Основи еволюції - Корж О.П. - 2006
Частина I. МІКРОЕВОЛЮЦІЯ
Розділ 8. Адаптація як біологічне явище
8.2. Пристосувальне значення властивостей організмів
Засобами пасивного захисту вважаються такі, сама наявність яких забезпечує збільшення ймовірності виживання відповідних особин. До них можна віднести колючки кактуса, шкаралупу горіха, голки їжака, панцир черепахи, черепашку равлика,
кутикулу комахи тощо. Якщо організми з подібними властивостями мають ще й забарвлення, близьке до загального фону довкілля, то виживаність їх значно зростає.
Переважна більшість тварин має криптичне (захисне) забарвлення, пристосоване до певних умов існування. Причому тваринам різних систематичних груп, які мешкають в однакових умовах, властива конвергенція забарвлення. Систематично близькі організми, розходячись по екологічних нішах, дають приклад адаптивної радіації, набуваючи забарвлення під загальний фон середовища. Інколи той самий вид у різних місцях існування має деякі відмінності в забарвленні (утворюються так звані локальні форми).
Саме забарвлення часто виявляється недостатнім, тому необхідною стає ще й відповідна поведінка. Так, на острові Мартініка в однакових умовах існують три види ящірок роду Anolis - буруваті, зелені та світло-сірі. У стані занепокоєння вони ховаються на тлі відповідного забарвлення: зелені - у зеленій траві, бурі — у засохлих кущах, а світло-сірі - на стовбурах дерев, завдяки чому всі вони стають непомітними. Особлива поведінка стає також необхідною птахам, які гніздяться на землі: вони повинні вибрати таку ділянку, щоб забарвлення яєць максимально наближалось до фону навколишнього середовища.
Ще більш наочними прикладами є здатність багатьох видів тварин змінювати своє забарвлення відповідно до змін довкілля. Ці пристосування можуть бути тимчасовими, сезонними або постійними, специфічними чи такими, що регулюються. Численними є приклади зміни забарвлення в життєвому циклі, що найбільш типово для комах.
У тому випадку, коли організм на різних фазах розвитку мешкає не в одних умовах, кожна фаза набуває свого захисного забарвлення. Так, у багатьох видів метеликів гусениці нагадують гілки рослин, якими вони живляться, лялечки зберігаються в ґрунті і мають відповідне забарвлення, а імаго пристосовані до місць схованок (наприклад - на корі дерев).
Зміни забарвлення в онтогенезі властиві й хребетним тваринам, що може пояснюватися зміною біотопів мешкання на різних етапах їх розвитку. Так вугри (Anguilla anguilla) і деякі інші риби на стадії пелагічної личинки, як і представники зоопланктону, прозорі й не забарвлені. У дорослому стані вони живуть на літоралі (дно водойм) і набувають відповідної пігментації під своє оточення. У птахів і ссавців молодь може мати плямисте чи смугасте забарвлення (перепілка, фазан, олень, свиня тощо), у той час як у дорослих тварин забарвлення зовсім інше.
Подібні пристосування організмів до фону середовища в ході онтогенезу розвиваються поступово, є необоротними і отримали назву морфологічних. Зокрема, у багатьох метеликів, таких, як павиноочка (Vanessa іо), капусниця (Pieris brassicae) та інших, забарвлення лялечок зумовлюється високою чутливістю гусениць до кольору загального тла, на якому вони розвиваються. Тому останні, регулюючи процес пігментації своєї кутикули, виробляють відповідне захисне забарвлення.
Загальновідомою є також здатність саранових до зміни свого забарвлення в широкому спектрі під загальний фон довкілля. Наприклад, акриди навесні зелені і непомітні серед стеблин злаків, а восени вони набувають буро-жовтого забарвлення й нагадують солому.
Деякі комахи Африки набувають захисного забарвлення під степові ділянки, що потерпають від пожеж. Таргани, богомоли, цвіркуни, гусениці тощо в подібних умовах набувають чорного кольору різної інтенсивності.
Здатність до швидкої зміни забарвлення під тло стає важливою для активних тварин. Викликається вона переміщенням пігментних зерен у спеціалізованих клітинах - хроматофорах, що отримало назву «фізіологічна зміна забарвлення». Властива ця здатність відносно високорозвиненим тваринам - молюскам, членистоногим і хребетним. Механізм фізіологічної зміни забарвлення складний, включає рефлекторні дії, що спираються на зорове сприйняття довкілля і гормональну регуляцію, а інколи викликаються прямим впливом світла на шкіру. Проте птахам та ссавцям такі пристосування невластиві, оскільки їх шкіра вкрита пір'ям або волоссям, через що їх колір стає непринциповим.
Усім відомі приклади швидкої зміни забарвлення хамелеонами, але велика кількість видів хребетних і безхребетних тварин також має подібні властивості і навіть більш розвинені, ніж у цих плазунів. Представниця безхребетних каракатиця (Sepia officinalis) володіє надзвичайними здібностями змінювати своє забарвлення - від перламутрово-білого через плямисті варіанти різних відтінків до майже чорного. Крім неї, широку гаму кольорів забарвлення свого тіла — від чорного до світлого - може створювати восьминіг.
У багатьох видів риб (особливо представників коралових рифів) здатність до зміни свого забарвлення просто вражає: деякі види за кілька секунд можуть змінити його шість-вісім разів. Причому вдаються вони до цього не лише в разі переміщення з одного середовища до іншого, а й при вертикальних переміщеннях: піднімаючись із дна, особина майже відразу змінює крапчасте забарвлення на одноманітне. У деяких випадках риби і головоногі молюски під час руху розмальовують себе поздовжніми смугами, що маскує їхній рух уперед, а в стані спокою мають поперечні смуги, завдяки яким наче розчиняються на плямистому тлі.
