажном состоянии они спирально закручены вокруг споры. При высыхании спор гаптеры развертываются, однако остаются соединенными серединами друг с другом и с эндоспорием (I). При этом они вытягиваются, но при увлажнении снова закручиваются (см. 8.4) и своими гигроскопическими движениями могут служить для распространения спор, сцепленных группами; соответственно гаметофиты часто растут тесными группами. Споры сохраняют всхожесть только несколько дней.
Все споры одинаковые. Они прорастают в сильно рассеченные на лопасти, зеленые заростки (см. рис. 11.141, А).
Заростки представляют собой сильно рассеченные дорсовентральные курчавые пластинки, которые могут быть однодомными или двудомными1. Определение пола потенциально двуполых заростков осуществляется фенотипически под влиянием внешних факторов. В условиях голодания возникают преимущественно ♂ гаметофиты. Развитие до половой зрелости длится всего лишь от 3 до 5 недель — очевидно, столь быстро для того, чтобы как можно скорее завершить гаметофитную фазу, чувствительную к наличию воды и низко конкурентоспособную по отношению к мхам. Мужские гаметофиты в противоположность женским сильно пигментированы каротиноидами — явление, которое известно также у мхов (стенки антеридиев) и грибов (♂ гаметы Allomyces) и может иметь значение как защита от мутагенного облучения.
1 Правильно — обоеполыми или однополыми. — Примеч. ред.
Антеридии погружены в ткань заростка, архегонии выступают над его поверхностью1. Спиралевидные сперматозоиды возникают в количестве примерно 250 — 1 000 на антеридий и имеют многочисленные жгутики (см. рис. 11.141, В).
1 Над поверхностью заростка выступают только шейки архегониев. — Примеч. ред.
Первая перегородка (базальная стенка, 1 — 1 на рис. 11.141. С) разделяет зиготу на две половины, из которых в противоположность Lycopodium (см. рис. 11.135, D) обе участвуют в образовании зародыша; подвесок не развивается. Первые листья появляются на побеговом полюсе сразу расположенные мутовкой и кольцевидно окружают апекс побега, который растет дальше посредством верхушечной клетки в виде трехгранной пирамиды (см. рис. 3.2, А). Зачаток первого корня находится сбоку от продольной оси зародыша (см. рис. 11.141. С: w); он растет вниз, прорывая ткань заростка (D).
Большинство видов распространенного от тропиков до холодных зон рода Equisetum предпочитает влажные местообитания. Южноамериканский Е. giganteum и некоторые другие тропические виды, будучи лазящими растениями, достигают до 12 м в длину, тогда как высота (Е. telmateia) видов, обитающих в Германии, может быть максимум 2 м.
У Е. arvense — хвоща полевого (см. рис. 11.141), как и у других видов с однолетними надземными частями, короткие боковые веточки подземных корневищ превращаются в округлые зимующие клубни, содержащие запасные вещества; однако имеются и вечнозеленые виды (например, Е. hyemale).
У некоторых видов хвощей одни надземные побеги стерильны и обильно ветвятся, другие же несут на вершине стробилы спорангиофоров, ветвятся позже и слабо или вообще не образуют боковых ветвей (см. рис. 11.141, Е, К).
Оба последующих семейства полностью вымершие. Встречавшиеся только в нижнем карбоне виды Archaeocalamitaceae имели вильчатые листья (рис. 11.143, А), которые на соседних узлах располагались друг над другом соответственно прохождению в узлах проводящих пучков.
Calamitaceae отличаются от Equisetaceae следующими признаками. На репродуктивных осях чередовались мутовки щитовидных спорангиофоров и ланцетовидных листьев («прицветников») (рис. 11.143, С). Наряду с равноспоровыми видами имелись и разноспоровые (D). Гаптер у спор не было. Виды широко распространенного в верхнем карбоне и перми рода Calamites (см. рис. 11.142, С) составляли важный компонент каменноугольных лесов и наряду с лепидодендронами и сигилляриями внесли существенный вклад в образование каменного угля. Экземпляры некоторых видов достигали высоты 30 м и благодаря мощному развитию вторичной древесины диаметр их стволов доходил до I м (С, D), однако, как и у Equisetum, имели большую сердцевинную полость («трубчатые деревья»). Стволы у большинства видов несли мутовки ветвей, но у некоторых не ветвились. Проводящие пучки (так же, как и у Equisetum) на верхнем конце междоузлия разветвлялись на три веточки. Две боковые веточки сливались каждая с боковой веточкой соседнего пучка, давая пучок следующего междоузлия, тогда как третья веточка отходила наружу как пучок листового
следа1 (Е). Радиально идущие «внутриузловые каналы», возникавшие в результате лизиса тонкостенных клеток, служили, вероятно, для аэрации. Листья (рис. 11.143. В) были простые, ланцетные, с одной жилкой; на конце листа находилась — как на листовых зубчиках ныне живущих хвощей, гуттирующих воду, — гидатода. Соответственно чередованию первичных пучков в следующих друг за другом междоузлиях листья располагались чередующимися мутовками.
1 На с. 250 эту часть проводящей системы правильно описывали как трубчатый участок артростелы («сифоностелу»). — Примеч. ред.
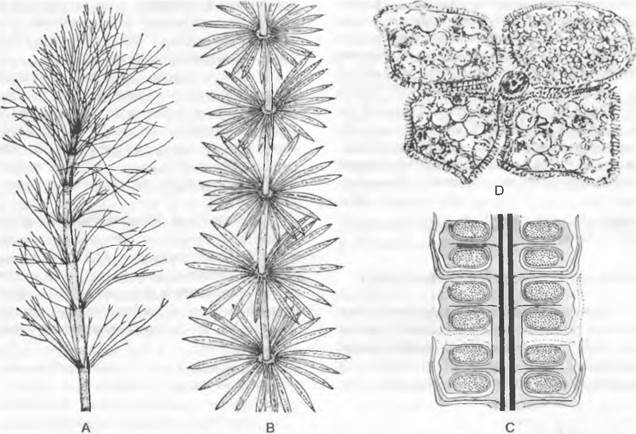
Рис. 11.143. Equisetales: А — Archaeocalamitaceae, Archaeocalamites radiatus (1 /Зх); В — D — Calamitaceae; В — Annularia stellata (1/2x), C — Calamostachys binneyana, продольный шлиф стробила со стерильными листьями (4х), D — Calamostachys casheana, тангенциальный шлиф через спорангиофор, несущий три мегаспорангия и один микроспорангий (22х)
Класс Еquisetopsida достиг наибольшего расцвета в палеозое и к настоящему времени весь, кроме рода Еquisetum, вымер (см. рис. 11.163). Этот род объединяет остатки более высокоразвитых форм, поскольку в мезозое среди представителей рода Equisetites были древовидные формы с вторичным утолщением. Таким образом, наши современные хвощи — всего лишь реликты, которые мы, однако, не можем считать потомками разноспоровых представителей из палеозоя, так как гетероспория всегда должна быть производной от изоспории, но не наоборот. Стало быть, современные хвощи должны были произойти от более ранних, еще равноспоровых форм. Некоторые из вымерших форм (Calamites, Sphenophyllum) были разноспоровыми; уровня образования семени, как у Lepidospermae, достиг род Calamocarpon.
4. Класс: псилотовые (Psilotopsida)
Сохранившиеся до наших дней виды Psilotum показывают определенное внешнее сходство с вымершими псилофитами (вильчатое ветвление побегов, отсутствие корней). Однако латеральные спорангии, срастающиеся в синангии, а также настоящие листья (микрофиллы) указывают на дальность псилотовых от Psilophytopsida и большую эволюционную продвинутость.
С другой стороны, помимо отсутствия корней наличие гаметофита, еще имеющего проводящие ткани, можно рассматривать как примитивный признак. Однако отсутствие корней интерпретируют и как вторичное упрощение (ср. лишенный корней водный папоротник Salvinia, утрату корней у эпифитов)1.
1 Отсутствие корней у псилотовых связано с их микотрофностью. — Примеч. ред.
Единственный порядок Psilotales включает низкие многолетние дихотомически разветвленные травы (рис. 11.144, А) с вильчатыми листьями (С). Надземные стебли имеют актиностелу или уже намечающуюся сифоностелу с одревесневшей сердцевиной (В). Безлистные протостелические корневища лишены корней (даже зародыш без зачатка корня), но несут нитевидные ризоиды и содержат микоризные грибы. Листья представляют собой чешуйки1 (микрофиллы, у Psilotum очень мелкие и без жилок), расположенные по рыхлой спирали. Спорангии имеют многослойную стенку, сращены по три2 в синангии (С, D) и еще не имеют настоящего тапегума (изоспоры получают питание от стерильных клеток археспория, которые не только окружают спорофиты, но и перемежаются с ними). Синангий сидит на очень короткой ножке в пазухе3 пары чешуек с вильчато разделенной верхушкой.
1 Вся надземная структура, включая чешуевидные образования и несущие их оси, может быть уподоблена вайе папоротников. — Примеч. ред.
2 У Tmesipteris— по две. — Примеч. ред.
3 Синангий располагается на алаксиальной лопасти перышка листа. — Примеч. ред.
