ОСНОВИ МЕДИЧНОЇ БІОЛОГІЇ - 2012
Молекулярно-генетичні механізми онтогенезу. Особливості пренатального періоду розвитку людини. Порушення онтогенезу та їх місце в патології людини
Онтогенез (грец. ontos - істота, genesis - розвиток), або індивідуальний розвиток, - розвиток особини від моменту запліднення і утворення зиготи (при статевому розмноженні) і до смерті. Його основу складає реалізація генетичної інформації на всіх стадіях онтогенезу за певних умов середовища. Онтогенез тісно пов'язаний з філогенезом (історичним розвитком). Закономірності індивідуального розвитку вивчає наука - біологія розвитку. Розрізняють два типи онтогенезу - непрямий і прямий. Непрямий розвиток існує в личинковій формі, прямий - у неличинковій і внутрішньоутробній.
Непрямий, або личинковий, розвиток зустрічається у видів, яйця яких бідні на жовток. Ці організми в своєму розвитку мають одну або декілька личинкових стадій, які за будовою більше чи менше відрізняються від дорослої форми. Личинки ведуть активний спосіб життя, у більшості випадків самі добувають їжу, але в деяких видів (переважно паразитичних) мають лише органи розселення (мірацидії і церкарії сисунів, корацидії ряду стьожкових червів). Для здійснення життєвих функцій у личинок є ряд провізорних (тимчасових) органів, які відсутні в дорослому стані. Цей тип розвитку супроводжується перетворенням - метаморфозом.
Прямий розвиток. Неличинкова форма прямого розвитку має місце у видів, яйця яких багаті на жовток і його достатньо для завершення онтогенезу (риби, рептилії, птахи). Особина, яка з'являється на світ, має будову, характерну для дорослої форми, але відрізняється меншими розмірами, незрілістю органів і систем. Внутрішньоутробна форма прямого розвитку характерна для вищих ссавців і людини. Яйцеклітина при цьому типі розвитку майже не містить поживного матеріалу. Всі життєві функції зародка здійснюються через материнський організм за допомогою провізорного органу - плаценти. Цей, найбільш пізній у філогенетичному відношенні тип онтогенезу, найкращим чином забезпечує виживання зародка, але новонароджений організм не здатний до самостійного життя, тому що потребує специфічного живлення секретом молочних залоз материнського організму - молоком.
Періоди онтогенезу: ембріональний, або зародковий, і постембріональний, або післязародковий. У вищих тварин і людини прийнятий поділ на пренатальний або антенатальний (до народження) і постнатальний (після народження) періоди. Крім ембріонального і постембріонального періодів, в онтогенезі виділяють ще передзиготний період.
Передзиготний період онтогенезу - це період утворення статевих клітин (гаметогенез). Яйцеклітини і сперматозоїди розвиваються з первинних статевих клітин. У людини вони з'являються в стінці жовткового мішка через 24 доби після запліднення. У 4-тижневого зародка ці клітини виявляються в брижі задньої кишки, а ще через тиждень - у зачатку статевої залози. Утворення остаточних статевих клітин відбувається в процесі мейозу.
У передзиготний період в яйці відбуваються процеси, пов'язані із забезпеченням ранніх стадій розвитку зародка. До цих процесів належать: 1) ампліфікація (утворення копій) генів рРНК; 2) нагромадження «про запас» рРНК та іРНК для біосинтезу білків після запліднення;
3) нагромадження поживого матеріалу - жовтка.
В яйці створюється запас поживного матеріалу - жовтка. Залежно від його кількості і характеру розподілу розрізняють наступні типи яєць: алецитальні - яйця не містять жовтка; ізолецитальні - яйця містять небагато жовтка і він розміщений рівномірно по всій клітині (голкошкірі, ланцетник, ссавці); у телолецитальних яйцях жовтка багато і він зосереджений на одному з полюсів - вегетативному, протилежний полюс, який містить ядро і цитоплазму без жовтка, називається анімальним (молюскі, земноводні, рептилії, птахі); у центролецитальних яйцях жовток знаходиться в центрі клітини, а цитоплазма - на периферії (яйця комах).
