БОТАНІКА З ОСНОВАМИ ГІДРОБОТАНІКИ (ВОДНІ РОСЛИНИ УКРАЇНИ) - Б.Є. Якубенко - 2011
VI. СИСТЕМА РОСЛИННОГО СВІТУ
НИЖЧІ СПОРОВІ РОСЛИНИ
ЗАГАЛЬНА ХАРАКТЕРИСТИКА ТА ОСОБЛИВОСТІ
Відділ Ціанопрокаріоти (Синьозелені водорості) — Cyanoprokaryota (Cyanophyta, Cyanobacteria)
Ціанопрокаріоти (синьозелені водорості) - об'єднують прокаріотичні фотоавтотрофні рослинні організми та налічують близько 2000 видів. Це одна з найдавніших груп організмів на Землі, їхній вік становить близько 3,5-3,8 млрд. років. Вони подібні за своєю будовою до фотосинтезуючих бактерій, проте мають низку характерних особливостей чи відмінностей: 1) облігатні фототрофи, які не здатні (за винятком окремих видів) засвоювати екзогенні органічні сполуки; 2) вирізняються наявністю двох фотосистем, розміщених на мембранах спеціалізованих фотосинтезуючих структур - тилакоїдів, переважно не з’єднаних з плазмалемою; 3) продукування кисню пов’язано з фотолізом води, який здійснюється за рахунок енергії сонячного світла; 4) відсутність дихання на світлі; 5) можливість проходження в клітині двох процесів - оксигенного фотосинтезу та анаеробної азотфіксації в мембранах тилакоїдів, які виконують функції пластид та мітохондрій почергово, а також відсутність джгутиків та рухливих стадій.
Зовнішня морфологічна подібність до бактерій, прокаріотична клітинна організація та подібність геномного складу надають підставу деяким вченим віднести ці організми до царства бактерій як ціанобактерії та підпорядкувати їхньому кодексу бактеріальної номенклатури. Проте, враховуючи зазначені раніше особливості та специфіку цих організмів, доцільно їх розглядати серед інших рослин царства Plantae як прокаріотичних представників і відповідно номенклатурним принципам Міжнародного кодексу ботанічної номенклатури.
Ціанопрокаріоти (синьозелені водорості) надзвичайно поширені у різноманітних водних та позаводних біотопах і мають широку екологічну амплітуду та пристосування до існування за різних умов зростання. Спектр їхнього забарвлення охоплює діапазон від синьо-зеленого (властивого для більшості представників) до жовто-зеленого, зеленого, сіро-сталевого, червоного чи чорного.
Характер забарвлення обумовлений умовами зростання та проявом пігментного складу, до якого входить хлорофіл a (як виняток — хлорофіл b) , фікобілінові пігменти - фікоціанін, алофікоціанін, фікоеритроціанін та фікоеритрин (червоний пігмент); каротиноїди - β-каротин, ксантофіли лютеїнового циклу (лютеїн та зеаксантин) і специфічні ксантофіли - осцилоксантин, міксоксантин, афаніцин та афанізофіл).
Хлорофіл b (хлорофілоподібний пігмент - дивініл-хлорофіл а- подібний Mq-порфірин), a інколи також α-каротин властиві для групи зелених прокаріотичних водоростей (її розглядали раніше в ранзі Prochlorophyta), приналежність яких до ціанопрокаріот доведена молекулярно-генетичними дослідженнями.
Основним продуктом асиміляції ціанопрокаріот є глікогеноподібний полісахарид - крохмаль синьозелених водоростей, а також ціанофіцин та волютин.
Синьозелені водорості - одноклітинні, колоніальні або багатоклітинні організми, нерідко оточені слизом, формуючи пухкі слизисті угруповання. Для одноклітинних та колоніальних форм характерний кокоїдний тип тіла, а для багатоклітинних - нитчастий, інколи спостерігається тенденція до пластинчастого чи об’ємного розміщення клітин.
Сукупність фізіологічно пов'язаних клітин в нитчасту структуру (розгалужену чи не нерозгалужену) називають трихомом. Ці клітини поєднані між собою за допомогою плазмодесм, які проходять через пори поперечних перетинок клітин. Окремі трихоми чи їхня сукупність можуть знаходитись в слизовому утворі - піхві.
Трихоми бувають нерозгалуженими (клітини діляться лише в одній площині) та розгалуженими (клітини здатні до поділу в кількох площинах). Останній тип галуження трихом називають справжнім, а якщо галузяться нитки, а самі трихоми лишаються нерозгалуженими, то такий тип галуження вважають несправжнім. Трихоми розрізняються також за функціями та формою клітин: гомоцитні - сформовані однотипними не диференційованими вегетативними клітинами та гетероцитні - клітини різні за формою та своїми функціями. До складу гетероцитних трихомів входять вегетативні клітини, гетероцисти (виконують функцію фіксації атмосферного азоту в аеробних умовах) та акінети (спочиваючі клітини, за допомогою яких водорості переносять несприятливі умови). Інколи у нитчастих форм присутні верхівкові клітини, за рахунок ділення яких відбувається ріст таломів.