Змінювати забарвлення можуть і наземні тварини, у першу чергу мешканці дерев, у яких головним захистом залишається їх забарвлення. Зміни відтінків забарвлення в широких межах властиві багатьом амфібіям і рептиліям. Незважаючи на видові особливості кольорових гам і механізми, що їх створюють, головним результатом подібних властивостей цих організмів є наближення забарвлення і навіть малюнка тіла до загального фону середовища існування, що зменшує їх помітність.
Скрадаюча протитінь - ще один спосіб створення захисного забарвлення.
Навіть однотонно забарвлений предмет під променями сонця завдяки взаємодії світла і тіні набуває певної рельєфності і сприймається як об'ємне тіло. Верхня частина, освітлена, як правило, інтенсивніше, здається світлішою, а нижня, навпаки, - темнішою (рис. 8.1 а). Тому навіть організм з повного відповідністю будови і забарвлення тіла навколишньому середовищу може стати помітним.
Принцип протитіні полягає в тому, що внаслідок поступового переходу від темнозабарвленої спини до світлішого черева (рис. 8.16) округлена поверхня справляє враження плоскої. Таке забарвлення компенсує дію сонячних променів, створюючи оптичний ефект відносно однотонного забарвлення (рис. 8.1 в). Особливо добре спрацьовує подібний ефект, коли тварина знаходиться на тлі однакового з нею забарвлення, у цьому випадку весь контур і поверхня зливаються з фоном. Найбільш наочно подібний принцип реалізується в різноманітних видів риб.
За принципом протитіні може «працювати» і певний малюнок (відрізняється від забарвлення тим, що його формують принаймні два кольори з чітким протиставленням). Досягається ефект шляхом зменшення кількості або розмірів плям (дрібні котячі з плямистим забарвленням - цивети, генети, оцелоти тощо) чи зменшенням кількості або ж ширини поперечних смуг (зебри, тигр тощо). У цьому випадку явище протитіні залишається тим самим: спина виявляється забарвленою більш інтенсивно порівняно з черевом, що компенсує дію сонячного світла.
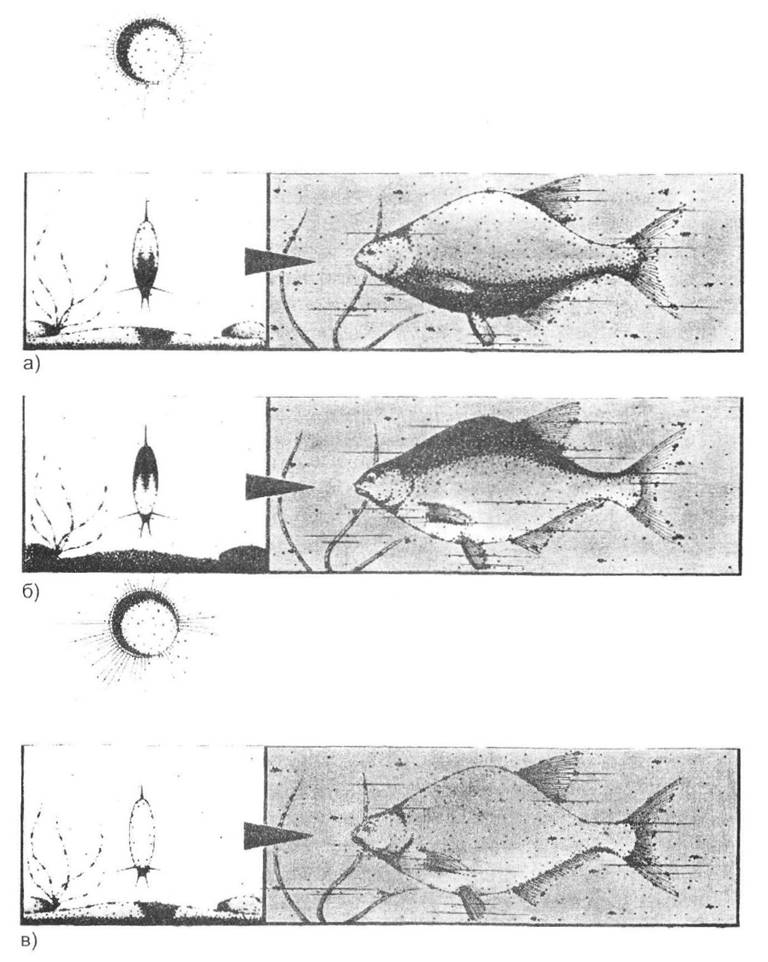
Рис. 8.1. Механізм дії забарвлення за принципом иротитіні на прикладі риби (рис. О.В. Карнаухова):
а - однотонно забарвлена риба під променями сонця набуває об'ємного вигляду через появу тіні на череві; б - значна кількість риб має в природних умовах темніше забарвлену спину та світліше черево; в - таке природне забарвлення під променями сонця компенсує явище тіні й робить тіло риби наче плоским та непомітним
Протитінь стає одним із найважливіших принципів у забарвленні тварин і набуває широкого розповсюдження в природі. Вона розвивалася схожими шляхами в багатьох систематичних групах тварин як на суходолі, так і на морі.
Ступінь вираження протитіні багато в чому залежить від умов освітлення: тварини, що мешкають при гіршому освітленні (донні риби, лісові мешканці), мають слабкий прояв протитіні, оскільки відмінності в освітленні спинної і черевної частин тіла незначні. Тварини пустелі також мають незначний контраст у забарвленні, що пов'язано з відбиваючою здатністю піску. На значних глибинах (або в печерах), де світло взагалі відсутнє, немає й забарвлення за принципом протитіні. Максимального розвитку воно набуває при гарному освітленні в контрастних умовах, у яких перебувають морські мешканці верхнього шару води, наземні степові форми тощо.