Ембріональний період онтогенезу, його стадії
Ембріональний період (ембріогенез) починається з утворення зиготи і завершується виходом із яйцевих оболонок (при личинковій формі розвитку) або виходом із зародкових оболонок (при не личинковій формі розвитку). Наука, яка вивчає закономірності ембріонального розвитку, називається ембріологією. Стадії ембріонального періоду: зигота, дроблення, гаструляція, гісто- і органогенез. Зигота - одноклітинна стадія ембріонального розвитку, яка утворюється в результаті злиття сперматозоїда і яйцеклітини (запліднення).
Дробленням називають ряд мітотичних поділів зиготи, які швидко йдуть один за одним. Ріст клітин при цьому не спостерігається, збільшується лише їх кількість. З кожним поділом дочірні клітини (бластомери) стають все меншими і меншими, тому бластула, яка утворюється в результаті дроблення, майже не відрізняється за розміром від зиготи. Залежно від типу яєць дроблення може бути повним і неповним, рівномірним і нерівномірним, синхронним і асинхронним. Дроблення завершується утворенням бластули. Бластула - одношаровий багатоклітинний зародок, який зовні нагадує пухирець переважно з порожниною всередині. Шар клітин, який утворює стінку бластули, називають бластодермою, а порожнину бластули - бластоцелем або первинною порожниною. Починаючи з бластули, клітини зародка прийнято називати не бластомерами, а ембріональними клітинами. Дроблення зиготи людини повне, нерівномірне й асинхронне закінчується утворенням бластоцисти (стерробластули).
У всіх багатоклітинних тварин за бластулою настає гаструляція (утворення гаструли), яка являє собою складний процес переміщення ембріонального матеріалу з утворенням зародкових листків. У зародків нижчих багатоклітинних (губки, кишковопорожнинні) утворюються два зародкові листки - ектодерма і ентодерма. Ці тварини дістали назву двошарових. У решти тварин між екто- і ентодермою закладається ще третій зародковий листок - мезодерма. Ці тварини називаються тришаровими. У результаті гаструляції з одношарового зародка (бластули) утворюється гаструла - спочатку двошарова, а потім тришарова. Розрізняють чотири типи, або способи, гаструляції - імміграція, інвагінація, епіболія і делямінація (рис.) Імміграція (вселення) - найдавніший спосіб гаструляції, який поширений у губок і кишковопорожнинних. Окремі клітини бластодерми іммігрують (вселяються) в бластоцель і утворюють тут внутрішній шар - ентодерму, який підстилає зовнішній шар - ектодерму. Зародок стає двошаровим, бластула перетворюється в гаструлу.
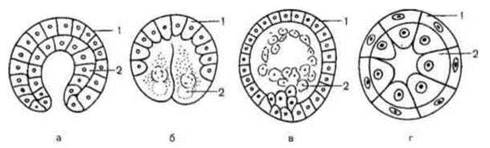
Рис. 37. Типи гаструляції:
а - інвагінація; б - епіболія; в - імміграція; г - делямінація: 1 - ектодерма; 2 - ентодерма.
Інвагінація (впинання) - найпоширеніший спосіб гаструляції. Зустрічається, наприклад, у ланцетника. Вегетативна частина бластули впинається всередину бластоцеля, у результаті чого утворюється двошаровий зародок - гаструла. У будові гаструли розрізняють: 1) стінка гаструли, яка складається з зовнішнього (ектодерми) і внутрішнього (ентодерми) зародкових листків; 2) порожнина, яка називається гастроцелем, або первинною кишкою і 3) бластопор, або первинний рот — спочатку дуже широкий, а потім звужується до невеликого отвора, який сполучає гастроцель із зовнішнім середовищем. Краї бластопора називаються губами. У первинноротих (більшість типів безхребетних) бластопор перетворюється в дефінітивний (остаточний) рот, у вторинноротих (голкошкірі, хордові) з нього формується анальний отвір або він заростає, а рот утворюється на протилежному кінці тіла.
