ТЕОРЕТИЧНІ ОСНОВИ РИБНИЦТВА - І.М. Шерман - 2011
2.ТЕОРЕТИЧНІ ОСНОВИ ФОРМУВАННЯ ТА ВИКОРИСТАННЯ ПРОДУКТИВНИХ ВЛАСТИВОСТЕЙ РИБ
2.6. Теоретичні аспекти живлення і годівлі риб
Інтенсивне виробництво риби потребує застосування кормів, а сама інтенсифікація супроводжується збільшенням особин на одиниці площі або об’єму води, що за аналогією з тваринництвом, можна розглядати, як стійлову форму вирощування сільськогосподарських тварин, яка передбачає ефективне використання кормів штучного походження.
За цих умов природні корми, тобто рослинна і тваринна їжа, їх рештки, які є основою раціону певних видів риб у межах ареалу, практично відсутні або їх кількість незначна і не може істотно впливати на живлення риби. Ще складніша ситуація створюється у разі використання теплих за походженням і термальних вод використання яких задовольняє бажання оптимізувати термічний режим з метою подовження періоду ефективної вегетації за рахунок закономірної інтенсифікації фізіологічних процесів, що реально для пойкілотермних тварин. Фізіологічні процеси, у свою чергу, вимагають відповідного енергетичного забезпечення, що потребує додаткових витрат кормів і має одночасно гарантувати ефективне збільшення іхтіомаси видів, які культивують.
Сьогодні левова частка собівартості продукції рибництва, яку вирощують за інтенсивними технологіями, припадає на корми, тому важко переоцінити актуальність проблеми, її теоретичної та практичної складових, в умовах сучасного ставового та індустріального рибництва.
Годівля є найбільш дієвим і ефективним заходом інтенсифікації рибництва в спеціальних рибничих ставах індустріального типу, малих і середніх водосховищах, забезпечує отримання максимальних показників рибопродуктивності за умов забезпечення визначальних абіотичних параметрів середовища.
Розглядаючи підняту проблему, шукаючи шляхи її вирішення, необхідно володіти певною інформацією відносно основних закономірностей живлення та годівлі різних видів риб.
Виходячи з існуючих сучасних уявлень про живлення риб, необхідно наголосити на тому, що живлення є провідним фактором, який визначає існування виду в ареалі. Поряд з цим загальновідомо - жоден організм не може збільшити свою особисту масу на більший показник ніж масу сухої речовини, яку мають спожити їм кормові компоненти раціону. При цьому ступінь засвоєння різних компонентів раціону штучного і природного походження має здатність демонструвати різний рівень засвоєння. Показник ефективності живлення представлений двома складовими, а саме це кормовий коефіцієнт, який характеризує кількість одиниць необхідного корму спожитого для отримання одиниці приросту особини. Друга складова залежить від виду риби, її біологічних особливостей, здатності акумулювати і трансформувати корми в особисту біомасу. З віком і ростом риби кормовий коефіцієнт зростає на фоні ідентичного раціону за поживним складом, що необхідно враховувати при годівлі риб. Невідповідність якості харчів вимогам риби кормовий коефіцієнт зростає, що призводить до загальної перевитрати кормів, аналогічна ситуація спостерігається при живленні і годівлі риби в умовах порушення відповідних параметрів фізико - хімічного стану середовища.
Термін кормовий коефіцієнт є придатним виключно для загальної оцінки якості кормів, а в умовах аквакультури виключного значення набуває критерій, якій характеризує фактичну витрату кормів на отримання одиниці продукції культивуємих видів риб, яким є витрата корму на одиницю продукції.
Повертаючись до теоретичних аспектів годівлі риб наголошуємо на характері живлення риб, який сформувався протягом онтогенезу, що необхідно враховувати при формуванні якісних і кількісних параметрів раціону у часі і просторі.
Повертаючись до філогенезу теорії живлення риб, доцільно наголосити на тому, що первинними продуцентами органічної речовини є хемосинтезуючі бактерії, але левова частка продуцентів сучасних гідроекосистем представлена фітопланктоном та макролітами. Флора є їжею багатьох безхребетних, а також окремих видів риб. Безхребетні, в свою чергу, виступають в якості кормових гідробіонтів для багатьох видів риб, які віднесені до мирних твариноїдних риб. Хижаки, у свою чергу споживають переважно мирних рослиноїдних і твариноїдних риб. Складні харчові взаємовідносини формуються у трофічні ланцюги, які характеризуються відповідними якісними і кількісними параметрами.