Рис. 11.144. Psilotopsida, Psilotaceae: А — Е — Psilotum, Р. triquetrum: А — внешний вид (12х), В — поперечный срез стебля с актиностелой (точнее сифоностелой) (40х), С — участок побега с синангием в пазухе вильчатого листа (2,5х), D — поперечный срез синангия (8х), Е — споры (250х); F, G — Tmesipteris, Т. tannensis-. F — внешний вид (12х), G — спорофилл (2,5х); Н — К — Psilotum, Р. triquetrum: Н — заросток (15х), J — поперечный срез заростка (40х), К — сперматозоид (990х); аn — антеридии; аг — архегонии; с — внешний зеленый слой коры; ер — эпидерма; m — внутренняя кора; mу — микоризные клетки; р — флоэма; х — ксилема
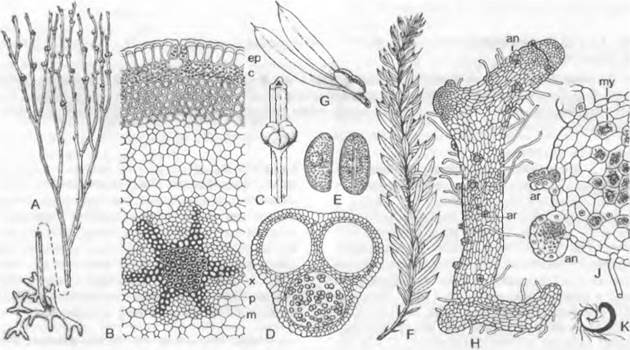
Гаметофиты, или заростки, достигают нескольких сантиметров в длину, радиально-симметричные, разветвленные (рис. 11.144, Н). Они бесцветны и живут под землей за счет микоризных грибов (J: mу). На поверхности они несут многоклеточные антеридии, в которых развиваются многочисленные сперматозоиды с множественными жгутиками. Мелкие архегонии (только с одной, редко с двумя шейковыми канальцевыми клетками) несколько погружены в ткань гаметофита. Особенно мощные заростки имеют проводящий пучок с одревесневшими кольчатыми трахеидами и эндодермой.
К Psilotopsida относят только два рода: Psilotum и Tmesipteris (с двумя видами в каждом, обитающими в тропиках преимущественно как эпифиты). У Tmesipteris вильчатые листья несколько крупнее. Они крыловидно сбегают по стеблю, а их плоскости параллельны стеблю. Однако их еще нельзя без оговорки приравнивать к листьям высших растений4. Иногда вильчатые надземные части трактуют как оси вай (листьев) с сидящими на них перистыми листочками, которые у Psilotum редуцированы сильнее, чем у Tmesipteris. Вильчатые разветвления лишь изредка соответствуют настоящей дихотомии (продольное деление трехгранной пирамидальной верхушечной клетки), чаще всего они происходят потому, что клетка, соседняя со старой верхушечной клеткой, становится второй верхушечной. Ризоиды гаметофита и спорофита развивают выводковые почки, с помощью которых осуществляется вегетативное размножение.
4 Они соответствуют перышкам на рахисе вайи папоротников. — Примеч. ред.
В ископаемом состоянии Psilotales не найдены. Однако их все-таки рассматривают как древние реликты, которые в то же время показывают четкие связи с рассматриваемыми далее Pteridopsida (через Gleicheniaceae с новокаледонским родом Stromatopteris и через Schizaeaceae).
5. Класс: папоротники (Рteridopsida) (= Filicopsida)
Представители этого класса папоротникообразных, собственно папоротники, отличаются крупными, часто сложно расчлененными мегафиллами, которые называют также «вайями».Спорангии, исходно располагавшиеся терминально, у более продвинутых форм находятся по краю листа или на его нижней стороне. Побеги, корни и листья растут — как и в ранее рассмотренном классе Equisetopsida — чаще всего благодаря деятельности единственной апикальной клетки, а не группы апикальных клеток, как у представителей Lycopodiopsida.
Вайи — чаще всего имеющие черешки и многочисленные жилки — в молодости обычно закручены на верхушке (исключение — Ophioglossales). Закручивание возникает из-за более быстрого роста абаксиальной (нижней) стороны молодых зачатков (соответственно листьев или теломов), и только потом происходит распрямление органа. Листья, развивающиеся таким акропластическим образом (см. 4 3.1.2), несут на нижней стороне многочисленные спорангии, часто собранные в группы (сорусы). Стебель чаще всего неветвящийся или слабо разветвленный. Возникновение листовых пластинок из систем теломов и переход спорангиев на нижнюю сторону листа вследствие более сильного роста его верхней стороны можно представить себе по схеме, приведенной на рис. 11.145, А — Н. Ископаемая переходная форма изображена на рис. 11.145, J. Аналогично можно представлять себе и возникновение крупных перистых вай. Представители вымерших Primofilices и ныне живущих групп папоротников Eusporangiatae, Leptosporangiatae и Hydropterides хотя и проявляют определенные родственные связи между собой, но являются не естественными таксонами, а ступенями развития, т. е. отражают ход развития разливных признаков.
Рис. 11.145. Pteridopsida: А — D — переход от фертильного телома к спорофиллу; Е — Н — перемещение спорангиев на нижнюю сторону листа; J — спорофилл Acrangiophyllum(напоминающего папоротник растения неясного систематического положения) из верхнего карбона (7х); К — М — жилкование перышек листьев растений, сходных с папоротниками; К — вееровидное жилкование (Archaeopteris, верхний девон), L — перистое жилкование (Alethopteris, представитель семенных папоротников из верхнего карбона), М — сетчатое жилкование (Linopteris верхний карбон) (12х)
Ступень развития; Первопапоротники1 (Primofilices) (= Protopteridiidae)
1 Значительная часть этой группы в принятом здесь объеме принадлежит к Progymnospermae и не имеет отношения к папоротникам. — Примеч. ред.
В качестве предков Рteridopsida рассматривают Psilophytopsida. Связующее звено между ними представляют Рrimofilices, которые, с одной стороны, еще сходны с псилофитами, а с другой стороны, имеют признаки более высокого развития. Общим для всех Рrimofilices было наличие терминальных спорангиев (рис. 11.146, В), а также то, что элементы перистых листьев располагались не в одной плоскости («объемные вайи»). Переход от псилофитов к Primofilices происходил столь постепенно, что относительно некоторых форм (Protopteridium и Рseudosporochnus, рис. 11.146, А — Е) может возникнуть сомнение, причислять ли их еще к псилофитам или уже к папоротникам1. Филогенетическому положению Primofilices соответствует и их распространение во времени: они появились в среднем девоне и вымерли в нижней перми.
1 Рrotopteridium можно причислять к Progymnospermae, но никак не к папоротникам. — Примеч. ред.
В пределах группы Рrimofilices происходит переход от расположенных почти пучками вильчатых ветвей с расположенными на их концах спорангиями (например, Рseudosporochnus; рис. 11.146, А, В) к клиновидно-уплощенным, неправильно вильчато разветвленным листьям и сходным с ними спорофиллам с расположенными по их краям спорангиями (например, Сladoxylon: рис. 11.146, М)2. Спорофиллы у некоторых видов располагались вперемежку со стерильными листьями, а у некоторых видов — группами, хотя еще и не были объединены в «цветки»3. Прогрессировавшее срастание теломов боками привело к появлению широких плоских листьев с вильчатым жилкованием, какие встречались уже в верхнем девоне (см. рис. 11.145, J; ср. также Adiantum из современных Leptosporangiatae — см. рис. 4.60, А).
2 Признать спорангии Сladoxylon расположенными по краям спорофиллов было бы слишком поспешным. — Примеч. ред.
3 Цветков нет и у всех остальных папоротников; они присущи только покрытосеменным. — Примеч. ред.
Позднее из вильчатого жилкования у папоротников и семенных папоротников постепенно развилось сетчатое жилкование. В верхнем девоне встречалось вееровидное жилкование с вильчатым ветвлением жилок, в нижнем карбоне впервые появилось перистое жилкование, а в верхнем карбоне — сетчатое, которое наиболее совершенным образом обеспечивало лист водой и питательными веществами (см. рис. 11.145, К — М).
Предпосылкой для появления таких плоских листовидных образований было то, что теломы расположились в одной плоскости. Однако у примитивных форм они еще отчасти располагались перпендикулярно друг к другу (как это имеет место и ныне, например, у Ophioglossales), даже уплощение могло еще отсутствовать, так что «листья» были круглыми в сечении. И то и другое имело место у Stauropteris (рис. 11.146, G). верхний карбон; в таких цилиндрических листьях-теломах уже могла быть палисадная паренхима.
Primofilices были преимушественно равноспоровыми; отдельные виды были уже разноспоровыми (Aichaeopteridales)1. Они имели спорангии с многослойной стенкой, т. е. были эвспорангиатными. У отдельных представителей уже встречались особые механизмы вскрывания спорангиев. Увеличивалось также разнообразие стел — от протостелы до эвстелы.
1 Аrchaeopteridales — это не папоротники, а праголосеменные. Рrogymnospermae. — Примеч. ред.
Возникновение признаков, характерных для современных Pteridopsida, можно, таким образом, проследить на группе форм Primofilices, так что мегафильные Pteridopsida вместе с Equisetopsida можно вывести из Psilophytopsida как эволюционную ветвь, параллельную микрофильным Lycopodiopsida.
Систематика. Primofilices в целом составляют весьма гетерогенную группу, которую можно подразделить на 5 порядков.
1. Порядок; Pseudosporochnales2. Представители порядка встречаются в (нижнем) среднем девоне; например, Pseudosporochnus (рис. 11.146) едва достигал высоты 1 м, с неразветвленным стволом и множеством одинаково развитых, слабо вильчато разветвленных боковых осей, которые несли многочисленные тонкие дихотомирующие веточки. В отдельных случаях концы ветвей были несколько расширены — начало уплощения и слияния (в терминах теломной теории). Боковые ветви с их расширенными ассимилирующими пластинками можно рассматривать как структуры, предшествующие крупным, многократно перистораздельным листьям (мегафиллам), или вайям.