Існують навіть приклади зворотного забарвлення (черево має темніший колір порівняно зі спиною) тих видів, які повернуті саме черевом до сонячних променів. Зворотне забарвлення властиве виключно нільській рибі Synodontis batensoda, яка плаває голічерева, а також деяким видам павуків і гусениць (рис. 8.2), які живляться і відпочивають переважно в такому положенні. Аналогічне забарвлення скунсів, навпаки, виконує демаскуючу функцію, роблячи цю тварину помітною (далі йтиметься про апосематичне (попереджаюче) забарвлення).
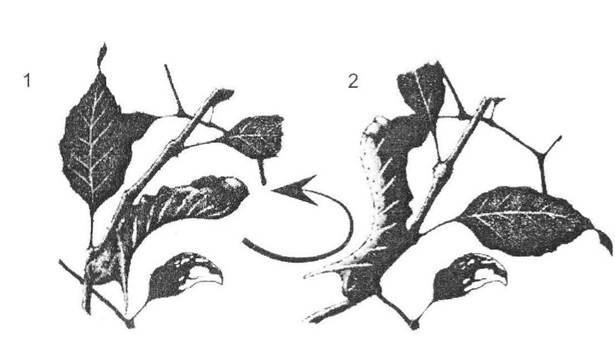
Рис. 8.2. Зворотне забарвлення за принципом протитіні на прикладі Smerinthus ocellatus (рис. О.В. Карнаухова):
1 - нормальне розташування гусені під листком - вона здається плоскою; 2 - при перевертанні гілки забарвлення ще посилюється сонячними променями та гусінь стає особливо помітною
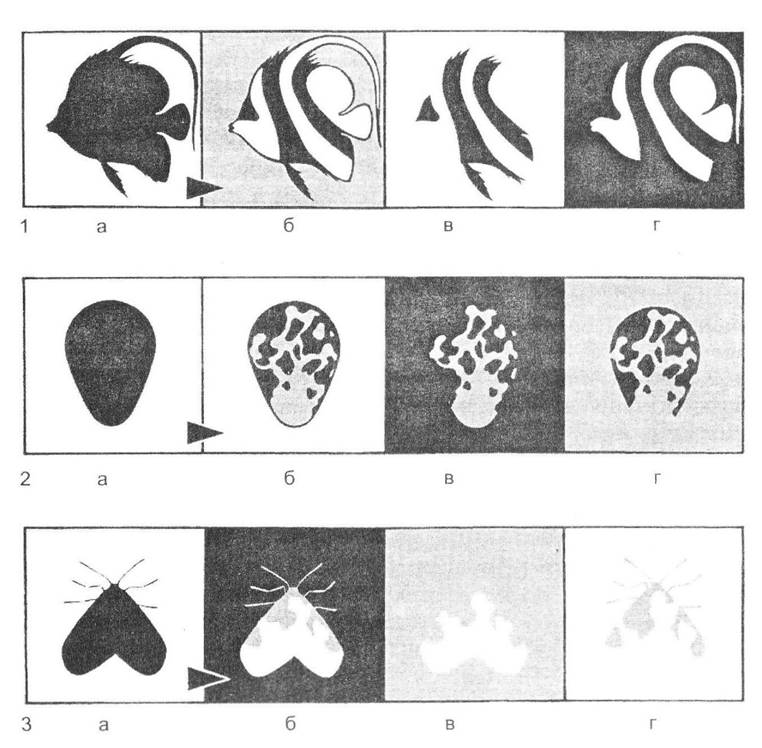
Рис. 8.3. Механізм дії розчленованого забарвлення (рис. О.В. Карнаухова)
Абрис тіла риби (1), яйця (2) або метелика (3) на одноманітному фоні (а) є дуже чітким та легко розпізнається ймовірними ворогами. Якщо організм має розчленоване забарвлення (б), то форма тіла розпізнається гірше. Якщо ж частина забарвлення тіла збігається з тлом фону (в, г) впізнати організм майже неможливо.
Найбільш цікавим типом «камуфляжу» є так зване розчленоване забарвлення.
Використання зазначених типів критичного забарвлення - подібності до тла та скрадаючої протитіні - дозволяє тварині ставати непомітною в умовах одноманітного оточення. Але в природі воно трапляється не так часто. Безперервність поверхні тіла тварини може виказати її місцезнаходження. Уявне порушення цілісності форми досягається спеціальним малюнком - розчленованим забарвленням. Плями або смуги в цьому типі пристосувань можуть бути досить яскравими, але тварина стає непомітною для інших видів. Одним із прикладів подібного пристосування є східноафриканська жаба Rana adspersa, яка забарвлена в буро-зелені кольори, але вздовж спини пролягає яскрава жовта смуга, яка начебто поділяє поверхню навпіл. Смуга ця досить помітна, але не асоціюється з самою жабою і не дозволяє швидко розпізнати організм. Подібним чином розмальовані деякі саранові, змії, що також забезпечує їм маскування тіла. Дієвість подібного пристосування значно зростає в тому випадку, коли деякі компоненти забарвлення збігаються з фоном, а інші проступають досить чітко (рис. 8.3). Тому гама кольорів не повинна відрізнятися від забарвлення середовища, а малюнок не може збігатися з формою тіла, яку він маскує, що досягається поєднанням контрастних кольорів (темний малюнок на світлому фоні). Чим більший контраст між сусідніми елементами малюнка, тим ефективніша дія розчленованого забарвлення.