Для багатьох видів риб типовим є зміна кормових об’єктів протягом онтогенезу. Для раннього постембріогенезу характерним є використання в якості корму дрібних кормових об’єктів низького трофічного рівня, з віком здійснюється перехід на крупні кормові об’єкти високого трофічного рівня. Виходячи з викладеного вище, можливість певного забезпечення і його сенс полягає в тому, що в процесі онтогенезу внутрішньопопуляційна харчова конкуренція поступово нарощується і є мінімальною на ранніх етапах онтогенезу.
Розглядаючи ситуацію, яка є типовою для природних акваторій, доцільно врахувати, що чисельність особин з віком падає і тут передує природна смертність у поєднанні з інтенсивністю процесу, на цьому фоні біотичних і антропогенних факторів харчова конкуренція знижується.
Частина популяції може гинути протягом раннього онтогенезу в період переходу з жовткового живлення на зовнішній корм. При цьому суттєве значення набуває забезпеченість вільних ембріонів або передличинок жовтковою їжею, а це у свою чергу залежить від умов нагулу батьківського матеріалу у попередній сезон.
В межах фауністичних комплексів напруженість харчових взаємовідносин, споживаючих аналогічні корми послаблена, що обумовлено високим рівнем спеціалізації живлення різними видами організмів. За різких змін умов види, які утворюють певний комплекс, здатні переходити на споживання нетипових для виду об’єктів, що може приводити до загострення харчових взаємовідносин з видами - конкурентами конкретного комплексу.
Кожний вид в процесі тривалого філогенезу напрацював необхідні передумови для ефективної трансформації кормового ресурсу, а точніше його частину у кормову базу, що фактично узгоджено з віковими змінами та статевими змінами. Відносно здатності живлення особин багатьох видів у період раннього постембріогенезу сходною їжею необхідно наголосити, що формально це явище може розглядатися в якості конкуренції між певними видами, але вона фактично відсутня.
Запропонована концепція фактичної відсутності харчової конкуренції пов’язана з тим, що особини різних видів демонструють розтягненість нересту в часі і просторі, а це означає, що їх ембріогенез відбувається далеко не одночасно. Виходячи з цього, періоди раннього постембріогенезу, коли живлення формально однотипне, не співпадають у часі, а це забезпечує фактичну відсутність конкуренції в період проходження однакових стадій і етапів розвитку різних видів риб.
Така біологічна особливість різних видів риб є основою для використання їх видоспецифічних рис живлення в умовах аквакультури, яка орієнтована на культивування різних видів риб за принципом полікультури. Поряд із цим знання розглянутих особливостей живлення дозволяють ефективно корегувати технологію полікультури в умовах різних широт, в індустріальному рибництві.
Підсумовуючи викладене, необхідно наголосити на тому, що в природних умовах об’єктивно існує внутрішньовидова та міжвидова конкуренція, яка пом’якшується біологічними особливостями іхтіофауни певних акваторій і широт. Об’єктивний характер процесів живлення в природних умовах дає в руки фахівця вагомий важіль впливу на формування технології годівлі певних видів і вікових груп з урахуванням індивідуальних умов рибничих підприємств різної спеціалізації.
Світова іхтіофауна за характером живлення умовно поділена на еврі - і стенофагів, що має виключне значення для становлення природних іхтіоценозів і одночасно орієнтує науковців і практиків на шляхи створення високопродуктивних штучних іхтіоценозів. Виходячи з цього, розглядаючи біологічне значення явища еврифагії і стенофагії, доцільно певну увагу приділити загальним уявленням відносно живлення риб.
Виходячи з характеру живлення певних видів риб, у складі світової іхтіофауни достатньо умовно розрізняють відповідні групи, які представлені рослиноїдними, твариноїдними і хижими видами.
До рослиноїдних віднесені види, основу живлення яких складають зелені рослини, представлені фітопланктоном і макрофітами. Вони отримали відповідні назви, а саме фітопланктофаги і макрофітофаги. Твариноїдні риби живляться безхребетними, які не здатні до активного руху, малорухомі, населяють товщу води - зоопланктон і в ґрунті - зообентос. Відповідно до характеру живлення у спеціальній термінології вони сприймаються в якості зоопланктофагів і зообентофагів. Хижаки переважно живляться рибою, але можуть споживати поряд із рибою таких хребетних як жаби, птахи, плазуни, ссавці.