2 Вероятнее всего, этот порядок следует объединять с Сladoxylalех. — Примеч. ред
2. Порядок: Protopteridales1. Представители этого порядка по строению своих «вай» также еще более или менее напоминают псилофиты. У некоторых родов был мощный ствол с лестничными трахеидами2 во вторичной древесине. Роды Нуеnia и Сalamophyton, рассматриваемые ранее как примитивные Equisetopsida, также относятся, поданным современных исследований, к этому кругу форм, равно как и Protopteridium (рис. 11.146, С). Aneurophyton, Тetraxylopteris, Rhacophyton и Pertica (рис. 11.146, F) 3.
1 Это не папоротники, а праголосеменные. — Примеч. ред.
2 У настоящих Рrotopteridales были точечные трахеиды вторичной ксилемы. — Примеч. ред.
3 Protopteridium, Aneurophyton и Tetraxylopteris — это праголосеменные, Нуеnia и Саlamophytor — представители Сladoxylales (= Рseudosporochnales), Rhacophyoton принадлежит к примитивным Zygopteridales, а Рertica — к Trimerophytales. Их неправомерно относить к одному порядку. — Примеч. ред.
3. Порядок: Сladoxylales. Виды порядка (Сladoxylon. рис. 11.146, J) жили от среднего девона до нижнего карбона. Строение их стелы, которая состояла из многочисленных V-образных в поперечном сечении отдельных пучков (рис. 11.146, L), иное, чем у всех других сосудистых растений1.
1 К Сladoxylales, относят растения со стелой кладоксилеевого типа. Растения, у которых не удается достоверно выявить строение стелы, включают в порядок Рseudosporochnales. Сохранение обоих порядков оправдано исключительно практикой палеоботаников строить отдельные классификации для разных органов и для органов разной сохранности — в противоположность практике неонтологов классифицировать целые организмы. — Примеч. ред.
Рис. 11.146. Primofilices:
А — В — Pseudosporochnales: А — Pseudospo- rochnus, реконструкция, В — то же, конец ветви (натуральная величина); С — F — Protopteri- dales: С — Е — Protopteridium hostimense,девон, С — вайя ( 1 /4х), D — стерильные, Е — фертильные перышки (Зх), F — Pertica quadrifaria, девон, ветвь со спорангиями (s); G, H — Coeno- pteridales, Stauropteris oldhamia,карбон: G — участок стерильной вайи, реконструкция (натуральная величина), Н — то же, спорангий с участком стенки, по которому он вскрывался (35х); J — М — Cladoxylales, Cladoxylon scoparium, средний девон: J — участок ветви (2/Зх), К — листочки (2х), L — поперечный срез плектосте- лы (4х). М — группа спорангиев (2х)

4. Порядок: Coenopteridales. Виды порядка (от верхнего девона до нижней перми, расцвет в нижнем карбоне) полностью характеризуются разветвленными еще в трехмерном пространстве вайями (Stauropteris, рис. 11.146, G; Воtryopteris, лазящий папоротник Ankyropleris и многие другие1). В стенке спорангия некоторых видов уже можно распознать группу толстостенных клеток, обеспечивавших его вскрывание по заранее закладывавшейся линии разрыва, как в спорангии Osmunda (см. рис. И. 156, А).
1 У Аnkyropteris и Воtryopteris были настоящие перистые вайи, вполне сравнимые с вайями большинства современных папоротников. — Примеч. ред.
5. Порядок: Archaeopteridales2. К этому порядку (разноспоровому) относят верхнедевонский космополитный род Archaeopteris, представленный многими видами (см. рис. 11.288. С). Это были уже стройные деревья с дважды перисто- раздельными объемными вайями. Лопатковидные перышки имели перистое жилкование (см. рис. 11.145, J); по обе стороны от основания вай располагались два прилистника, как у Marattiales. Нижние перышки фертильных вай несли спорангии, расположенные по краю, обращенному к верхушке вайи. Микроспорангии содержали много микроспор диаметром 0,03 мм, а мегаспорангии — 8 — 16 мегаспор диаметром 0.3 мм (см. рис. 11.288, С: 8). В стволах, достигавших высоты до 9 м и толщины до 1,5 м, имелась мощная вторичная ксилема из трахеид с араукариоидной поровостью стенок (см. 11.2, Araucariaceae). Archaeopteris объединяет, таким образом, признаки папоротников3 и голосеменных.
2 Это праголосеменные растения, а не папоротники. — Примеч. ред.
3 Сходство его с папоротниками конвергентное. — Примеч. ред.
Некоторые авторы объединяют Protopteridales — промежуточное звено между Psilophytopsidr и Filicopsida, и Archaeopteridales, эволюционно находящийся между Psilophytopsida и Gymnospermae, в группу «Progymnospermae». По вторичному росту в толщину и ряду других признаков они представляют собой обширную переходную группу между птеридофитами и голосеменными1.
1 Progymnospermae представляет собой переходную группу между Trimerophytales и голосеменными. — Примеч. ред.
Ступень развития: эуспорангиатные папоротники (Eusporangiatae) (= Ophioglossidae)
Спорангии, имеющие многослойную стенку (рис. 11.147, F), развиваются каждый из нескольких клеток.
1. Порядок: Ophioglossales. Представители порядка (единственное семейство Ophioglossaceae, около 80 равноспоровых видов) имеют вайи, которые состоят из ассимилирующей (лишенной палисадной ткани) зеленой и перпендикулярно к ней расположенной фертильной желтоватой частей (рис. 11.147, A, D). Листья представляют собой трехмерные вайи и в этом отношении соответствуют исходному типу вай (ср. Primofilices). У спороносной части рост в ширину подавлен.
Продольный рост обеспечивает не одиночная крупная апикальная клетка, а несколько инициальных клеток, слагающих конус нарастания. На коротком подземном стебле ежегодно развивается чаще всего лишь одна-единственная вайя с длинным черешком и небольшим кожистым влагалищем. В молодости она не закручена. В нижних частях стебля находится протостела, которая выше разделяется на трубку проводящих пучков. Сильно редуцированные, имеющие в длину несколько миллиметров подземные заростки лишены хлорофилла и представляют собой многослойные клубеньки, часто многолетние, которые существуют благодаря симбиозу с микоризными грибами. Антеридии и архегонии погружены в ткань заростка (рис. 11.147, С). Зародыш, возникающий из оплодотворенной яйцеклетки, у некоторых видов в течение нескольких лет ведет подземный образ жизни.
У Воtrychium как ассимилирующая часть вайи с вильчато расходящимися жилками, так и ее спороносная часть перистые. Округлые спорангии располагаются по кра-
ям перьев и не срастаются между собой (рис. 11.147, D, Е). В подземном побеге происходит слабое вторичное утолщение (единственный случай среди всех современных папоротников).
У Ophioglossum зеленая часть листа имеет форму языка и сетчатое жилкование, а желтая спороносная часть — форму простого цилиндра. Спорангии погружены в ткань, располагаются в два ряда и срастаются между собой боками (рис. 11.147, А). Питанию растений, очевидно, способствуют микоризные грибы, всегда присутствующие в корнях. У Ophioglossum simplex лист большей частью вообще не имеет ассимиляционной ткани, а только несет спорангии. Заросток цилиндрический (рис. 11.147, С). Число хромосом необычайно большое (у О. vulgatum п = 256, у О. reticulatum п = 630).
Рис. 11.147. Eusporangiatae, Ophioglossales:
А — С — Ophioglossum, О. vulgatum: А — спорофит (1/2х), В — продольный срез верхушки фертильной части листа (2х), С — заросток с антеридиями (а), архегониями (аg), молодым спорофитом (s), имеющим первый корень, гифами гриба (h) в ткани заростка (10х); D — G — Botrychium, В. lunaria: D — спорофит (1/2х), Е — спорангии, вид снизу, F — продольный срез незрелого спорангия с многослойной стенкой; внутри материнские клетки спор, окруженные клетками тапетума (10х), G — срез заростка (35х); е — зародыш

2. Порядок: Marattiales. В ограниченный тропиками порядок включают также примитивные и древние папоротники. Их современные виды несут на коротком клубневидном стебле пучок вай, чаще всего имеющих несколько метров в длину, многократно перистораздельных, в молодости закрученных и у основания снабженных парными прилистниками.Жилкование открытое (ср. Ophioglossum с сетчатым жилкованием). Спорангии с одинаковыми спорами у некоторых родов срастаются боками в капсуловидные многокамерные, позже резко вскрывающиеся синангии (рис. 11.148, В, С), у других родов они свободные и сгруппированы в сорусы.
Рис. 11.148. Еusporangiatae, Магattiales: А — древовидный папоротник Меgaphyton (семейство Мarattiaceae), реконструкция (верхний карбон). На стволе в два ряда располагаются рубцы от отпавших вай. Основание ствола усилено мантией из отходящих от него корней, растущих вниз; В — Рtychocarpus unitus, верхний карбон, нижняя сторона перышек с синангиями (8х); С — то же, поперечный срез синангия (60х)
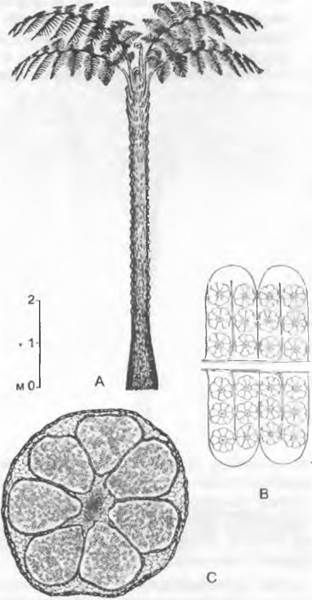
Долго живущие заростки содержат эндофитные микоризные грибы, однако обитают на поверхности почвы в виде автотрофных многослойных талломов. Внешне они напоминают печеночники. На нижней стороне у них развиваются погруженные антеридии и архегонии.