Форма клітин ціанопрокаріот - різноманітна, але найчастіше трапляється сферична чи напівсферична та еліпсоїдна, менш поширена - веретеноподібна, видовжена, грушоподібна тощо. Розміри вегетативних клітин від 0,2 до 10 мкм.
Вегетативні клітини, що здійснюють фотосинтез та здатні до поділу, містять протопласт, оточений особливими оболонками - клітинна оболонка, капсули, слизові обгортки.
Клітини ціанопрокаріот мають типову прокаріотичну будову: вони позбавлені морфологічно оформленого ядра, мітохондрій, хлоропластів, ендоплазматичної сітки, комплексу Гольджі, лізосом, а також джгутиків. Ці водорості не здатні до мітозу, мейозу та статевого процесу.
Оболонка клітин - двошарова, з яких зовнішній шар побудований переважно з пектинових речовин та мікрофібрил, а внутрішній - утворений
муреїном. Назовні від клітинної оболонки у більшості видів виділяються пектинові речовини, що утворюють колоніальний слиз або слизові піхви (рис. 10). Усі клітинні оболонки (разом зі слизом) є похідними протопласта, що відповідає цитоплазмі, оточеній цитоплазматичною мембраною - плазмалемою, з різноманітними складками.
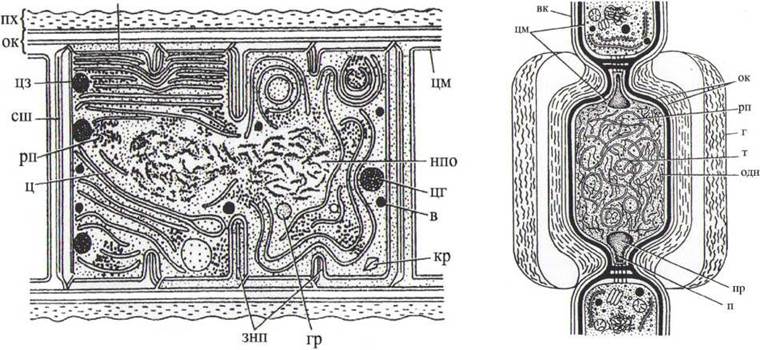
Рис. 10. Схематичне зображення будови вегетативної клітини (А) та гетероцисти (Б) синъозелених водоростей: в - поліфосфатна гранула (волютин); вк - вегетативна клітина; пх - піхва; г - гетероциста; гр - гранула невідомого складу; знп - зачатки нових клітинних поперечних перетинок - септ; ок - оболонка клітини; кр - кристал; нпо - нуклеоплазматична область (нуклеоплазма); п - пора гетероцисти; к - коркоподібний утвір, що замикає канал пори гетероцисти; одн - окремі нитки ДНК; рп - рибосоми та полісоми; сш - середній шар поперечної частини клітинної стінки (поперечна перетинка); т - тилакоїд; ц - цитоплазма; цм - цитоплазматична мембрана; цз - ціанофізоване зерно (структурована гранула).
Сформоване ядро (а також ядерце) відсутнє, а його функції виконує нуклеоїд. Основна маса нуклеоїду зосереджується в центрі клітини, через що цю зону цитоплазми називають нукпеоплазмою або центроплазмою. У цій частині клітини вирізняють особливі структури - хроматинові елементи, які крім ДНК містять основні білки. Фотосинтетичний апарат не організований у вигляді пластид, а представлений тилакоїдами, що є похідними інвагінацій плазмалеми, які не відокремлюються від цитоплазми двомембранною оболонкою. Тилакоїди розташовані поодиноко у оптичному мікроскопі ця зона має інтенсивне забарвлення. На поверхні тилакоїдів є особливі структури - фікобілісоми, що містять фікобілінові пігменти. До елементів фотосинтетичного апарата опосередковано належать також поліедральні тіла, які є попередниками піреноїду еукаріотичних водоростей.
У цитоплазмі ціанопрокаріот знаходяться також рибосоми, газові вакуолі та включення, які представлені гранулами специфічного крохмалю синьозелених водоростей, ціанофіцину та поліфосфатними гранулами. Ймовірно, що великі гранули ціанофіцину (понад 1 мкм) слугують джерелом азоту в разі його дефіциту в середовищі та можуть використовуватись як джерело енергії в період перебування клітини в темноті за анаеробних умов.