Доцільно наголосити на тому, що розглянутий розподіл має умовний характер. За певних обставин мирні твариноїдні риби споживають молодь різних видів риб, не виключений канібалізм. Багато хижаків здатні споживати зоопланктон та зообентос, а багато видів демонструють змішане живлення. Для певних видів риб існує здатність змінювати характер живлення протягом доби, сезону, року, існують і зміни широтного характеру.
На цьому фоні певну цікавість викликають такі екологічні групи, як еврибіонти та стенобіонти. Еврибіонти населяють переважно прісні, малі за площею та об’ємами водойми для яких типовим є часта і достатньо різка зміна фізико - хімічних параметрів середовища, що безпосередньо впливає на біологічну складову загальної гідроекосистеми. Переважна більшість аборигенної флори і фауни досить легко переносять такі явища, швидко адаптуються і комфортно почувають себе на тлі динамічних змін абіотичних параметрів середовища. Стенобіонти переважно населяють великі за площею і об’ємом акваторії, в яких відсутня стрибкоподібна зміна динаміки абіотичних параметрів середовища, представлених головним чином морями та океанами.
Виходячи з викладеного, наочно, що серед еврибіонтів переважна більшість видів, завдячуючи наявності відповідних механізмів адаптації, здатна не тільки витримувати зміну абіотичних параметрів середовища, але і ефективно на цьому тлі споживати кормові ресурси, трансформувати їх у кормову базу відповідно до видового складу, чисельності і біомаси гідробіонтів. Для стенобіонтів така ситуація може мати критичний характер - зміна фізико-хімічних умов існування може негативно вплинути на видовий склад, чисельність і біомасу кормових гідробіонтів, що відповідно викликає напругу у забезпеченні біологічних потреб організму, а це може поєднуватися з фізіологічною складовою живлення і засвоєння кормових гідробіонтів.
Працюючи в технологічному режимі аквакультури, і, культивуючи еврибіонтів і стенобіонтів, безумовно необхідно враховувати біологічні
особливості екологічних груп риб, які розглядаються, і створювати відповідні умови в процесі годівлі.
Для риб, на фоні їх приналежності до певних екологічних груп, є типовою демонстрація деякої вибіркової здатності в процесі живлення. Достатньо умовно, а це значною мірою залежить від умов, в яких опинилися певні групи риб. Виходячи з цього, можливо розрізнити їжу по відношенню до неї риби на улюблену, замінюючу і вимушену. За наявності оптимальних умов риба надає перевагу улюбленій їжі, яка домінує у кишківнику і шлунку. За умов, коли улюблена їжа відсутня, її мало, формуються умови, коли вона стає недоступною, риба переходить на споживання їжі що замінює, а за умов відсутності останньої риба переходить до споживання вимушеної їжі.
Біологічні особливості риб демонструвати різні риси своєї екології та етології формують здатність до вживання за відносно тривалої відсутності відповідних харчів і змін спектру живлення відповідно до наявності відповідних кормів, не виключене тривале голодування.
Протягом життя різні види риб здатні демонструвати певні особливості живлення, що має пристосувальний характер, несе у собі вікові і статеві складові, що робить актуальним використання розглянутих особливостей у рибництві.
Абсолютна більшість представників світової іхтіофауни демонструє різну анатомічну будову органів і систем, які безпосередньо пов’язані з процесами живлення. Дивергенція будови тіла, систем окремих органів є результатом тривалого філогенезу, який забезпечив різноманітність видового складу іхтіофауни, завдячуючи удосконаленню адаптаційних можливостей різних видів риб на фоні достатньо специфічних факторів оточуючого середовища.
Специфічна анатомічна будова різних видів риб демонструє свої характерні риси з віком, а на ранніх стадіях і етапах постембріонального розвитку конвергенція мінімальна, фізіолого - біохімічні процеси з якісного боку мають багато спільного , що орієнтує переважну більшість
компонентів природного іхтіоценозу на споживання одних і тих же харчових гідробіонтів.
Керуючись формальним підходом до запропонованої концепції, виникає думка про те, як це можливо для всіх компонентів іхтіоценозу споживати спільно однотипні харчові гідробіонти, якщо кормових організмів на всіх вистачає. Справа в тому, що мова йде про зоопланктон, який є основою природного раціону практично всіх видів риб в період раннього постембріогенезу. Саме тому сумніви відносно можливості забезпечення зоопланктоном всіх мешканців певної акваторії достатньо проблематично.