Современные Мarattiles насчитывают примерно 200 видов из нескольких родов. Они населяют тропические леса: например, Angiopteris встречается в Азии (длина вай до 5 м!), Danaea — в Южной Америке, Маrattia — по всему тропическому поясу.
Эволюция. Первые Маrattiales появились в карбоне. По-видимому, они произошли от изоспоровых Рrimofilices, которые почти все имели спорангии с многослойными стенками. До красного лежня их видовое разнообразие было больше, и они были более широко распространены, чем в настоящее время. Это были деревья со стволами, покрытыми корнями и достигавшими высоты 10м (самым мощным и широко распространенным растением была Asterotheca arborescens). Тогда они полностью господствовали над Leptosporangiatae. Особенно приметным был Megaphyton с вайями, расположенными не по спирали, а в два ряда (рис. 11.148, А).
Ступень развития: лептоспорангиатные папоротники (Leptosporangiatae) (= Pterididae)
Спорангий (см. рис. 11.128) развивается из одной эпидермальной клетки и имеет тонкую стенку, которая после раннего лизиса тапетума состоит из одного слоя клеток. Относимые сюда папоротники — преимущественно тенелюбивые растения. Многочисленные их виды (90 % всех Pteridopsida, около 9000 видов) распространены по всему земному шару; наиболее широко они представлены в тропиках, где весьма разнообразны: от карликов размером в несколько миллиметров (например, виды Didymoglossun из семейства Нmenophуllасеае) до пальмовидных деревьев высотой до 20 м (см. рис. 11.150). Ствол древовидных папоротников, чаще всего толщиной с человеческую руку (семейство Суаtheaceae, роды Суathеа, Dicksonia, Сibotium), не ветвится и несет на вершине розетку многократно перистораздельных вай длиной до 3 м. В Германии встречаются папоротники, наоборот, в виде преимущественно травянистых растений с подземным многолетним вертикальным или приподнимающимся, слабо разветвленным корневищем, которое у Рteridiumможет быть до 40 м длины и жить до 70 лет1.
1 Корневища ряда европейских видов, в том числе и Рteridium aqulinum, горизонтальные и ветвятся интенсивно. — Примеч. ред.
Стволы — у травянистых форм корневища — в молодости имеют чаще всего центральную протостелу, которая в более старых частях переходит в структуру типа сифоно- и полистелы самых разнообразных форм (см. бокс 4.2, рис. А) чаще всего с центральной ксилемой и периферической флоэмой (рис. 11.151, А; 11.149; см. также 3.2.4)2. Изредка даже образуются сосуды (как у Рteridium aquilinum, рис. 11.149). Проводящий пучок окружен эндодермой (рис. 11.149). Вторичного утолщения нет, а прочность стволов достигается иначе, чем у Lycopodiopsida и hquisetopsida: многочисленные пучки листовых следов проходят чаще всего на большом расстоянии в коре и вместе со склерен- химными пластинками (рис. 11.151, А) укрепляют стебель (см. также 3.2.3). У некоторых древовидных папоротников прочность ствола повышается также благодаря мантии из жестких придаточных корней. Иногда такой покров может быть чрезвычайно толстым (до нескольких дециметров!).
2 У папоротников бывает не полистела, а диктиостела, иногда полициклическая. Сифоностела и диктиостела находятся не в старых, а, наоборот, в более молодых участках ствола (корневища). — Примеч. ред.
Рис. 11.149. Leptosporangiatae, Рteridales, Pteridium aquilinum. Проводящий пучок, поперечный и продольный разрезы (100х); е — эндодерма; g — основная паренхима; s — ситовидные клетки; t — лестничные сосуды
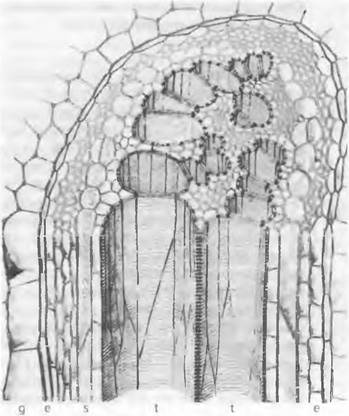
Мегафиллам, возникшим из многих теломов, свойственны разные варианты жилкования. Древние вильчатые листья (см. рис. 11.157) или вильчатые жилки (см. рис. 4.60, А; на листьях со слитными перышками) встречаются редко; как правило, они бывают только у листьев проростков и молодых растений. Полностью развитые листья весьма разнообразны. Часто вайи перисто-сложные (например, от дважды- до четыреждыперистых у Pteridium aquitinum, орляка; дваждыперистые у Dryopterisfilix-mas, щитовника мужского; однократноперистые у Polypodium vulgare, многоножки обыкновенной). Однако встречаются и простые листья с доминирующей центральной и менее мощными боковыми жилками (Phyllitis scolopendrium, листовник сколопендровый, рис. 11.151, С). Их очень долгий, иногда неограниченный верхушечный рост в противоположность росту листьев семенных растений обусловлен деятельностью двусторонней апикальной клетки, которая, однако, позже часто заменяется группой инициальных клеток.
Лист часто развивается в течение многих лет. У орляка (Pteridium aquilinum), например, на каждой короткой веточке корневища ежегодно закладывается только один лист, которому требуется 3 года, чтобы полностью развиться. После отмирания листья оставляют — прежде всего у древовидных папоротников, где они сохраняются несколько лет после того, как полностью развернутся, — крупные, бросающиеся в глаза рубцы (см. рис. 11.150). По гистологическому строению (наличию палисадной и губчатой паренхимы) эти листья в значительной мере напоминают листья высших наземных растений, однако клетки эпидермы у папоротников чаще всего содержат хлоропласта.
Рис. 11.150. Pteridopsida, Сyatheales, Cyathea crinita. Древовидный папоротник из Шри- Ланки (1/100х)

Спорангии образуются в большом числе по краю или чаше всего на нижней стороне листьев (см. бокс 4.1, рис. А; рис. 11.152, В — D). Спорофиллы по внешнему облику, как правило, лишь немного отличаются от стерильных листьев (трофофиллов); однако у представителей некоторых родов они имеют явно иной облик — прежде всего вследствие редукции пластинок перышек (ср. Matteuccia, Blechnum, Osmunda).
Рис. 11.151. Pteridopsida: А — Pteridium aquilinum, поперечный срез корневища (7х); В — Asplénium nidus, схема роста; С — Phyllitis scoiopendrium (1/4х); al — внешний проводящий пучок; е — эпидерма; il — внутренний проводящий пучок; р — паренхима; s — склеренхимные пластинки; sr — склеренхимное кольцо
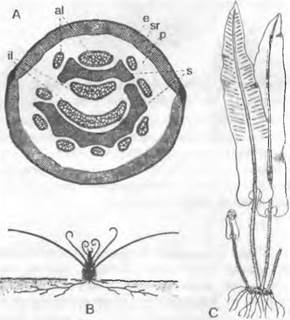
В типичном случае (например, у Aspidiales, к которым относится подавляющее большинство европейских папоротников) спорангии помногу объединяются в сорусы. Эти спорангии возникают на выступе ткани листа — плаценте (рис. 11.152, В; плаценту называют также рецептакулом) и у многих видов до созревания покрыты и защищены кожистым выростом поверхности листа — так называемым покрывальцем (или индузием; В — D). Отдельный спо рангий в зрелом состоянии представляет собой маленькую коробочку на ножке, которая содержит большое число мейоспор почти всегда одинакового размера (изоспория!)1. Очень характерно по-разному устроенное кольцо, которое у Рolypodiacaea в виде выступающего ряда клеток с сильно утолщенными радиальными и внутренними стенками проходит через спинную сторону и вершину спорангия до середины его брюшной стороны (см. рис. 11.156, D). Посредством механизма сцепления между стенками клеток кольца и находящейся в них водой (при участии клеток устья, см. 8.4) кольцо осуществляет вскрывание спорангия и выбрасывание спор (см. рис. 8.37).
1 У лептоспорангиатных папоротников чаще всего в спорангии развивается 64 споры. — Примеч. ред.
Рис. 11.152. Рteridopsida, Аspidiales, Driopteris: А — внешний вид (1/4х); В — разрез через сорус: показана плацента со спорангиями и зонтиковидным индузием (30х); С — перышко с молодыми, еще покрытыми индузием сорусами; D — то же, на более поздней стадии, со сморщившимися индузиями (Зх)
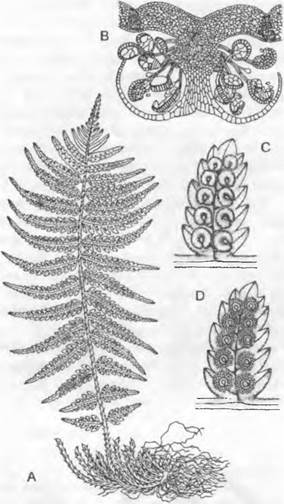
Из прорастающей споры развивается недолговечный гаплоидный заросток (см. рис. 11.128, А, В; 11.153). который не превышает нескольких сантиметров в длину и, как правило, образует как антеридии, так и архегонии. Определение пола осуществляется в норме модификационно. Заростки потенциально обоеполые. Только у представителей австралийского рода Platyzoma (семейство Gleicheniасеае) образуются споры двух типов, которые развиваются соответственно в однополые, т. е. ♂ и ♀ заростки (рис. 11.153, D, Е).