Існує також чимало прикладів складеного розчленованого забарвлення. Якщо розчленоване забарвлення створює уявлення про розірваність цілісних предметів, то складене розчленоване забарвлення справляє враження безперервності кількох поверхонь. Наслідком останнього різновиду маскування є неможливість розпізнати тіло в цілому або його частину чи орган (рис. 8.4). Так, у деяких тварин маскується око як один з найголовніших органів чуття. Навіть у далеких систематичних групах тварин досягається воно майже однаково - за рахунок поздовжніх (інколи - поперечних) більш темних порівняно з іншим забарвленням смуг, які облямовують око (риби, жаби, змії, ящірки, деякі ссавці і безхребетні).
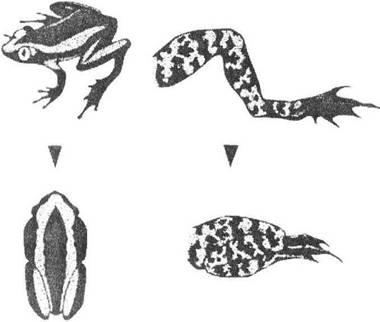
Рис. 8.4. Приклади складеного розчленованого забарвлення, при якому ефект смуг виникає лише в об'єднаних частин тіла
(рис. О.В. Карнаухова)
Розчленований малюнок використовується не лише для маскування поверхні тіла, а й для скрадання обрису. Контур тіла може виказати тварину навіть за умови тотожності її забарвлення довкіллю. Малюнки, що розчленують контур, повинні пересікати останній і закінчуватись на його межі. І, навпаки, якщо малюнок проходить вздовж контуру, то він підкреслює його, завдяки чому тварина стає ще помітнішою (використовується в апосематичному забарвленні).
Максимального захисного значення розчленоване забарвлення набуває в тих видів, загальний малюнок яких наближається до кольорової структури довкілля - місця відпочинку, на якому ця особина сидить переважно нерухомо. Подібне криптичне забарвлення властиве представникам багатьох систематичних груп і залежить від безпосереднього оточення тварини. Але в деяких випадках важливим стає ще й правильна орієнтація організму на відповідному фоні (рис. 8.5), що ще раз підкреслює значення вихідної активності особин.
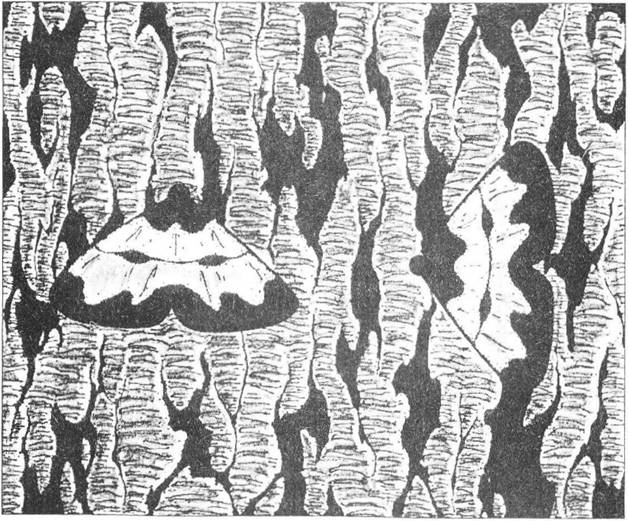
Рис. 8.5. Помітне (1) та непомітне (2) розташування метелика на корі дерева (рис. О.В. Карнаухова)
Наближення малюнка до певного середовища властиве переважно стенобіонтним видам, які обмежені певного кормовою рослиною, ділянкою для розмноження тощо, тобто спеціалізоване криптичне забарвлення трапляється лише серед спеціалізованих видів з дуже вузькою екологічною нішею.
Тварини з криптичним забарвленням стикаються з проблемою наявності тіні: вона може бути більш помітною, ніж сама тварина. В усіх попередніх прикладах пристосувань тінь не усувалася, через що не досягалася й повна непомітність тварин. Оскільки на нерівній поверхні (гущавина трави, кущі, листя) вона майже втрачається, то це було використано мешканцями відкритих просторів - ящірками, птахами, метеликами, які відпочивають серед скель, стовбурів дерев, на голій землі тощо.
Метелики, які відпочивають зі складеними догори крилами, орієнтують тіло відносно сонця, щоб тінь від крил перетворилася на майже непомітну смугу. При цьому тварини розгортають своє тіло відповідно до руху сонця (геліотропізм). Подібні пристосування властиві багатьом видам, зокрема європейським сатирам. В останніх геліотропізм може також виявлятись у нахилі тіла відносно вертикального положення, а в окремих видів — навіть до лежачого стану (Thecla rubi), завдяки чому крила закривають власну тінь. Необхідність таких пристосувань зумовлена тим, що тінь від крил значно помітніша за саму тварину, яка має маскуюче забарвлення.
У багатьох геконів на хвості наявні лопаті (у різних видів різної ширини), які допомагають маскувати тінь хвоста на відносно рівних поверхнях скелі, стіни або стовбура дерева (рис. 8.6). Найбільшого розвитку подібні пристосування набули в лопате-хвостого гекона (Ptychozoon kuhli), який, крім великих хвостових лопатей, має ще й бокові складки шкіри вздовж тіла. Оптичним наслідком таких сплощених форм тіла є також зміна світла й тіні вбік сплощення рельєфу.
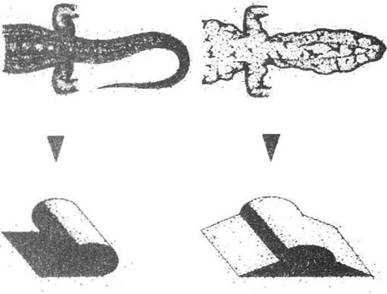
Рис. 8.6. Механізм приховування тіні на сонці (рис. О.В. Карнаухова)
Підтверджує пристосувальне значення критичного забарвлення і його відсутність або слабший прояв у тих форм, які його не потребують. Так, птахи як найбільш активні й рухливі тварини знаходяться в постійному переміщенні в пошуках їжі та інших ресурсів. Наслідком цього стає швидка зміна навколишнього середовища, тому більшість представників цієї групи має переважно неспеціалізовану систему захисного забарвлення. Але ті види, що гніздяться чи відпочивають в умовах відносно відкритих ландшафтів (дрохви, курині, дрімлюги тощо), теж набувають криптичного забарвлення і відповідної поведінки.