В процесі тривалого філогенезу сформовані різні екологічні групи риб, які здатні існувати в умовах однієї акваторії. Однією складовою цього процесу є об’єктивна реальність - існування у високих і середніх широтах видів риб з весняно-літнім і осінньо-зимовим нерестом і відповідно різною тривалістю ембріогенезу. Така особливість суттєво знижує навантаження на зоопланктон, його дрібні, середні, крупні форми, забезпечуючи розходження спектрів живлення не формально, а в часі і просторі.
Поряд з тим в межах кожної екологічної групи розглянутої вище за наявності практично всіх рівних факторів існує достатньо жорстка диференціація відносно нерестових температур, а це забезпечує позитивне розходження в часі нересту різних видів риб. При цьому процес, що розглядається, включає в себе такий фактор, як різна тривалість ембріогенезу. Викладене об'єктивно свідчить про те, що ідентичні стадії і етапи раннього постембріогенезу для переважної більшості риб відповідного іхтіоценозу протікають з великим розривом у часі і просторі, що значною мірою виключає або суттєво знижує фактичну міжвидову конкуренцію в живленні. Внутрішньовидова харчова конкуренція суттєва, але вона згладжується, завдячуючи розтягнутості в діапазоні нерестових температур для особин одного виду, але різних вікових груп, які можуть демонструвати різний фізіологічний стан системи відтворення.
Викладене вище свідчить про те, що конкуренція між різними видами існує, але вона суттєво пом’якшена за рахунок біологічних особливостей видів і екологічних груп в період раннього постембріогенезу.
Поступово, з віком всі види риб переходять на харчовий раціон, типовий для конкретного виду, що є основою віднесення певного виду за характером живлення до фітопланктофагів, макрофітофагів, які безпосередньо споживають флору, яка розглядається в якості головного продуцента первинної органічної речовини. Поряд з рослиноїдними видами риб, яких не багато, існують твариноїдні види риб, які живляться зоопланктоном, зообентосом, детритом. Як було акцентовано увагу вище, вони поєднані в групи мирних твариноїдних і представлені зоопланктофагами, зообентофагами, детритофагами. Окрему групу складають хижаки - основу їхнього раціону складають хребетні, це переважно риба, птахи, жаби, плазуни, ссавці.
При культивуванні різних видів риб доцільно звертати увагу не тільки на видові особливості живлення, а ї на статеві складові процесу живлення, що пов’язано з анатомічною будовою відповідних органів і систем фізіологічної відповідності.
Виходячи з викладеного, вважаємо за доцільне запропонувати достатньо обмежену інформацію з анатомії і фізіології живлення риб у зв'язку безпосередньо з рибництвом.
Анатомічні особливості живлення риб представлені специфічною системою органів живлення і травлення і складаються з ротового отвору, ротової порожнини, глотки, стравоходу, шлунка та кишківника. Ефективне травлення у риб забезпечують великі секреторні залози — печінка і підшлункова залоза. Проте умови існування і харчові потреби риб змусили їх адаптуватись до ширшого харчового спектра, що спричинило значне різноманіття будови та функціонування травної системи. Споживання кормових об'єктів, які знаходяться лише у водному середовищі, сприяло виникненню індивідуальних пристосувань, відсутніх у наземних тварин.
Відповідно до характеру живлення анатомічні та фізіологічні складові мають характерні ознаки і характеристики, які пов’язані з процесами споживання і засвоєння поживних речовин різних кормових гідробіонтів. Більшість фітофагів поїдає обмежене число рослин, має відповідні спеціальні структури для подрібнення їжі, призначені для вилучення максимальної кількості поживних речовин з цього низькоенергетичного виду корму. Необхідно наголосити на тому, що досить умовно до рослиноїдних риб зараховують детритофагів, які споживають детритні маси, що складаються з решток відмерлих гідробіонтів рослинного і тваринного походження, сапротрофних мікроорганізмів та мінеральних часточок, що дозволяє віднести детритофагів до риб, яким притаманне змішане живлення.
У харчовому спектрі всеїдних риб трапляються організми як тваринного, так і рослинного походження, їх наявність у раціоні залежить від доступності окремих кормових організмів, пори року і фізіологічного стану риби.
Споживання кормових компонентів різної енергетичної цінності, які містили неоднакову кількість баластних речовин, зумовило появу певних відмінностей у будові органів травлення рослинно-, тваринно- і всеїдних риб. Насамперед це відбилося на довжині травного каналу відносно довжини тіла риби, що добре ілюструють дані табл. 2.1. Крім харчового раціону, на довжину травного каналу риб впливають інтенсивність живлення, вікові та сезонні зміни, доступність кормових організмів.