Рис. 11.153. Pteridopsida: А, В — Aspidiales, развитие заростка Matteuccia struthiopteris из споры (70х): А — 11-дневный, В — 21-дневный, с апикальной клеткой (s) и образованными ею сегментами (I—X); С — Hymenophyllales, Trichomanes rigidum, нитчатый заросток с архегониофорами (а), из которых один с проростком спорофита; D, Е — Gleicheniales, Platyzoma microphyllum (20х), ♂ заросток; Е — то же, ♀ заросток
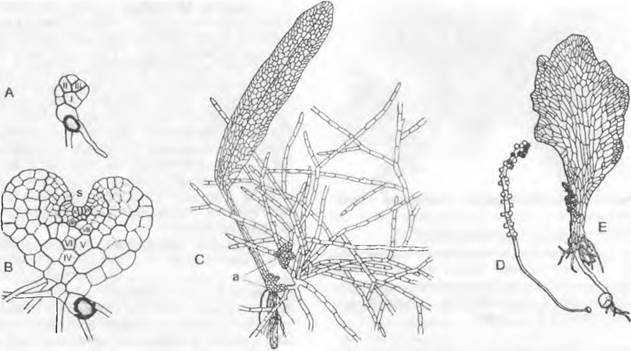
Сначала вырастет нитевидная, снабженная ризоидами «протонема», которая, однако, лишь у немногих форм сильно развита и тогда, например, у Тrichomanes (Нуmenophyllасеае) и Schizaea (Schizaeacea), несет на своих ветвях антеридии, а на особых многоклеточных боковых ветвях — архегонии (рис. 11.153, С). Обычно нитевидная стадия существует очень недолго и, образовав всего несколько клеток, дает на своем конце клиновидную двустороннюю апикальную клетку; отчленяемые ею сегменты делятся дальше (А, В). Таким путем формируется чаше всего сердцевидный, распростертый по субстрату, тонкий, пластинчатый заросток (см. рис. 11.128, А).
Наконец, апикальная клетка заменяется несколькими инициалями. Антеридии и архегонии возникают на стороне, защищенной от падающего света, т. е. обычно на стороне, обращенной к почве и влаге; к концу своего развития они не погружены или слабо погружены в ткань заростка1. Архегонии образуются чаще всего позже, чем антеридии. При очень плохом питании образование архегониев полностью подавлено.
1 Архегонии погружены брюшком в ткань заростка, как и у остальных Pteridophyta (исключение составляют Нуmenophyllacee и Schizaeaceae с нитчатыми заростками). — Примеч. ред.
Антеридий представляет собой округлое выступающее образование, которое без ножки сидит непосредственно на одной эпидермальной клетке; он произошел от нее как сосочковидный вырост, отделенный поперечной перегородкой (рис. 11.154). Образуемые в нем сперматозоиды изогнуты в виде штопора и имеют множество жгутиков (F). Как и вообще у всех архегониат, основной объем сперматозоида занимает ядро. Сначала он несет на своем заднем конце пузыревидный остаток цитоплазмы с мелкими пластидами и крахмальными зернами в качестве запасного вещества, который, однако, при проникновении в архегонии отбрасывается.
Рис. 11.154. Pteridopsida, Aspidiales: А — E — развитие антеридия у Dryopteris filix-mas (250х), объяснение в тексте; F — сперматозоид Thelypteris palustris (3000х); G — N — развитие архегония у Dryopteris filix-mas (20х), объяснение в тексте
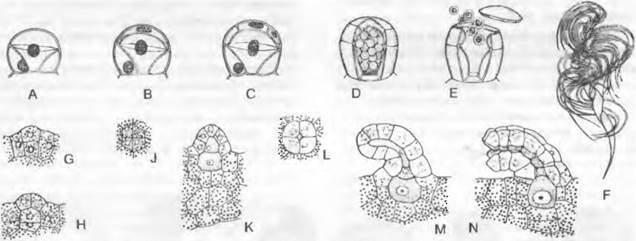
Архегонии возникают в многослойной средней части более взрослых заростков путем деления одной поверхностной клетки каждый. Внутри зачатка архегония образуются шейковая канальцевая клетка и центральная клетка; последняя снова делится на яйцеклетку и брюшную канальцевую клетку (рис. 11.154, М). У некоторых видов может быть несколько шейковых канальцевых клеток (сравнительно-эволюционные аспекты — см. Обзор). Архегонии становится готовым к оплодотворению после разрыва брюшной и шейковых канальцевых клеток, расплывания содержащейся в них слизи и раскрывания шейки на вершине. Сперматозоиды привлекаются к шейке архегония и яйцеклетке (см. 8.2.1.1) по механизму хемотаксиса.
После первых делений зиготы (рис. 11.155, А) апекс побега (s) зародыша дифференцируется эндоскопически рядом с будущей гаусторией (f), а зачатки первого листа (b) и корня (w) обращены к шейке архегония. Корень возникает на лишенном подвеска (суспензора) зародыше не против побега, а сбоку от продольной оси зародыша, как и у всех папоротникообразных. Поскольку архегоний находится на нижней стороне заростка, то побег и первый лист зародыша после выхода из архегония должны, проявляя отрицательный геоторопизм, изгибаться вверх (В). Спорофит остается еще некоторое время связанным через гаусторию (f) с заростком, пока тот не отомрет. К первичному корню позже добавляются многочисленные придаточные корни. Положение оси зародыша не может быть изменено ни силой тяжести, ни светом; следо
вательно, уже заросток у Leptosporangiatae должен проявлять полярность, которая затем передается цитоплазме яйцеклетки.
Рис. 11.155. Pteridopsida, Pteridales Pteridium aquilinum. Развитие зародыша: А — после первых делений зиготы в архегонии; В — на более поздней стадии гаустория погружена в расширенное брюшко архегония (а); b — первый лист; f— гаустория: р— заросток; s — апекс побега; w — корень
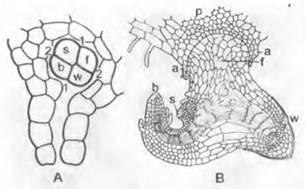
На листьях нередко встречаются адвентивные (выводковые) почки, которые отпадают и участвуют в вегетативном размножении; вегетативному размножению служит также превращение стеблей и даже листьев в ползучие столоны. У некоторых видов наблюдается отклонение от нормального хода смены поколений из-за апогамии и апоспории (см. 10.1.3.3); чаще всего это бывает у полиплоидных форм, многие из которых у Pteridopsida имеют большие хромосомные числа.
Систематика. Классификация многочисленных видов лептоспорангиатных папоротников основана на различиях в строении спорангиев и их положении на спорофилле. На основе этих признаков различают несколько порядков.
У представителей первых трех порядков (1 — 3) кольцо на спорангиях отсутствует или оно поперечное.
Рис. 11.156. Leptosporangiatae. Спорангии: А — Osmunda regalis (Osmundales, устье открыто, 40х); В — Anemia caudate (Schizaeales); С — Нуmеnophyllum dilatatum (Hymenophyllales); D — Dryopteris filix-mas (Aspidiales, устье в плане и сбоку) (В — D — 70х)
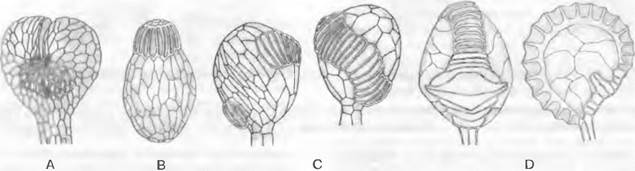
1. Порядок: Osmundales. Сорусы отсутствуют, а спорангии без кольца: стенка спорангия разрывается на его вершине благодаря группе толстостенных клеток (рис 11.156, А). Индузия и чешуй нет. Заростки долговечные, часто даже многолетние. Виды этого порядка, объединяемые в одно-единственное семейство (Оsmundасеае), имеют спорангии либо на особых спорофиллах (Osmunda cinnamomea), либо на определенных участках трофофиллов: у осмунды королевской (О. regalis) видоизменены верхние перья вай, в остальном имеющих нормальный облик, у О. сlaytoniana — средние. Семейство известно с верхнего карбона, в настоящее время представлено немногими родами.
2. Порядок: Сleicheniales. Сидячие спорангии имеют кольцо, проходящее поперечно выше середины. Они собраны небольшими группами в сорусы, которые, однако, не защищены покрывальцем. В ископаемом состоянии представители этого порядка известны с верхнего карбона; в настоящее время они широко распространены в тропиках. Вайи (ложно-)дихотомически ветвящиеся, со «спящими» почками в разветвлениях (рис. 11.157). Принадлежность рода Platyzoma (с выраженной гетеро- спорией, см. рис. 11.153, D, Е) к этому порядку спорна.
Рис. 11.157 Pteridopsida, Gleicheniales, Gleichenia circinata, Австралия (1/5х)
3. Порядок: Schizaeales. Расположенные по краю листа сидячие спорангии вскрываются продольной щелью посредством находящегося у самой верхушки поперечного кольца (см. рис. 11.156, В). Этот порядок также известен в ископаемом состоянии с верхнего карбона; в настоящее время его распространение ограничено преимущественно тропиками. Листья уSchizaea (-сеае) напоминают листья злаков, но дихотомируют, у Anemia (-сеае) они перистые с фертильной нижней парой перьев, у Lygodium (Lygodiасеае) — вьющиеся.
Следующие три порядка (4 — 6) имеют косо проходящее кольцо, как, например, у Нymenophyllum (см. рис. 11.156, С) из 4-го порядка.
4. Порядок: Hymenophyllales. Спорангии почти сидячие. Сорусы располагаются по краю листа, каждый на рецептакуле (продолжении листовой жилки), иногда сильно удлиненном, и защищены чашевидным или двустворчатым покрывальцем. Листья чаще всего тонкие, с однослойной пластинкой без устьиц. Достоверные ископаемые остатки известны только с третичного периода. Ныне живущих видов известно около 650. Они обитают преимущественно во влажных лесах тропи ков и субтропиков, как, например, виды родов Hymenophyllum(однако Н. tunbrigense очень редко, но встречается в Европе) и Didymoglossum.