Епіболія (обростання) зустрічається в тварин, які мають телолецитальні яйця. При цьому способі гаструляції клітини анімального полюса (не містять жовтка) бластули діляться швидше, чим клітини вегетативного полюса (перевантажені жовтком). У зв'язку з цим дрібні клітини анімального полюса поступово зовні наповзають на великі клітини вегетативного полюса, обростаючи їх і утворюючи ектодерму, а клітини вегетативного полюса формують ентодерму.
При делямінації (розшаруванні) клітини зародка діляться паралельно його поверхні, утворюючи зовнішній і внутрішній зародкові листки (кишковопорожнинні). Часто зустрічається змішаний тип гаструляції, коли одночасно відбуваються впинання, обростання й імміграція (земноводні).
Основні способи утворення мезодерми - телобластичний і ентероцельний. При телобластичному способі початок мезодермі дають дві клітини (телобласти), які симетрично розташовані по боках первинної кишки між ектодермою та ентодермою (молюски, черви, членистоногі). Ентероцельним способом мезодерма утворюється в голкошкірих, хордових. З двох боків від первинної кишки (ентодерми) утворюються випинання - кишені (целомічні мішки). Всередині кишенів знаходиться порожнина, яка є продовженням первинної кишки (гастроцеля). Целомічні мішки повністю відшнуровуються від первинної кишки і розростаються між ектодермою і ентодермою. Клітини стінки мішків дають початок мезодермі, порожнина - вторинній порожнині (целому). Дорзальний відділ мезодерми, який лежить по боках від нервової трубки і хорди, розчленований на сегменти - соміти. Вентральний її відділ утворює суцільну бічну пластинку, яка знаходиться по боках кишкової трубки. Соміти диференціюються на три відділи - медіальний (склеротом), центральний (міотом) і латеральний (дерматом). У вентральній частині мезодермальної закладки розрізняють нефрогонотом (ніжка соміта) і спланхнотом. Закладка спланхнотома розділяється на два листки, між якими утворюється порожнина. На відміну від бластоцеля вона отримала назву вторинної порожнини, або целома. Вісцеральний листок (спланхноплевра) межує з ентодермальною кишковою трубкою, парієтальний (соматоплевра) лежить безпосередньо під ектодермою.
У людини перший етап гаструляції проходить переважно за типом деламінації, а другий етап шляхом виселення клітин (імміграція). Утворення мезодерми здійснюється ентероцельним способом. На стадії гаструляції зародок імплантується (проникає) у слизову оболонку матки.
Першими відомостями про зародкові листки наука зобов'язана російським академікам К.Ф. Вольфу, Х.І. Пандеру і К.М. Беру. Численні дослідження з вивчення ранніх стадій розвитку тварин належать О.О. Ковалевському і І.І. Мечникову. Засновником сучасної теорії зародкових листків по праву вважається О.О. Ковалевський, який встановив схожість зародкових листків у різних типів тварин не лише за походженням, але й за похідними, що з них утворюються (1901).
Гістогенез і органогенез. Гістогенез - процес утворення тканин, органогенез - процес формування органів. В органогенезі виділяють дві фази: 1) нейруляція - утворення осьових органів (нервова трубка, хорда) і 2) побудова решти органів. Зародок на стадії нейруляції називається нейрулою. Початок всім тканинам і органам дають зародкові листки. З ектодерми розвиваються: нервова система, епідерміс шкіри та його похідні (пір'я, волосся, нігті, шкірні й молочні залози), епітелій ротової порожнини, емаль зубів, сприймаючі клітини органів зору, слуху, нюху. Похідними ентодерми є епітелій шлунково-кишкового тракту і залоз - залозистої частини печінки, підшлункової залози, слинних залоз, а також епітелій органів дихання та їх залоз. На початок стадії органогенезу мезодерма представлена сомітами. Дерматом утворює сполучнотканинний компонент шкіри (дерму), склеротом бере участь в утворенні скелетних структур, міотом дає початок скелетній мускулатурі, нефрогонотом - органам виділення і статевим залозам, спланхнотом - сполучній тканині внутрішніх органів, кровоносним судинам, гладкій мускулатурі кишок, дихальних і сечостатевих шляхів. Матеріал спланхо- і соматоплеври використовується для розвитку статевої, серцево-судинної і лімфатичної систем, плеври, очеревини, перикарда. Органогенез завершується в основному на кінець ембріонального періоду розвитку. Проте диференціювання та ускладнення органів продовжується і у постембріональний період.