Таблиця 2.1. Відносна довжина травного каналу риб різного типу живлення
Вид риби |
Тип живлення |
Відносна довжина травного |
|
Середня |
Коливання |
||
Щука |
Іхтіофаги |
1,0 |
0,80-1,20 |
Судак |
— |
0,8 |
0,70-0,85 |
Окунь |
— |
1,1 |
0,95-1,15 |
Жерех (білизна) |
— |
0,95 |
0,90-1,15 |
Жовтощок |
— |
0,6 |
0,55-0,70 |
Пічкур |
Зоофітофаги |
0,8 |
0,75-0,90 |
Карась |
— |
2,0 |
1,90-2,20 |
Короп |
— |
2,7 |
2,60-3,00 |
Лящ |
Зообентофаги |
1,2 |
1,10-1,25 |
Тараня |
— |
1,3 |
1,10-1,35 |
Білий амур |
Макрофітофаги |
3,1 |
2,50-3,80 |
Білий товстолобик |
Фітопланктофаги |
11,5 |
8,50-13,00 |
Піленгас |
Детритофаги |
4,5 |
4,45-4,60 |
З харчовим раціоном тісно пов'язана будова ротового апарату риб. Форма, розміри і положення ротового отвору, будова зубів і зяберних дуг визначаються типом живлення, розмірами їх кормових об'єктів та умовами водного середовища. Розрізнюють три основні типи положення рота (рис. 2.8): верхній рот — нижня щелепа більша за верхню і ротовий отвір спрямований догори — мають риби, які беруть поживу з верхніх горизонтів; кінцевий рот — коли обидві щелепи однакові — мають риби, які беруть поживу з товщі води; нижній рот — коли верхня щелепа більша за нижню і ротовий отвір спрямований донизу — мають риби бентофаги.
Залежно від розмірів кормових об'єктів, щільності їх розподілу та способу захоплення корму рибою формуються розміри ротового отвору і ротовий апарат. За будовою і функціями розрізняють кілька типів рота риб: хапальний — кінцевий або верхній, великий з гострими зубами як на щелепах, так часто і на лемеші та піднебінних кістках, зяброві тичинки короткі, рідкі і гострі; всмоктувальний — нижній, у вигляді трубки, часто висувний (рис. 2.9), як правило, без зубів, слугує для живлення донними безхребетними; подрібнювальний — кінцевий, з міцними зубами у вигляді пластин або шипів, слугує для подрібнення твердих панцирів безхребетних; планктоноїдний — кінцевий або верхній, зазвичай великий і, як правило, невисувний, зуби дрібні або частіше їх зовсім немає, зяброві тичинки довгі, діють як сито; перифтоноїдний — розміщений у нижній частині голови, у вигляді поперечної щілини, нижня губа має гострий ріжучий край, іноді вкритий роговим чохликом, зубів, як правило, немає.
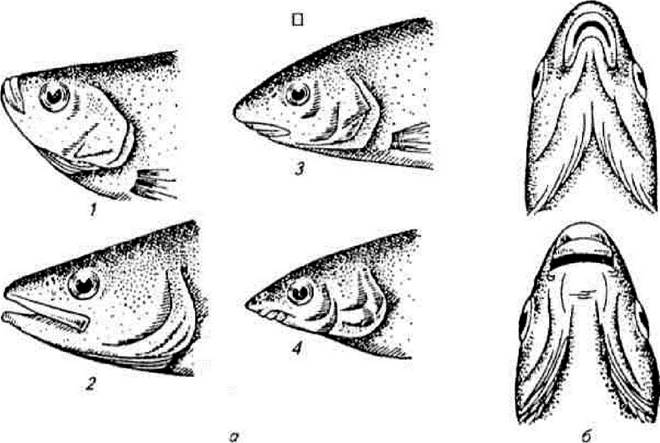
Рис. 2.8. Типи положення ротового отвору:
1 — верхній; 2 — кінцевий; 3,4 — нижній (а — вигляд збоку, б — вигляд знизу)
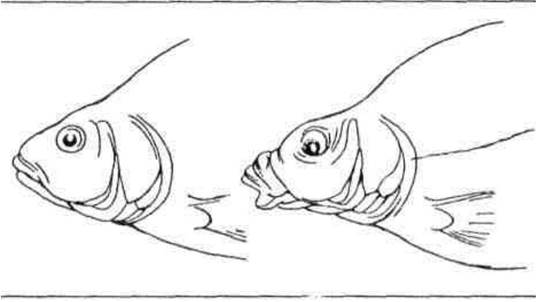
Рис. 2.9. Всмоктувальний (висувний) рот
Ротова порожнина, яку вважають переднім відділом травного каналу риб, переходить у глотку. Це різною мірою виділений м'язовий канал, у передню частину якого крізь відповідні отвори проникають зяброві тичинки і залежно від спектра живлення утворюють цідильний або проштовхувальний апарат різної будови. У задньому відділі глотки багатьох костистих риб є глоткові зуби, які забезпечують механічну обробку, подрібнення корму і попереднє формування кормової грудки. Глотка сполучає ротову порожнину із стравоходом, який має вигляд короткого широкого проходу з кільцем поперечнопосмугованих м'язів і забезпечує проштовхування їжі до шлунка або безпосередньо в передній відділ кишечнику безшлункових риб. У стравоході здійснюються додаткова смакова рецепція за допомогою смакових цибулин і змочування кормової грудки секретом слизових залоз.