Газові вакуолі (псевдовакуолі) відіграють роль «плаваючого міхура» та підтримання клітин в ширяючому стані у воді, а також під час осідання на дно водойми (за несприятливих умов). Типові вакуолі (з клітинним соком) у синьозелених водоростей відсутні, але для окремих видів властива своєрідна зворотня форма вакуолізації (керитомія), в разі відмирання чи паталогічного стану клітин.
В основі процесів розмноження усіх синьозелених водоростей лежить клітинний поділ. За клітинного поділу спочатку плазмалема, а за нею і муреїновий шар клітинної оболонки доцентрово вростають у протопласт, розділяючи клітину на дві (зрідка - більше) рівні або нерівні частини. Цей поділ може бути повним або неповним. За повного поділу плазмалема та клітинна перетинка повністю виокремлюють дочірні клітини одну від одної. У даному разі пори та плазмодесми не утворюються і кожна дочірня клітина представляє собою фізіологічно самостійний індивід. Повний поділ властивий одноклітинним ціанопрокаріотам.
У разі неповного поділу поперечна перетинка не повністю розділяє дочірні клітини і вони лишаються з'єднаними плазмодесмами. Внаслідок такого поділу утворюються багатоклітинні нитчасті трихоми. Короткі рухливі фрагменти трихом, що складаються з 2-50 клітин, називають гормогоніями, а нерухомі - гормоцитами. Гормогонії та гормоцити є спеціалізованими репродуктивними структурами багатоклітинних синьозелених водоростей. Деякі види ціанопрокаріот розмножуються за допомогою гонідій або сформованими всередині клітини ендоспорами чи відшнурованими від верхівки материнської клітини екзоспорами. Колоніальні одно- та багатоклітинні ціа- нопрокаріоти здатні розмножуватися також шляхом фрагментації колоній.
Розмноження синьозелених водоростей потрібно вважати як вегетативне та безстатеве. Типовий статевий процес відсутній, але виявлені парасексуальні процеси, в результаті яких спостерігається часткове об’єднання геномів різних клітин.
Синьозелені водорості здатні фіксувати атмосферний азот. Під час азотфіксації молекулярний азот відновлюється до сполук амонію і в цій формі він включається в основні шляхи клітинного метаболізму. Процес азотфіксації здійснюється звично в анаеробному середовищі, оскільки його каталізує нітрогеназний ферментний комлекс, робота якого повністю інгібується молекулярним киснем. Разом з цим, є чимало видів серед багатоклітинних ціанопрокаріот, що фіксують атмосферний азот також у присутності кисню за рахунок наявності особливих клітин - гетероцист у період активного розвитку.
Синьозелені водорості активно зростають у воді, ґрунті, аерофітних умовах та розвиваються в температурному діапазоні від + 750 С до - 830 С, в товщі води, поблизу донних відкладів, серед обростань різноманітних субстратів, як компоненти лишайників, окремих органів квіткових рослин, безхребетних тварин тощо. Вони першими оселяються на наземних субстратах (витримують надзвичайно високий рівень радіації - 1280 тисяч рентген на годину), розвиваються в прісних водах та морях, належать до абсолютних первинних продуцентів органічної речовини на планеті. Ці водорості, як азотфіксуючі організми, сприяють зростанню продуктивності ґрунту чи збереження родючості цілинних ґрунтів та урожайності сільськогосподарських культур, а за своїми фізіолого-хімічними характеристиками є додатковими об’єктами харчових домішок в раціоні тварин та людини, продуцентами лікарських речовин тощо. Так, види роду Arthrospira (A. platensis та A. maxima), під назвою «спіруліна», є промислово цінними об’єктами, введені в активну культуру і їхню біомасу використовують в різних галузях народного господарства (легка, харчова та медична промисловості), а природна біомаса видів Nostoc слугує харчовим продуктом жителям Центральної Африки.
Із негативних особливостей цих водоростей зазначимо їхню здатність до масового розвитку, що спричиняє «цвітіння» води у водоймах, особливо
слабко проточного типу, за умов високої температури (понад 23-25° С) та підвищеної концентрації біогенних елементів (азоту, фосфору, калію), отже вторинного забруднення водойм і загибель риби, наявність сильнодіючих отрут (токсинів) серед їхніх екзметаболітів тощо. Основними збудниками «цвітіння» води у водоймах України є Microcystis aeruginosa, Anabaena flos- aquae, Aphanizomenon flos-aquae.
Відділ містить два класи - Cyanophyceae та Hormogoniophyceae, які за морфологічними ознаками (тип поділу клітин, особливості слизоутворення, тип трихом та їхня здатність до справжнього галуження) поділяються на порядки - Chroococcales, Oscillatoriales, Nostocales та Stigonematales.