Провізорні органи. Зв'язок зародка із середовищем забезпечується спеціальними позазародковими органами, які функціонують тимчасово і називаються провізорними. Міра розвитку і функція цих органів різна. Для всіх тварин з неличинковим типом розвитку властивий такий провізорний орган, як жовтковий мішок. У ссавців жовтковий мішок є похідним спланхноплеври, яка згодом розщеплюється на внут-рішньозародкову і позазародкову частини. Із останньої формується жовтковий мішок. У стінки жовткового мішка вростають кровоносні судини, які утворюють по всій поверхні жовтка густу капілярну сітку. Клітини стінки жовткового мішка виділяють ферменти, які розщеплюють поживні речовини жовтка, що надходять у кровоносні капіляри і далі у організм зародка. Жовтковий мішок — це також перший кровотворний орган зародка, місце розмноження клітин крові. У ссавців редукований жовтковий мішок входить до складу плаценти. Призначення цих провізорних органів — забезпечення життєвих функцій зародка у різноманітних умовах середовища.
У дійсно наземних тварин, які втратили зв'язок з водним середовищем, зародки розвиваються у спеціальних оболонках. Такою оболонкою є перш за все амніон, який наповнений рідиною. Амніон здійснює функції обміну і захисту від висихання і механічних впливів. Амніотична рідина, у яку занурений зародок, що розвивається, являє собою водний розчин білків, цукрів, мінеральних солей, містить також гормони і сечовину. У процесі розвитку склад цього середовища змінюється. В акушерській практиці амніотичну рідину, яка виділяється перед родами, називають водами.
У ссавців і людини яйцеклітина бідна на білок, тому провізорні органи мають свої особливості. Жовтковий мішок закладається на ранніх етапах ембріогенезу, але не розвивається, а поступово редукується, розшаровується. Алантоїс (зародкова оболонка) також не розвинений. Зачаток його входить до складу нового специфічного провізорного органу — пупкового канатика. Функцію зовнішньої зародкової оболонки виконує хоріон, або ворсинчаста оболонка. Численні вирости, ворсинки хоріона (звідси і назва оболонки) вростають у слизову оболонку матки — спеціального органа материнського організму, який є тільки у ссавців. Місце най - більшого галуження ворсинок хоріона і найбільш тісного контакту їх із слизовою оболонкою матки носить назву дитячого місця, або плаценти.
Зв'язок тіла зародка з плацентою здійснюється через пуповину або пупковий канатик, який містить кровоносні судини. Кровоносні капіляри тіла зародка галузяться у ворсинках хоріона. Так встановлюється плацентарний кровообіг. Кров матері не змішується з кров'ю плода; вона обмиває ворсинки хоріона, але ніколи не проникає у капіляри плода. Через плаценту плід постачається поживними речовинами, киснем і звільняється від продуктів життєдіяльності. При цьому важлива роль належить епітеліальним клітинам, які утворюють хоріон і його ворсинки. Епітелій хоріона і стінок судин утворює специфічний клітинний бар'єр; мікроорганізми і ряд речовин із кровотоку матері у нормі не потрапляють у кровоток плода. Порушення плацентарного бар'єру, як правило, веде до розладу нормального розвитку плода, до патології вагітності. Плацента є бар'єром для ряду лікарських речовин, у тому числі наркотиків, виробничих і харчових отрут, чужорідних білків і антитіл. Вивчення біологічних особливостей зв'язку організму плода і матері у вищих ссавців, а отже, і у людини, має велике значення і лежить у основі правильної організації медичної служби у сфері охорони материнства.