У шлункових риб їжа із стравоходу потрапляє до шлунка, який має вигляд розширеної травної трубки, де відбувається механічна і хімічна обробка кормової грудки. Його розміри і форма залежать від характеру живлення риб. Риби, які споживають велику здобич (макрофаги) через відносно довгі інтервали часу, мають великий мішко- або сифоноподібний шлунок. Риби, які живляться дрібними кормовими організмами (мікрофаги) через короткі інтервали часу, мають невеликий грудкоподібний шлунок. Порожнина шлунка у вигляді дрібних складок вистелена одношаровим слизовим епітелієм, у заглибленнях якого відкриваються отвори залозистих трубок, крізь які надходять травні ферменти. Безшлункові риби не мають шлунка, його функції виконує передній відділ кишечнику.
Кишківник риб складається з переднього, середнього і заднього відділів. Як уже зазначалося, його довжина залежить від характеру корму і здебільшого становить 20—1200 % довжини тіла риби. Причому з віком відносна довжина кишківника риб, як правило, зростає. Стінка кишківника складається із чотирьох шарів: слизового, підслизового, м'язового і серозного. Внутрішня одношарова епітеліальна слизова оболонка утворена циліндричними абсорбційними клітинами з мікроворсинками, серед яких трапляються келихо - та грушоподібні секреторні клітини. У передньому відділі кишківника риб слизова оболонка утворює поздовжні складки, у далі розташованих відділах — складну сітчасту структуру. У низькоорганізованих риб (хрящові, осетро-, лососеподібні) на межі середнього і заднього відділів кишечнику внаслідок скручування епітелію та м'язової оболонки зберігається спіральний клапан, за рахунок чого збільшується площа "робочої" поверхні кишечнику. У деяких шлункових риб (оселедце-, лососе-, тріскоподібні) на початку переднього відділу кишечнику є добре розвинені, іноді досить численні (у лососів до 400) сліпі вирости кишки — пілоричні придатки, що збільшують його травну поверхню. У слизовій оболонці цих виростів відсутні секреторні клітини, які продукують травні ферменти.
Перетравлення кормової грудки забезпечує функціонування спеціалізованих утворів — печінки та підшлункової залози. Поділена на кілька частин печінка риб синтезує рідкий секрет — жовч, який накопичується у жовчному міхурі і по спеціальних протоках потрапляє у передній відділ кишечника у міру надходження туди поживних речовин. У кишечнику жовч сприяє розщепленню, омиленню, емульгації і всмоктуванню жирів, жиророзчинних вітамінів, активує ферменти і чинить антисептичну дію. До переднього відділу кишечника прилягають дрібні протоки підшлункової залози, яка має дифузорну будову і складається з окремих розкиданих і розгалужених часточок чи трубочок.
Особливості анатомічної будови тіла риб є наслідком специфіки філогенезу, що не тільки забезпечило високий рівень адаптації до умов існування, а й процвітання риб, їх значне поширення в акваторіях планети.
Для пізнання теоретичних аспектів живлення і годівлі риб необхідно розглянути фізіологічну складову, а саме фізіологічні особливості живлення риб.
Потреби організму риб в енергії, пластичному матеріалі та елементах, необхідних для забезпечення всіх життєвих функцій, задовольняються їх травною системою. До її складу належать як органи, які безпосередньо виконують травну функцію, так і органи, які її регулюють. Органи, які виконують травну функцію, об'єднані у шлунково- кишкову або кишкову трубку з пов'язаними з нею компактними секреторними залозами, їх позначають як травний канал. Регуляторну функцію забезпечують два рівні нервової системи: на місцевому — ентеральні нервові закінчення, на центральному — відповідні структури центральної нервової системи.