5. Порядок: Мatoniales. Сидячие спорангии собраны небольшими группами в сорусы, которые покрыты шитовидным покрывальцем. Этот порядок, широко распространенный в мезозое, ныне представлен лишь тремя видами, обитающими на Малайском архипелаге.
6. Порядок: Суatheales (включая Dicksoniales). Расположенные на ножке спорангии собраны в сорусы на поверхности или по краям листьев. Эти растения появились в юре. В настоящее время это преимущественно древовидные папоротники (похожие на пальмы деревья до 20 м высотой) в горных лесах тропиков и субтропиков. Представленные многими видами роды: Cyathea (включая Alsophila, см. рис. 11.150), Dicksonia, Cibothium.
У следующих порядков спорангии, чаще всего имеющие хорошо выраженную ножку, вскрываются благодаря продольному кольцу (рис. 11.156, D), в редких случаях— вертикальному кольцу.
7. Порядок: Роlypodiales. Сорусы образуются на нижней стороне листа; покрывальце отсутствует. Вайи у многолетних растений перисторассеченные или однажды перистые. Сюда наряду с родом Роlypodium относят также роды Drynaria (см. рис. 11.162, В), Рlatycerium (см. рис. 11.162, А), Мicrosorium и Рyrrosia.
8. Порядок: Рteridales. Сорусы расположены по краям перьев вай. Обычный представитель этого порядка — орляк, Рtегidium aquilinum (Dennstaedtiaceae), с дважды- и четыреждыперистосложными вайями до 2 м длины и длинными ползучими корневищами. Сорусы прикрыты с одной стороны закрученным краем листа, а с другой — покрывальцем. К тому же семейству относится род Аcrostichum. У Adianthum (Аdianthaceae) наблюдается примитивное вееровидное жилкование листьев (см. рис. 4.60, А), закрученный край которых защищает сорусы; индузий у них отсутствует. Сеratopteris (Parkeriaceae) — водное растение со стерильными плавающими листьями и фертильными водными листьями, которые несут не собранные в сорусы шаровидные, почти сидячие спорангии с вертикальным кольцом. Сюда относят также семейства Gymnogrammaceae с родом Anogramma, Sinopteridaceae с родами No- tholaena и Cheilanthes и Davalliaceae с родом Dаvallia.
9. Порядок: Aspidiales. Сорусы, защищенные покрывальцем, располагаются на нижней стороне листа. Листовая пластинка изредка простая или лопастная, а чаще всего одно-, четыреждыперистая. К этому порядку, включающему несколько семейств, относится большинство видов папоротников, известных в Германии. У видов семейства Thelypteridaceae (Thelypteris) сорусы сидят вдоль центральной жилки перышка, у Aspleniaceae (Asplénium, Celerach) — по бокам конечных жилок перышек. К последнему семейству относят также листовик, Phyllitis, с цельными языковидными вайями (см. рис. 11.151. С). Представители Aspidiaceae часто образуют щитовидные или почковидные индузии (например, Bryopteris, см. рис. 11.12, С: Bolbitis). К ним очень близки Athyriaceae с часто удлиненными сорусами (как у кочедыжника женского, Athyrium filix-femina). У также относящегося к предыдущему семейству страусника, Matteuccia struthiopteris. как и у представителей следующего порядка, спорофиллы отличаются от зеленых трофофиллов. которые воронковидно налегают друг на друга.
10. Порядок: Blechnales. У единственного встречающегося в Германии представителя этого порядка блехнума колосистого (Blechmim spicant) несколько темно-коричневых спорофиллов располагаются в розетке зеленых трофофиллов, сорусы вытянуты на всю длину узких фертильных перышек. Кроме рода Blechnum сюда же относят род Salpichlaena.
Ступень развития: водные папоротники (Hydropterides) (= Salviniidaе)
К водным папоротникам относят всего несколько родов трав, обитающих в воде или на болотах. Они все разноспоровые. Их мега- и микроспорангии тонкостенные, не имеют кольца и находятся внутри особых вместилищ, расположенных в основании листа (например, рис. 11.158, С). Мейоспоры покрыты своеобразным периспорием, возникающим из плазмодиального тапетума.
Рис. 11.158. Hydropterides, Salviniales. Salvinia, S. natans:
A — участок побега сбоку, видны округлые вместилища спорангиев (3/4х); В — то же, вид сверху (3/4х); С — вместилища мегаспорангиев (mа) и микроспорангиев (mi) в продольном сечении (8х); D — микроспорангий (55х); Е — микроспоры, погруженные в пенистую массу (250х); F — мегаспорангий с мегаспорой, окруженной периспорием (р), в продольном сечении (55х); G — J — ♂ заросток: G — деление микроспоры на три клетки I — III (860х), Н — зрелый заросток сбоку, J — вид с брюшной стороны. Клетка I поделилась на проталлиальные клетки pz1 и pz2 (pz1 — нефункционирующая ри- зоидальная клетка); клетка II — на стерильные клетки sz1, sz2 и две сперматогенные клетки sp1, каждая из которых образует 2 сперматозоида; клетка III — на стерильные клетки sz3, sz4 и две сперматогенные клетки sp2. Клетки sp1 sp1 и sp2 sp2 представляют собой два антеридия (в состав антеридия входят не только сперматогенные клетки, но и клетки стенки. — Примеч. ред.); клетки sz1—sz4 — их стенки; цифры 1 — 1 и 2 — 2 отмечают положение первых клеточных стенок (640х); К— зародыш (е) на продольном срезе, заросток (рг) с хлоропластами, b1 — b3— первые листья (100х); аг — остаток архегония; es — экзоспорий; f — гаустория; s — споровая клетка; ss — апекс побега; sw — стенка спорангия; wb — водный лист

К водным папоротникам относят два порядка примерно со 100 видами.
1. Порядок: Salviniales (единственное семейство Salviniaceae). Относящиеся к этому порядку виды — свободно плавающие водные растения. Род Salvinia во флоре Германии представлен ставшим редким плавающим папоротником S. natans, который на каждом узле слабо разветвленного побега несет по три листа.
Два верхних зеленых листа, плавающих на поверхности воды, овальные (рис. 11.158, А), со множеством крупных межклетников. Нижние листья (wb), напротив, разделены на свисающие в воде многочисленные бесцветные нитевидные лопасти, покрытые волосками. Они выполняют функцию отсутствующих корней (гетерофиллия1, см. 4.3.2). В основаниях водных листьев сидят по нескольку шаровидных вместилищ спорангиев (А); они заключают в себе спорангии, возникающие на столбиковидной плаценте (С).
1 Правильно: анизофиллия. — Примеч. ред.
По своему заложению плацента соответствует модифицированной лопасти водного листа, тогда как покров вместилища можно рассматривать как двухслойное покрывальце. Оно возникает в форме кольцевого валика, который в виде чаши и, наконец, сферы обрастает плаценту с расположенным на ней сорусом спорангиев и затем плотно смыкается на вершине.
Каждое такое вместилище содержит один сорус — либо из более многочисленных микроспорангиев, либо из мегаспорангиев в меньшем числе (рис. 11.158, C:mi, mа); определение пола заростка осуществляется, таким образом, дипломодификационно. Спорангии обоих типов расположены на ножках и имеют в зрелом состоянии однослойную стенку (D, F); в них после редукционного деления возникают мейоспоры.
Микроспорангии содержат 64 микроспоры, образующиеся в тетрадах. Они погружены в пенистое, твердеющее вещество (периспорий, рис. 11.158, Е).
Из каждой микроспоры развивается короткий трубковидный ♂ заросток, который состоит всего из немногих клеток. Он образует только 2 антеридия (рис. 11.158, Н). Каждый антеридий образует (из 2 сперма- тоге иных клеток, дающих начало 4 сперматидам) 4 сперматозоида; они выходят наружу через разрыв в клеточных стенках. Таким образом, заросток сильно редуцирован. Его развитие происходит внутри спорангия, который не вскрывается. Вместо этого заростки вытягиваются наподобие пыльцевой трубки, локально прободают его стенку, высвобождая сперматозоиды.
Мегаспорангии крупнее, чем микроспорангии, и также имеют однослойную стенку (рис. 11.158, F), однако содержат только одну мегаспору, так как лишь одна из 32 возникающих (из 8 материнских клеток спор) спор завершает свое развитие за счет остальных. Мегаспора плотно заполнена белковыми тельцами, капельками масла и крахмальными зернами; на ее вершине находится более плотная цитоплазма с ядром. Коричневая оболочка (экзоспорий) покрыта толстым ячеистым покровом — периспорием. Мегаспора остается внутри спорангия, отделяется вместе с ним от материнского растения и плавает по поверхности воды. На вершине микроспоры при прорастании возникают состоящий из мелких клеток♀ заросток (рис 11.158, К) и лежащая позади него крупная клетка (s), богатая запасными веществами: она служит для питания заростка и не делится дальше, хотя ее ядро в результате митозов дает многочисленные постенно расположенные дочерние ядра1.
1 И мелкоклеточная ткань, и крупная многоядерная клетка представляют собой части одного заростка. — Примеч. ред.
Оболочка споры вскрывается тремя створками, стенка спорангия также разрывается, и заросток выставляется наружу в виде маленького дорсовентрального образования. Хотя он и содержит хлоропласты, однако использует запасные вещества в крупной клетке (s). Он развивает несколько архегониев, однако только одна яйцеклетка после оплодотворения дает зародыш, который своей гаусторией внедряется в расширяющееся и наконец разрывающееся брюшко архегония (рис 11.158, К). Если ни один из архегониев не оплодотворен, то образуются новые архегонии.