Далеко не всі тварини мають захисне забарвлення, але не тому що воно не має пристосувального значення. Подібний «камуфляж» є лише одним із можливих шляхів пристосування до певних умов існування. Оскільки будь-які властивості вимагають певних матеріальних та енергетичних ресурсів, то організм постає перед вибором найбільш прийнятного для себе способу захисту.
Близькою за змістом до криптичного забарвлення є імітація форми, структури та забарвлення довкілля, його компонентів, зокрема неживих або неїстівних об'єктів. Хижаки можуть причаїтися у вигляді байдужих чи навіть привабливих предметів, що значно підвищує результативність їх полювання, наприклад, рибоподібний виріст у морського чорта (Lophius piscatorius).
Багатьом тваринам з віддалених систематичних груп властиве наслідування форми й забарвлення різноманітних листків (рис. 8.7). Як модель можуть використовуватись будь-які листки - різного кольору, форми та стану (мертві й живі, висохлі, покороблені тощо). Але в усіх випадках листок має бути тонким, і враження плаского тіла може досягатися різними засобами.
Найлегше подібний ефект створюється різними видами комах за рахунок плоских поверхонь крил. Найбільш досконалим наслідуванням листка відзначаються представники метеликів індо-малайського роду Kallima, обрис і забарвлення яких разом з уявним черешком, системою середньої й бокових жилок, загостреною вершиною і навіть «пошкодженнями» від хвороб і комах доводять загальний вигляд тварини майже до повної тотожності з листком.
Подібний тип пристосування властивий також багатьом південноамериканським лісовим метеликам (роди Pierella, Аnоеа тощо). У деяких із них (Haetera) крила прозорі, і метелики стають майже непомітними завдяки листкам, які просвічують крізь крила. У нічних метеликів, зокрема п'ядунів, крила розгорнені і нагадують листок лише у випадку складання - складений малюнок (рис. 8.7).
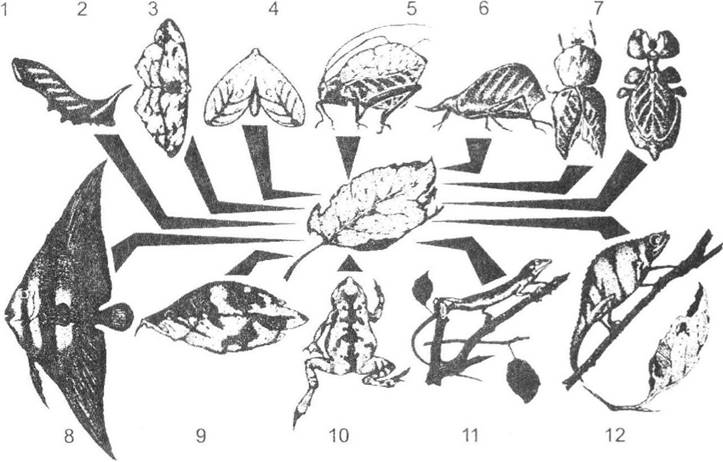
Рис. 8.7. Наслідування ознак листка представниками хребетних та безхребетних тварин (рис. О.В. Карнаухова):
1 - гусениця (Smerinthus ocellatus); 2 - п'ядун (Timandra amata); 3 - метелик (Minioides ornata); 4 - коник (Cycloptera sp.); 5 - представник саранових (Systella rafflesii); 6 - богомол (Choeradodis rhomboidea); 7 - листотілка (Phyllium crurifolium); 8 - риба (Platax vespertilio); 9 - риба (Monocirrus polyacanthus); 10 - ропуха (Bufo typhonius); 11 - ігуана (Polychrus marmoratus); 12 - хамелеон (Rhampholeon boulengeri).
Більш вишукане наслідування пласкості листка досягається сплющенням тіла, що властиве деяким представникам хребетних тварин. Справжнє сплощення обмежене необхідністю відповідної перебудови внутрішніх органів тварини.
Амазонський вид риб Monocirrhus polyacanthus (місцеві жителі називають її «риба-листок») дуже схожий на мертвий листок (рис. 8.7). Подібність така, що впійману рибу складно відрізнити від виловленого листка. Досягається вона завдяки значному латеральному сплющенню тіла, створенню обрису листка, спеціальному забарвленню з імітацією системи жилкування і розвитку «бороди», яка нагадує черешок. До цього підключається ще й поведінковий аспект: зазначений вид живе лише в умовах майже стоячої води і рухається дуже непомітно, наче листя, підхоплене невеликою течією. Оскільки цей представник є хижаком (живиться дрібними рибками), такий комплекс
пристосувальних ознак стає особливо важливим: жертва повинна підпустити його близько до себе, інакше полювання буде невдалим. В інших випадках при наслідуванні структури листка (ропуха Bufo typhonius, комахи р. Phyllium тощо) сплющення тіла відбувається в дорзовентральному напрямку. Враження тонкості може створюватися за принципом протитіні (гусінь бражників, ящірки тощо) або за рахунок бічних складок тіла чи в інший спосіб.
Наслідуватись можуть також кора дерев, лишайники, водорості, каміння, ліани, екскременти тварин та інші предмети, які знаходяться в безпосередньому оточенні тварини. Важлива також відповідна поведінка, без якої імітація втрачає будь-який сенс.
Протилежного значення набуває апосематичне (попереджуюче) забарвлення.