Цілеспрямована травна поведінка риб формується за участю гіпоталамуса та інших відділів головного мозку. Кінцевим результатом діяльності травної системи є гідроліз харчових речовин (білків, жирів, вуглеводів) до мономерів (амінокислот, моногліцеридів, жирних кислот, моносахаридів) та їх транспорт з травного каналу до внутрішнього середовища організму. Фізико-хімічні процеси, які забезпечують цей результат, відбивають сутність травлення і всмоктування. Вони реалізуються в разі виконання травним каналом таких функцій, до яких можливо віднести: тимчасове збереження кормової грудки, розщеплення харчових речовин під дію сполук, які продукуються секреторними клітинами, моторно-евакуаційна дія на кормову грудку м'язового шару клітин, розташованих у стінці травного каналу, всмоктування мономерів епітеліальними клітинами кишківника, інкреторне виведення неперетравлених решток у зовнішнє середовище.
Однією з найважливіших характеристик ефективності дії травної системи є швидкість проходження їжі крізь шлунково-кишковий тракт. Слід зазначити, що пропускна здатність шлунково-кишкового тракту значно менша, ніж ковтального апарату. Передня частина травної системи діє як накопичувач кормової суміші, і поступово пропускає у вужчу дистальну частину невеликі порції первинно обробленого і розрідженого корму — хімусу. За рахунок цього зростає ступінь перетравлення кормової грудки і полегшується подальше всмоктування поживних речовин. Однак при цьому подовжується час перебування корму у травній системі, на що впливають температурний режим, якість корму та фізіологічний стан риби (табл. 2.2.).
Таблиця 2.2. Час спорожнення травного каналу деяких видів риб
Температура, °С |
Час спорожнення, год |
||
Короп |
Білий товстолобик |
Канальний сомик |
|
12 |
60 |
— |
90 |
17 |
35 |
16 |
35 |
20 |
30 |
13 |
28 |
24 |
24 |
10 |
20 |
28 |
- |
7 |
- |
30 |
— |
5,5 |
- |
Швидкість спорожніння травного каналу рослиноїдних риб вища, ніж всеїдних і твариноїдних. Травний канал рослиноїдних риб пристосований до пропускання крізь кишечник великої кількості низькопоживної їжі, з якої вони одержують незначну частину засвоюваних речовин. Раціон риб, який складається з більш енергетично цінного тваринного корму, утримується у травному каналі значно довше, перетравлюється повільніше, неперетравлені рештки виводяться пізніше. Існує чітко виражена обернена залежність, за якою зі зниженням енергетичної цінності їжі відповідно зростають частота живлення, інтенсивність травлення і швидкість виведення харчових решток.
За інтенсивністю процес травлення поділяють на дві стадії: "ефективну", під час якої руйнуються легкоперетравлювані компоненти корму, і "залишкову", пов'язану з руйнуванням важкоперетравлюваних компонентів корму. На першій стадії перетравлюється до 80 % маси кормової грудки. Ця процедура у хижих риб триває близько 3 діб, у мирних твариноїдних риб — близько 2, рослиноїдних — менше однієї доби. "Залишкове" травлення триває від 1,5 до 3 діб.
Істотно відрізняється ефективність травлення у шлункових і безшлункових риб. Шлунок забезпечує більшу приймальну місткість травної системи, тому шлункові риби живляться з більш вираженою періодичністю. Так, якщо періодичність живлення шлункових риб становить 1—2 доби, то безшлункових — 6—15 год.
На швидкість перетравлювання їжі у шлунку риб істотно впливає маса спожитого корму. Зі збільшенням об'єму кормової грудки знижуються ефективність травлення і засвоєння корму, гальмується моторика кишківника, підвищується частка поживних речовин, які не встигають всмоктатись, і виводяться назовні. Кількість їжі, яку риба може спожити за один прийом, значно варіює залежно від виду риби, середньої маси, сезону та екологічних умов живлення. Глибоководні хижаки здатні проковтнути рибу-жертву значно більших розмірів, ніж вони самі. Ємкість шлунка хижаків-засадників становить близько 50 % маси їх тіла. Зазвичай хижі риби поглинають їжі за один прийом від 5 до 25 % маси власного тіла. Раціон мирних риб значно менший і становить від 0,5 до 1,2 % маси їхнього тіла.