Механізми росту і морфогенезу
В ембріогенезі відбувається інтенсивний ріст і морфогенез. Морфогенез - це процес виникнення нових структур і зміна їх форми в процесі онтогенезу. Морфогенез виражається в утворенні тканин, органів, завдяки чому організм у цілому та його окремі частини набирають форми, типової для дорослих станів. Морфогенез - процес незворотний. Він відбувається лише за певних умов. Основні механізми морфогенезу: детермінація, проліферація (розмноження клітин), міграція (переміщення клітин), сортування клітин, запрограмована елімінація (загибель), диференційовка клітин, індукція і компетенція (контактні взаємодії), дистантна взаємодію клітин, тканин і органів (гуморальні, нервові механізми інтеграції).
Ділення клітин (проліферація) забезпечує утворення із зиготи багатоклітинного організму; визначає його ріст, вибіркове розмноження клітин забезпечує морфогенетичні процеси. Експерименти показали, що ізольовані бластомери на ранніх стадіях дроблення тотіпотептні (рівноспадкові), тобто кожний з них розвивається в повноцінний організм. У багатьох гідромедуз клітини тотіпотентні до стадії 32, у тритона - до стадії 16, у кроликів - до стадії 2 бластомерів. Про існування подібної тотіпотентності бластомерів у людини свідчать випадки народження двох, трьох, чотирьох і, навіть, семи монозиготних близнят.
Міграція клітин, або клітинні переміщення, поряд з іншими клітинними процесами мають велике значення, починаючи з процесів гаструляції і далі у процесах морфогенезу. Порушення міграції клітин у ембріогенезі призводить до недорозвитку органів, зміни нормальної локалізації, тобто виникнення природжених вад розвитку. В процесі ембріогенезу сортування клітин. Вони не тільки активно переміщуються, але й «впізнають» одна одну, тобто утворюють скупчення, пласти тільки з відповідними клітинами. Значні переміщення клітини характерні для періоду гаструляції (утворення зародкових листків).
В процесі розвитку зародка поряд з розмноженням велике значення мають процеси загибелі клітин. Розрізняють два типи клітинної загибелі: апоптоз та некроз. Апоптоз - фізіологічний процес досягнення характерних для відповідного біологічного виду рис його морфофункціональної організації. Апоптоз є природним, еволюційно обумовленим механізмом морфогенезу, що генетично контролюється. Некроз клітин виникає у нефізіологічних умовах, наприклад, у зв’язку з дією несприятливих факторів: стійке кисневе голодування, токсини, тощо. Некроз супроводжується запаленнями і є патологічним процесом.
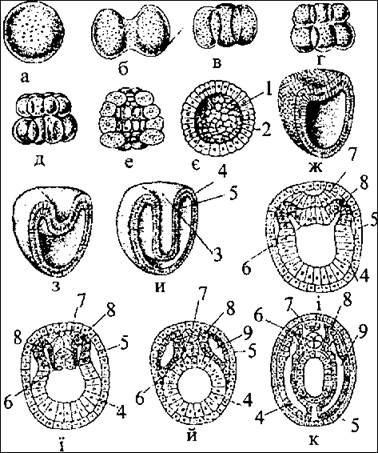
Рис. 38. Розвиток зародка ланцетника:
а - стадія зиготи; б-е - стадія дробіння (2, 4, 8, 16, 32 бластомери); є-бластула (у розрізі); ж-и - стадія гаструли (у розрізі); і-к - послідовні стадії закладання тканин і органів зародка; 1 -бластоцель; 2 - бластодерма; 3 - бластопор (рот гаструли); 4 - ентодерма; 5 - ектодерма; 6 - хорда; 7 - нервова пластинка; 8 - закладання мезодерми; 9 - вторинна порожнина (целом).