Апосематичне забарвлення, на відміну від наведених пристосувань, переслідує зовсім іншу мету - зробити тварину дуже помітною в тих умовах, в яких вона мешкає. Криптичне забарвлення навіть з використанням яскравих кольорів наближує тварину за загальним виглядом до її оточення. У попереджуючому забарвленні використовується саме така гама кольорів, яка завжди стає помітною для ймовірних ворогів (у першу чергу це червоні, білі, чорні, жовті кольори у вигляді різко окреслених плям або смуг).
Забарвлення тварини, яке виникає під впливом статевого добору (статевий диморфізм), також має на меті привернення уваги. Але малюнки і кольори, які використовуються в цьому випадку, є переважно ніжними, на відміну від яскравих і дуже помітних апосематичних забарвлень.
Апосематичні тварини мають певні відмінності й у поведінці. Якщо організми з криптичним забарвленням переносять піки своєї активності на сутінки (під час відпочинку завмирають або займають відповідне положення, в інший час рухи їх є дуже прудкими), то при попереджуючому забарвленні особини обирають для своєї активності той час доби, коли вони максимально помітні (переважно день). Рухаються вони досить повільно, цілком покладаючись на свої захисні властивості. Це стосується як наземних, так і інших форм (політ сонечка — повільний та незграбний - може бути тому підтвердженням).
Значні відмінності властиві навіть організмам з близьких систематичних груп. Так, більшість пустельних ящірок покладається переважно на свою пильність, прудкість, захисне забарвлення та здатність до закопування. Але отрутозуби підозрілий (Helodorma suspectum) і жахливий (Н. horidum), єдині отруйні представники своєї групи, майже не здатні до втечі, проте мають апосематичне забарвлення (рожеві та жовті кольори на чорному тлі) і є спритними та небезпечними в захисті.
Денний спосіб життя для тварин з апосематичним забарвленням не обов'язковий. Перехід до нічної активності (скунс та деякі інші представники куницевих, великі жуки, наприклад турун Anthia sexguttata) зумовлений тим, що ці тварини полюють на організми з нічним способом життя (переважно комах).
Якщо проаналізувати річну динаміку, то виявляється, що найбільша щільність подібних видів припадає на той проміжок часу, коли чисельність інших тварин є досить високою. Пов'язане це з тим, що молоді тварини під час «навчання», а інколи й дорослі, припускаються помилок, з'їдаючи й представників видів з апосематичним забарвленням. Однак в періоди напруженої боротьби за існування, зокрема через нестачу їжі, хижаки можуть переходити з улюбленої на будь-яку іншу, у тому числі й несмачну чи навіть неїстівну. Тому найбільшого ефекту апосематичне забарвлення надає тварині за умови наявності в хижаків великого вибору їстівних тварин.
Крім попереджуючих якостей, апосематичні тварини повинні мати й певні захисні властивості. Це стосується будь-яких засобів захисту, які роблять тварину непривабливою чи неприємною на смак для потенційних ворогів.
Отрута є одним із найдієвіших та спеціалізованих засобів захисту від ворогів. Токсичні властивості притаманні багатьом тваринам, але отруйність є відносною, оскільки залежить від біохімічних властивостей отрути та чутливості до неї окремих організмів. Іншими словами, тварини, що є отруйними для одних видів, виявляються їстівними для інших, і, навпаки, їстівні організми можуть виявити отруйні властивості стосовно тварин. Це видно на прикладах циклів незахищеності отруйних тварин Є.Н. Павловського (рис. 8.8).
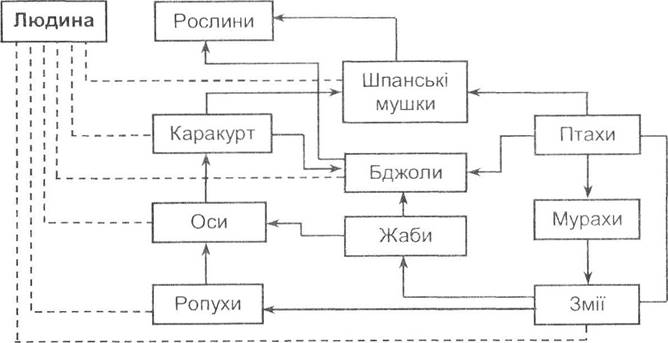
Рис. 8.8. Цикли незахищеності від отруйних організмів (за Є.Н. Павловським, 1961)
Шпанська мушка (Lytta vesicatoria) містить кантаридин у гемолімфі і є отруйною для людини, але з'їдається каракуртом (Latrodectus tredecimguttatus). Останній, незважаючи на дуже загрозливу для людини отруйність, може стати здобиччю деяких видів ос. Личинки ос і бджіл (навіть за наявності добрих захисних властивостей імаго, суспільного способу життя і піклування про нащадків) виявляються незахищеними від земноводних - жаб та ропух. Амфібії мають отруйні залози в шкірних покривах, але дія їх не поширюється на змій. Змії, у свою чергу, можуть стати здобиччю мурах або інших тварин, зокрема свиней. Мурах, бджіл та шпанських мушок безпроблемно вживають птахи.
Незважаючи на певну обмеженість практичної дії отрути, її наявність сама по собі збільшує ймовірність виживання відповідних тварин, оскільки звужує коло потенційних ворогів. Існують такі види тварин, у яких розвинені справжні отруйні залози зі спеціально модифікованими колючками тощо, які використовуються як зброя для нанесення уколу й отруєння рани. До них можна віднести окремих представників риб, зокрема скатів (Trygon, Aetobatis), велику кількість змій, перетинчастокрилих тощо. Інший тип являють тварини, саме тіло яких містить отруту. Це риби-скалозуби (Tetrodontidae - рис. 8.9), амфібії (Salamandra maculosa, Bombinator igneus, Bufo marinus тощо).