Слід пам'ятати, що тривалість перебування їжі у травній системі зростає з віком риби. Найшвидше спорожнюється травний канал у личинок риб; у личинок форелі за температури води 8 °С кишечник спорожнюється через 45—50 год., у мальків масою 2,5 г — в середньому через 60 год., у однорічок масою 150 г — через 150—200 год.
Процес перетравлення корму у риб забезпечується функціонуванням відповідних секреторних утворів. У ротову порожнину, глотку і стравохід риб секретується слиз, який не містить травних ферментів, і забезпечує лише захист епітеліальних тканин та полегшує проходження кормової грудки. Травні ферменти починають виділятись у наступних відділах травного каналу, але рівень їх ферментативної активності, енергетичні та кінетичні характеристики відрізняються залежно від типу живлення, складу їжі і фізіологічного стану організму. На відміну від вищих хребетних тварин ферменти риб менш теплотривкі і більш чутливі до зміни
температури зовнішнього середовища. Температурний оптимум ферментативної активності риб перебуває у межах 20—40 °С. Влітку вона вдвічі вища, ніж узимку, що справедливо для регіонів з чіткими порами року.
У шлункових риб утворюється кислий шлунковий сік (рН = 1,2...5). Кислотність шлункового соку риб-хижаків вища, ніж всеїдних риб. Ферменти шлунка представлені кількома типами протеаз, серед яких провідну роль відіграє пепсин. Ефективна дія цього ферменту спостерігається за рН = 2...4. Оптимальні умови травної активності пепсину забезпечуються виділенням соляної кислоти, кількість якої залежить від характеру корму, його об'єму та температури. У безшлункових риб секреція пепсину і соляної кислоти не відбувається.
Для кишкової секреції риб характерно виділення до 20 ферментів, що належать до трьох основних класів — протеази, ліпази і карбогідрази, які гідролізують три відповідні класи харчових речовин. Об'єм кишкового соку, його склад і ферментативна активність залежать насамперед від складу корму. Найвищу ферментативну активність сік має через 5—6 год. від початку годівлі риби. Простежується чітка залежність, за якою у хижих риб переважає протеолітична активність кишкового соку, у рослиноїдних - карбогідролітична. Основним протеолітичним ферментом у кишечнику риб є трипсин, менш виражені ектопептидаза і катепсин, максимальна гідролізна дія яких виявляється за рН = 7... 11. Джерело трипсину важко локалізувати, але, ймовірно, більша його частка надходить з підшлункової залози, менша — із секреторних клітин стінок кишківника, включаючи і пілоричні придатки. Ліполітична активність, пов'язана з розщепленням жирів до гліцерину і жирних кислот, виявлена в екстрактах підшлункової залози, печінки, кишківнику і пілоричних придатків. Карбогідрази, гідролізна дія яких спрямована на розщеплення вуглеводів, виявлено у великій кількості (мальтази, сахарази, лактази, целобіази, глюкозидази), більш виражені вони у рослиноїдних риб. З процесом травлення тісно пов'язана печінка, яка секретує жовч, що накопичується у жовчному міхурі. Жовч не містить ферментів, а є сумішшю органічних і неорганічних солей, пігментних речовин, холестерину, жирних кислот, білірубіну, лецетину, води. Жовч крізь відповідну протоку надходить у передній відділ кишківнику або у пілоричні придатки, забезпечує перетравлення й адсорбцію ліпідів і споріднених речовин, таких як жиророзчинні вітаміни (А, Д, Е, К), активує дію травних ферментів.
Розщеплені в процесі травлення кормові компоненти всмоктуються крізь стінку кишківнику і переносяться у кров. Засвоєння з'їденого корму у риб коливається в широких межах і залежить від ступеня перетравлення кормової грудки. У фітофагів і детритофагів, раціон яких містить значну частку баластових речовин, ступінь засвоєння корму не перевищує 20 %, для хижаків досягає 80 %.
Наведені дані дають певні уявлення стосовно фізіології живлення риб у природних і штучних умовах, що необхідно для свідомого підходу до загальної справи, якою у сучасному рибництві є годівля. Водночас не слід забувати, що рівень перебігу фізіологічних процесів тісно пов'язаний з умовами зовнішнього середовища, яке часто визначає кількісні характеристики відправлень організму і впливає на ефективність годівлі риб.
Наведена інформація дає певні уявлення стосовно різних аспектів теорії живлення і практики годівлі риб, що необхідно для свідомого підходу до такої важливої справи, як сучасні погляди на живлення риб і практику застосування годівлі в різних умовах рибництва.