Второй род — Azolla — преимущественно тропический; эти изящные, обильно разветвленные плавающие растеньица несут тесно налегающие друг на друга листья, расположенные в два рада, а на нижней стороне побега — длинные корешки (рис. 11.159, А).
Рис. 11.159. Hydropterides, Salviniales. Azolla:
А — растение, вид сверху (4х); В — вершина побега, сверху (12х); С — то же, в поперечном сечении (12х); D — продольный срез верхней лопасти листа; в полости Anabaena azollae(70х); Е — ♂ (вверху) и ♀ (внизу) сорусы (20х); F — микроспорангий (65х), G — окруженный индузием мегаспорангий, содержащий мегаспору с поплавками (65х); Н — часть массулы с глохидиями (160х); J — мегаспора, у которой удалена верхняя половина индузия, чтобы были видны поплавки, к эписпорию прикрепились три массулы посредством глохидиев (65х)

Каждый лист разделен на две лопасти, из которых верхняя находится на плаву и ассимилирует, а нижняя погружена в воду и участвует в ее поглощении (А, В); кроме того, на отдельных боковых ветвях нижние лопасти листьев преобразованы во вместилища спорангиев и прикрыты выростом лопасти. В полостях верхней лопасти в качестве симбионта обитает цианобактерия Anabaena azollae, связывающая атмосферный азот; поэтому Azolla используют на рисовых полях как зеленое удобрение (ср. Суanobacteriota). Azolla интересна своими приспособлениями к обеспечению надежного оплодотворения. После выхода из микроспорангия 64 микроспоры оказываются размещенными по нескольку в 5 — 8 образованных периплазмодием округлых комках, способных плавать, — так называемых массулах. Каждая массула имеет на поверхности выросты в виде двусторонних крючков на ножке — глохидии (Н, J), которые также возникают из вещества периплазмодиального тапетума. Эти крючки служат для прикрепления к мегаспоре, которая перемещается в воде с помощью особого поплавка, содержащего воздух (0,1). Он образуется из сильно вакуолизированного периплазмодия на верхушке мегаспорангия. Заросток развивается, как и у Salvinia.
2. Порядок; Marsileales. К порядку относят роды, виды которых обитают на заболоченной почве. Род Marsilea (Маsilеасеае), который до недавнего времени был представлен в Германии видом М. quadrifolia, ныне исчезнувшим (рис. 11.160, А), характеризуется ползучим разветвленным корневищем с расположенными поодиночке листьями на длинных черешках, несущими очень близко сидящие пары перышек. Свойственные листьям никтинастии не встречаются у остальных папоротников. Над основанием черешка возникают попарно, а у других видов в большем числе, овальные вместилища спорангиев на ножках. В противоположность Salviniales у Маrsileales оболочка каждого вместилища соответствует по своему происхождению ассимилирующему перышку, у которого вследствие усиленного роста нижней стороны зачатки сорусов смещаются внутрь (В). Поэтому эти вместилища называют спорокарпиями.
Рис. 11.160. Hydropterides, Marsileales:
А — Marsilea quadrifolia, внешний вид (2/Зх); В — срез молодого спорокарпия; точками показан зачаток соруса (200х); С — зрелый спорокарпий (8х); D — вскрывшийся спорокарпий М. salvatrix (натуральная величина); Е — проросшая микроспооа с двумя антеридиями (150х); F — сперматозоид (700х); G — архегоний (150х); Н — Pilularia globulifera, внешний вид (2/Зх); g — кольцо хрящевидной ткани; I — проводящий пучок; те — мегаспорангий; mi — микроспорангий; s — спорокарпий; ss — камеры с сорусами
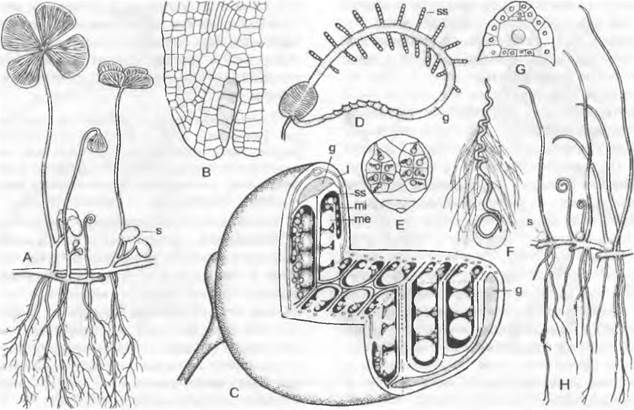
Сорусы, состоящие из мегаспорангия и многочисленных микроспорангиев, располагаются рядами, которые заключены в камеры (С). При созревании спор кольцо хрящевидной ткани (g), опоясывающее спорокарпий, расплывается и выдергивает наружу камеры с сорусами (D). Мужской заросток, который остается заключенным в оболочке микроспоры (Е), образует два антеридия, в которых возникают сперматозоиды (F), спирально закрученные подобно штопору. Женский заросток развивает один архегоний (G). Род Pilularia с встречающимся в Германии видом Р. Globulifera отличается от Marsilea простыми линейными листьями, у основания которых поодиночке возникают шаровидные спорокарпии, по происхождению также соответствующие ассимилирующему перышку (рис. 11.160, Н). В спорокарпиях Pilularia имеется четыре камеры с сорусами. Листья видов обоих родов, как и у большинства других папоротников, в молодости закручены улиткой (А, Н; акропластический рост, см. 4.3.1.2).
В ископаемом состоянии Azolla известна с нижнего мела, Salvinia — с верхнего мела и Pilularia — с миоцена. В Северной Америке Salvinia в миоцене вымерла.
Распространение и образ жизни папоротникообразных
Папоротникообразные распространены во всех климатических зонах, однако наибольших размеров (древовидные папоротники!) и наибольшего видового разнообразия они достигают — прежде всего Pteridopsida и Lycopodiopsida — в тропиках. Как и бриофиты, папоротникообразные предпочитают более влажные места, однако отдельные виды заходят и в сухие области. Засоленных мест они избегают; только папоротник Acrostichum aureum обитает в мангровых болотах всех тропических областей.
Папоротникообразные в ходе приспособления к различным условиям жизни на суше приобрели целесообразные морфологические и физиологические особенности и представлены теми же жизненными формами (см. рис. 4.19), что и семенные растения. По особенностям водного режима они занимают промежуточное положение между бриофитами и семенными растениями. Заростки большинства папоротникообразных столь же малоустойчивы к высыханию, как и протонемы мхов. Однако взрослые зеленые растения папоротников у большинства видов сами регулируют содержание воды в теле (гомойогидридные птеридофиты) — в противоположность полностью зависящим от содержания воды во внешней среде и устойчивым к высыханию бриофитам. Правда, у некоторых пойкилогидридных видов selaginella (Lycopodiopsida) и папоротников (Рteridopsida), например, у Ceterach, Notholaena и Cheilanthes, вечнозеленые листья могут высыхать до воздушно-сухого состояния и после пребывания в нем в течение многих месяцев при увлажнении восстанавливать жизнедеятельность.
Система внутреннего проведения воды играет решающую роль в поддержании водного режима; некоторые папоротникообразные дополнительно используют приспособления к капиллярному поглощению и накоплению воды (язычки у Selaginellaceae и Lepidodendrales).
Очень немногие ксерофиты, например, из Pteridopsida, защищены от высыхания восковым налетом, покровом из сухих чешуек и волосков или даже имеют суккулентные побеги (Davallia) или листья (например, некоторые виды Polypodium). У обитателей влажных мест (гигрофитов) мы наблюдаем гуттацию, будь то через гидатоды на зубцах листовых влагалищ Equisetum или через своеобразные гидропоты у некоторых папоротников (рис. 11 161).
Рис. 11.161. Leptosporangiatae, Polypodiaceae. Polypodium vulgare, гидропота (80x)
Наряду с вечнозелеными видами родов Lycopodium, Selaginella (Lycopodiopsida), Equisetum (E. hyemale', Equisetopsida) и Polypodium (Pteridopsida) значительная часть папоротникообразных умеренной и холодной зон летнезеленые (ср. в этом плане бриофиты). Папоротникообразные в разных эволюционных линиях, некоторые уже в девоне и карбоне, выработали древовидные формы (фанерофиты; жизненные формы, см. рис. 4.19); например, среди Lycopodiopsida — Sigillaria, Lepidodendron, Pleu- romeia; среди Equisetopsida — Catamites; среди Pteridopsida — различные Primofilices, эуспорангиатные (Megaphyton, рис. 11.148, А) и лептоспорангиатные папоротники (Cyathea. см. рис. 11.150). Предпосылками к этому были укрепление осей (отчасти благодаря вторичному утолщению, но чаще посредством других приспособлений и эффективной конструкции проводящих тканей. К фанерофитам относятся также распространенные, особенно в тропиках, лазающие (лианы) и эпифитные папоротникообразные; например, тропические Gleicheniaceae(лазящие растения); Lygodium и Salpichlaena со вьющимися рахисами до 15 м длиной; отдельные виды рода Polypodium, взбирающиеся с помощью корней на стволы деревьев в тропиках; Platycerium и Drynaria как эпифиты со стерильными листьями, образующими нишу для накопления гумуса (рис. 11.162). Есть основания предполагать, что все ныне живущие птеридофиты произошли от вечнозеленых древовидных предков с исходно автотрофными заростками. В дальнейшем возникли жизненные формы: хамефиты, некоторые из них имеют микотрофные (Lycopodium), а другие — существующие за счет запасов споры заростки (Selaginella), гемикриптофиты с автотрофными, недолговечными заростками (например, Dryopteris), геофиты с микотрофными (например, Ophioglossum) или автотрофными заростками (например, Pteridium, Equisetum) и терофиты (например, Anogramma).Если не считать заростков некоторых видов и в значительной мере микотрофного Ophioglossum simplex, то к гетеротрофному образу жизни (паразитизм) папоротникообразные, как и мхи (однако ср. Cryptothallus, Bryophytina), не перешли. Немногие представители вторично приспособились к жизни в воде (как гидрофиты): например, из Pteridopsida — Salvinia и Azolla (Salvi-
niales) как плавающие растения, а также Ceratopteris (Pteridales). Некоторые виды Ceratopteris живут как плавающие или в той или иной мере погруженные в воду, а некоторые обитают на влажной почве. Культивируемые в аквариумах Bolbitis heudeloitii и Microsorium pteropus образуют под водой только стерильные вайи, тогда как сорусы развиваются исключительно на выступающих из воды листьях. Виды рода Isoétes (Lycopodiopsida) обитают частью на периодически заливаемой почве, частью на дне озер, нередко на глубине 1 — 3 м.