Рис. 8.9. Скалозуб (Tetrodon fluviatilis) в нормальному та роздутому стані(рис. О.В. Карнаухова)
Ефективним засобом захисту можна вважати подразливі або відштовхуючі секрети. Він пов'язаний з активним виділенням твариною відштовхуючих речовин під час загрози їй з боку ймовірних ворогів: завдяки наявності спеціалізованих пристосувань виприскує їдкий секрет безпосередньо на ворога, зупиняючи його. Так роблять жуки-бомбардири (Brachinus), змія, що плюється (Sepedon hae mac hat es), та скунс (Merhitis merhitis).
У більш простих випадках тварина виділяє захисний секрет, запах або смак якого позбавляє нападника бажання поживитись цим об'єктом. Такою здатністю наділені передусім комахи: велика кількість клопів, деякі жуки, зокрема туруни, сонечка, шпанські мушки та ін.
Близьким за значенням та ефективністю є неприємний за пах або смак тканин чи рідини тіла тварин, тобто пасивна неїстівність певних тварин робить їх відносно захищеними від імовірних хижаків.
Деякі неотруйні види неїстівних властивостей набувають унаслідок накопичення відповідних речовин під час живлення. Так, гусінь данаїд живиться на ваточниках, що містять серцеві глікозиди - отруйні для птахів і ссавців. Ці речовини не лише не отруйні для комахи, а навіть накопичуються в її організмі і передаються імаго, роблячи останніх неїстівними. Якщо гусінь поїдатиме капусту чи ваточники, які не містять глікозидів, метелики стануть їстівними.
Способи демонстрації неїстівної сутності в різних апосематичних тварин можуть бути різними.
Крім попереджуючої дії постійних пігментів та малюнків, у певних обставинах можуть використовуватись і тимчасові пристосування, завдяки яким непомітна в звичайному стані тварина відразу набуває апосематичних властивостей.
Відносно простим способом використання попереджувальних властивостей є демонстрація шляхом збільшення розмірів тіла. Така здатність розвинулась у представників систематично далеких груп: риб, амфібій, рептилій тощо. Збільшення розмірів може бути удаваним, як у ссавців чи птахів унаслідок здиблювання
волосся чи пір'я відповідно. При цьому деякі форми справді збільшують розміри тіла шляхом розтягування легенів або травного тракту повітрям чи навіть водою. Інколи до цього додається ще й специфічна поза відносно ворога, яка посилює загальний ефект.
Прикладом справжнього збільшення розмірів тіла можуть бути такі риби, як скалозуби (Tetrodontidae) та їжаки-риби (Dioclontidae). Перші з них захищені надзвичайною отруйністю тіла, а другі мають майже суцільний шипуватий панцир. У випадку роздратування ці тварини роздуваються до кулеподібної форми (рис. 8.9). Таке пристосування властиве й деяким рептиліям (СhamaeLeontidae, Dispholidus typus тощо) та амфібіям (Bufo marinus, Callula pulchra та ін.).
Інколи аналогічний ефект досягається шляхом об'єднання великої кількості дрібних організмів, наприклад дрібних їжа- ків-риб. Якщо якась особина відривається від скупчення, її відразу з'їдає хижак.
Досить часто з метою попередження тварини використовують місцеве роздування чи піднімають спеціальні утворення - гриви, капюшони, зоби тощо. Звичайно ці утворення укладаються на одній площині і спрямовуються вбік нападника. Так поєднуються демонстрація певних структур з відповідною поведінкою, як у плащоносної ящірки (Chlamidosaurus kingi), роздуття шиї у деяких змій (Naja nigricollis, Thelothornis kirtlandii тощо).
Несподіване виставлення яскравого забарвлення — одна із попереджуючих демонстрацій, яка інколи поєднується з попереднім типом тимчасових пристосувань. Як бачимо, в одному організмі можуть поєднуватись як криптичне, так і попереджуюче забарвлення.
Деякі тварини з яскравим забарвленням нижніх частин тіла здатні перекидатися під час загрози на спину, демонструючи апосематичні властивості черева з метою відлякування ворога. Ці яскраво забарвлені поверхні у звичайних умовах приховані, а поза погрози приймається лише за особливих обставин. Поза ж, яку займає тварина, максимально демонструє апосематичні ділянки ворогові.
Прикладом може бути кумка червоночерева (Bombina bombina), яка перекидається на спину, виставляючи яскраві плями на череві. Інколи тварина дещо піднімається над субстратом, демонструючи ворогу частину тіла з яскравими тонами, що властиве зміям (кобри, деревні змії тощо), деяким ссавцям (перегузня стає на задні лапи, демонструючи біле черево) і багатьом членистоногим. У метеликів демонстрація яскравих ділянок тіла відбувається переважно за рахунок рухів крил, які в стані спокою приховують яскраве забарвлення черевця або нижніх крил.
Механізм попередження може полягати в розтягненні ділянок шкіри, в яких приховані яскраві апосематичні ознаки. Так, у сіамської ропухи (Caltula pulchra) при роздуванні тіла шкіра розтягується і на спині з'являються дві широкі смуги жовтого кольору. Подібне пристосування властиве також ящірці Liolepis bellina, яка має буре криптичне забарвлення. При загрозі тварина сплющує своє тіло і виставляє бічні пурпурні смуги з жовтими включеннями.
Складніший та більш спеціалізований механізм полягає в розгортанні віялоподібних структур, складки яких приховують попереджуючі ознаки. У морських півнів (Trigla kumu, Leptotrigla burgeri) роль віяла відіграють грудні плавці, які своїми розмірами та забарвленням попереджають про наявність отруйних шипів. Максимального розвитку цей тип пристосувань набув у плащоносної ящірки (Amblyrhynchus cristatus), про яку ми вже згадували раніше. Подібним пристосуванням можна вважати чубчик одуда (Upupa epops).