Рис. 11.162. Эпифитные папоротники с диморфными листьями (гетерофиллия: стерильные листья, образующие нишу для накопления гумуса, и споротрофофиллы):
А — Platycerium alcicorne; В — Drynaria quercifolia (1/6х)
Папоротникообразные особенно конкурируют с особями сходных жизненных форм: древовидные папоротники, например, — с голосеменными, пальмами и древесными двудольными, заростки папоротников, а также спорофиты Hymenophyllaceae — со мхами и лишайниками; Equisetum — с Juncaceae и Cyperaceae; Salvinia — с Lemnaceae и т. д. Некоторые папоротникообразные при этом оказываются весьма конкурентоспособными и в благоприятных условиях встречаются в таких количествах, что образуют чистые заросли, как орляк (Ptertuium aquilinum) по опушкам лесов или хвощ приречный (Equisetum fluviatile) по топким берегам озер. Некоторые виды распространены по всему земному шару, например, тот же орляк (как лазящее растение иногда до 4 м высотой) или плаун булавовидный (Lycopodium clavatum): можно привести примеры и ограниченных, разорванных и эндемических ареалов.
Обзор папоротникообразных. Папоротникообразные можно рассматривать как эволюционную ветвь, возникшую параллельно мохообразным от общей, возможно, уже обитавшей на суше, предковой группы («Propsilophytopsida»), которая в свою очередь произошла от водорослевых предков. Среди водорослей в качестве предков этих ранних обитателей суши подходят только стрептофиты. Ранее обсуждавшаяся эволюция от мохообразных к папоротникообразным, например, от форм типа Anlhocems, путем увеличения размеров, дифференциации и увеличивающейся самостоятельности спорофита (Sporogonites), напротив, представляется маловероятной. Если мхи уже с карбона не претерпевали заметного прогрессивного развития, т. е. уже примерно 250 млн лет назад были «готовыми», папоротникообразные достигли расцвета только после этого времени.
Если мхи завоевали сушу с помощью гаметофитов и при этом остались ограниченными особыми адаптивными зонами, то папоротникообразные и (прежде всего) семенные растения достигли доминирующей роли в формировании наземной растительности при прогрессирующем развитии своих спорофитов. Эволюционное преимущество спорофитных растений связано, вероятно, с их генетической стабильностью (буферный эффект) и повышенной частотой рекомбинаций (после каждого события сингамии возникающие диплоидные ядра воспроизводятся митотически; вследствие этого в мейоз вступают многочисленные диплоидные ядра), а также с развитием защитных приспособлений для чувствительного к наземным условиям гаметофита. Диплоидный спорофит папоротникообразных развит очень сильно и весьма разнообразен, что в противоположность мхам стало возможным благодаря прогрессирующему развитию в ходе эволюции одревесневших (соответственно эффективно работающих) проводящих пучков (проведение как воды, так и органических веществ1). Аналогичный результат имеет также развитие настоящих корней. Поскольку к тому же эпидерма кутинизирована, то побег может расти вверх в освещенное воздушное пространство, развивать листья и ассимилировать диоксид углерода. Его обеспечение органическими веществами, таким образом, не зависит от гаметофита, из-за чего отпадает необходимость в дальнейшем росте последнего.
1 Автор этого раздела, по-видимому, своеобразно представляет роль разных тканей в проведении тех или иных веществ, равно как и значение одревеснения. К высказанной автором идее, по-видимому, следует относиться с большой осторожностью. — Примеч. ред.
Верхушечный рост осуществляется чаще всего посредством апикальной клетки (рис. 3.2, А). Напротив, благодаря множественным инициалям растут эвспорангиатные папоротники и некоторые представители Lycopodiopsida (Lycopodium, Selaginella— у последнего рода еще отчасти посредством апикальной клетки). Среди Psilophytopsida этим прогрессивным признаком отличался уже род Rhynia2.
2 Строение верхушки телома у Rhynia точно не известно. — Примеч. ред.
Положение зародыша (см. рис. 11.155) чаще всего эндоскопическое (верхушка побега направлена от шейки архегония — Leptosporangiatae, а также Lycopodium, Selaginella), редко экзоскопическое (Eusporangiatae, за исключением Marattiales, Psilotum, Equisetum, Isoetes).
У исходных форм все споры одинаковые (изоспория); у более высокоорганизованных уже появляется дифференциация на микро- и мегаспоры. Гетероспория неоднократно и независимо появлялась в разных классах папоротникообразных (Lycopodiopsida; Equisetopsida — как у каламитов, так и у клинолистных, и Pteridopsida); с этим связано разделение функций между мелкими ♂ заростками и более крупными заростками. В виде «семенных плаунов» папоротникообразные независимо от семенных растений достигли такого уровня развития, как образование семени с крайней степенью гетероспории.
Гаплоидный гаметофит папоротникообразных остается (если еще не слишком сильно редуцирован) на уровне организации таллома (заросток) и изредка развивает трахеиды (Psilotum). Он рано заканчивает развитие, образовав антеридии и архегонии, которые часто устроены проще, чем у мхов; крупные многоклеточные гаметангии считают примитивными по сравнению с мелкими, состоящими из немногих клеток. Если листостебельные мхи (Вryopsida) имеют в архегонии многочисленные шейковые канальцевые клетки (10 — 30 или больше), то у печеночников (Мarchantiopsida) их 4 — 8, у антоцеротовых (Аnthocerotopsida) — 6, а у папоротникообразных часто только одна или несколько. У Аnthocerotopsidaэпидермальная клетка, становящаяся инициалью архегония, больше не образует клетки ножки, т. е. в отличие от Marchantiopsida и Bryopsida она сразу делится на осевую клетку и три клетки стенки архегония. У папоротникообразных нет и делений, дающих клетки стенки архегония1. Антеридии и архегонии Bryopsida и Marchantiopsida закладываются экзогенно и свободны, т. е. только позже окружаются тканью гаметофита. У Anthocerotopsida и папоротникообразных они уже на ранних стадиях развития заключены в ткань гаметофита (эндогенное образование)2.
1 Это утверждение весьма спорное действительности. — Примеч. ред.
2 У всех высших растений (кроме антеридиев Anthocerotophyta) гаметангии развиваются из поверхностной клетки, т. е. всегда экзогенны. — Примеч. ред.
Рис. 11.163. Развитие важнейших групп растений в истории Земли
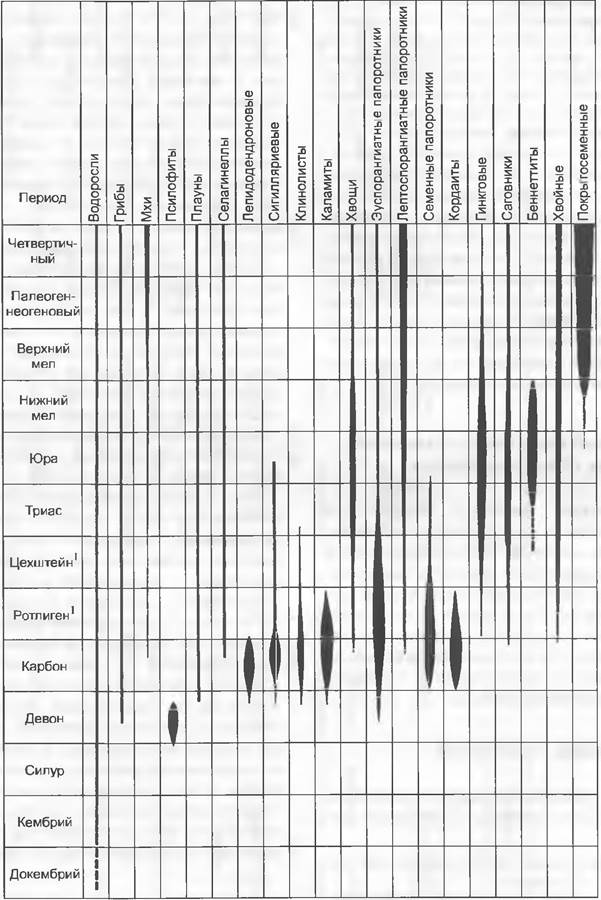
1 Вышедшие из употребления названия.
История развития. Psilophytopsida, Lycopodiopsida и Equisetopsida достигли наибольшего расцвета как по разнообразию форм, так и по численности особей в палеозое. Представительство Pteridopsida значительно возросло еще в мезозое, и они лучше сохранились до сих пор, чем два других доживших до нашего времени класса (рис. 11.163). Их формы, господствовавшие от карбона до триаса, в настоящее время сохранились лишь в виде немногих видов, тогда как те семейства, которые доминируют в настоящее время, появились только в мезозое.