У багатьох випадках апосематичне забарвлення супроводжується й відповідною попереджуючою поведінкою. Це може стосуватись як рухів (розмахування частинами тіла, кидки тощо), так і різноманітних звуків, які ніколи не бувають занадто гучними. До цих досить відомих пристосувань можуть додаватись ще й такі, як попереджуючий запах (дикобрази, скунси).
Важливою властивістю апосематичних тварин можна вважати їх підвищену живучість. Представники цих видів є дуже витривалими і життєздатними, що пояснюється необхідністю переносити пошкодження. Помітна зовнішність може провокувати напади молодих недосвідчених тварин, після чого апосема- тична тварина залишається без важких пошкоджень, які могли б виявитись смертельними для інших видів.
Одним із найцікавіших пристосувань, пов'язаних із забарвленням тіла, є мімікрія.
Часто мімікрію розглядають широко - як наслідування забарвлення чи форми певних тварин або органів рослин. Але ми використовуватимемо даний термін у вузькому розумінні - як наслідування властивостей апосематичних тварин. Одразу зазначимо, що мімікрія як пристосування відіграє таку саму захисну роль, як і криптичне забарвлення. Тому результат обох
.типів пристосувань є близьким - підвищення виживання організмів. Відмінність полягає лише в механізмі спрацювання. При криптичному забарвленні організм наслідує властивості предметів, які байдужі для ворога. У випадку мімікрії тварина стає помітною, наслідуючи особливості предметів, які ворог знає і яких уникає. Фахівці розрізняють два види мімікрії — бейтсівську та мюллерівську.
Під бейтсівською мімікрією розуміють випадки, коли незахищений їстівний вид наслідує властивості неїстівного або добре захищеного виду. Перший з них є імітатором, а другий - моделлю. Інакше кажучи, схожість організмів вводить імовірних ворогів в оману, хибно виконуючи функцію попереджуючого (псевдоапосематичного) забарвлення (рис. 8.10).

Рис. 8.10. Приклад бейтсівської мімікрії в африканського метелика (Papilio dardanus) (за М.С. Гіляровим, 1984):
1 - самка форми Hippocoon; 2 - її модель (Amauris niavius); 3 - самка форми сепса; 4 - її модель (Amauris echeria)·, 5 - самець цього виду не має міметичних властивостей
Одне з головних правил такого пристосування є те, що імітаторів завжди менше, ніж моделей. В іншому разі вороги швидко розпізнавали б обман, що послабило б навчальне значення нападу хижака на апосематичну тварину.
Міметичні пристосування призводять до значних змін в організації видів, відрізняючи їх від споріднених. Відбуваються відхилення від типової зовнішності групи, натомість наслідуються чужі ознаки, що набуває фундаментального значення. Так, окремі види ктирів нагадують бджіл з роду Хуіосора і зовнішньо дуже відрізняються від родичів, бо мають широке волохате черевце та пігментовані крила.
Проте зміни не обмежуються лише зовнішнім виглядом тварин — суттєво змінюється й поведінка останніх. Це може стосуватись часу активності, як у метелика склівки шершнеподібної (Aegeria crabroniformis), який на відміну від більшості представників метеликів групи Heteroptera веде денний спосіб життя, як і його модель. Схожими також можуть бути особливості рухів тварин і навіть спеціалізовані поведінкові акти. Спійманий жук-вусач Dyrphya, який за зовнішнім виглядом нагадує наїзника, згинає кінчик черевця, наче намагається ужалити, висовуючи при цьому білий стрижень, яким рухає наче жалом.
Мюлерівська мімікрія - це набуття кількома видами з апосематичними властивостями однакового вигляду, завдяки чому вони краще розпізнаються хижаками. Цей тип пристосувань прискорює навчання ворога щодо неїстівності певної групи апосематичних тварин завдяки тотожності їх попереджуючого забарвлення. Інколи це явище називають ще синапосематичним, або спільним попереджуючим, забарвленням. При мюлерівсь- кій мімікрії схожість видів може бути й незначною, але тварини мають спільні властивості (рис. 8.11).
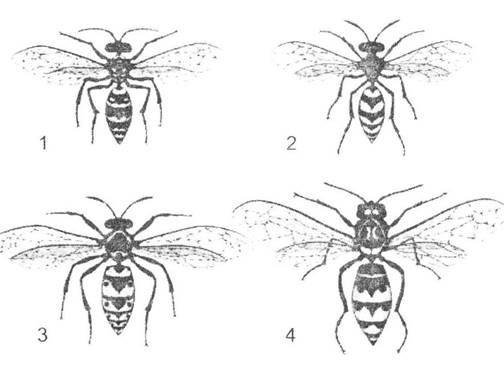
Рис. 8.11. Явище мюлерівської мімікрії на прикладі різних видів ос (рис. О.В. Карнаухова):
1 - гальська оса (Polistes gallicas); 2 - бджолиний вовк (Philanthus triangulus); 3 - германська оса (Paravespula germanica); 4 - шершень звичайний (Vespa crabro)
У природі розрізнити ці два види мімікрії досить складно через відносність поняття «неїстівність» та наявність спільних рис. Форми, пов'язані між собою міметичною схожістю, мешкають в одній місцевості, оскільки ефективною мімікрія є лише за умови спільного існування зазначених форм і їх спільного
потерпання від нападів ворогів. Близькою стає й поведінка цих видів, про що йшлося раніше.
Закономірність мімікрії полягає в тому, що схожість тварин поширюється лише на зовнішні, помітні для стороннього ока структури. Причому шляхи досягнення схожості можуть бути різними і не залежать від особливостей анатомо-морфологічної будови тварини.