На стадії бластули зародкові клітини втрачають тотіпотентність. Починається диференціювання - процес утворення відмінностей між клітинами, які походять з відносно однорідного матеріалу зародка. Клітини стають спеціалізованими, тобто набувають хімічних, морфологічних та функціональних особливостей. У процесі диференційовки клітини ембріональних закладок перетворюються в певний елемент дорослого організму. Вище було показано, що міотом дає початок скелетній мускулатурі, дерматом утворює дерму шкіри і т.д. Отже, кінцевий результат розвитку окремих ембріональних закладок визначений. Процес визначеності, зумовленості розвитку клітини лише в певному напрямку називають детермінацією. На стадії бластули клітини, хоч і втрачають тотіпотентність, але здатні ще до перевизначеності (трансдетермінації) шляху подальшого розвитку. Досліди на зародках амфібій показали, що перехресні пересадки певних ділянок ведуть до зміни подальшого розвитку відповідно до їх нового положення. Результати аналогічних пересадок на стадії гаструли свідчать про втрату здатності клітинного матеріалу до трансдетермінації. Таким чином, лабільна детермінація шляхів розвитку, яка спостерігається на ранніх стадіях, змінюється на стабільну. Диференціація й детермінація відбуваються на фоні інтеграції. Інтеграція — об'єднання і узгодження структур і функцій багатоклітинного організму в процесі ембріогенезу. На всіх стадіях зародок функціонує як єдиний цілісний організм, в якому всі процеси взаємопов'язані і взаємозумовлені.
Рис. 39. Плід людини:
а - 8 тижнів; б - 9 тижнів; в - 10 тижнів; г - 11 тижнів; д - 12 тижнів; е - 16 тижнів.
Ріст - це збільшення загальної маси в процесі розвитку, яка призводить до постійного збільшення розмірів організму. Основу росту складає поділ клітин, який збільшує число клітин, і збільшення маси органічної речовини, головним чином білків. Ріст забезпечується наступними механізмами: збільшенням розмірів клітини, зростанням числа клітин,
накопиченням неклітинної речовини, продуктів життєдіяльності клітин.
Розрізняють два типи росту: обмежений і необмежений. Необмежений ріст триває впродовж усього періоду онтогенезу (наприклад, у риб). При обмеженому рості - ріст відбувається в певні періоди онтогенезу, а згодом уповільнюється і гальмується. Швидкість росту неоднакова в різних ділянках організму і на різних стадіях розвитку. Максимальна швидкість характерна для перших чотирьох місяців ембріонального розвитку, що пояснюється інтенсивним поділом клітин. По мірі росту плода кількість мітозів зменшується і після шести місяців внутрішньоутробного розвитку майже не утворюються нові м’язові і нервові клітини.
Ембріональна індукція
Ембріональною індукцією називають вплив однієї частини зародка, що виникла раніше, на спрямований розвиток іншої частини, яка виникає пізніше. Основу ембріональної індукціі складає взаємодія між частинами зародка. Початок вивченню цього явища поклав дослід німецького ембріолога Г.Шпемана (1924). У ньому при нормальному розвитку тритона ектодерма, яка знаходиться на спинному боці зародка, дає початок нервовій трубці. Проте, якщо на більш пізній стадії гаструляції видалити ділянку верхньої губи бластопора, яка прилягає до ектодерми на спинному боці зародка, то нервова трубка не утворюється. Якщо ж спинну губу бластопора пересадити під ектодерму черевного боку, то тут виникає спочатку нервова трубка, інші осьові органи (хорда, соміти), а потім формується додатковий зародок. Спостереження за розподілом пігментованих і непігментованих клітин показали, що тканини формуються виключно з клітинного матеріала реципієнта. Ділянка зародка, яка визначає розвиток іншої ділянки в певному напрямку, була названа організаційним центром (організатором, індуктором). Дорзальна губа бластопора є первинним індуктором (організатором) в амфібій. У риб йому відповідає дорзальний край бластодиска, у птахів - первинний вузлик. Спинна губа має здатність організовувати, запускати розвиток зародка. Ніякі інші клітини цією здатністю не володіють. За відсутності спинної губи розвиток не відбувається. Губа індукує розвиток нервової трубки, а сама диференціюється в хорду і соміти. Первинна ембріональна індукція - перший крок у ланцюгу послідовних (вторинних, третинних) індукційних процесів у подальшому розвитку. Зачаток майже будь-якого органу проходить у своєму розвитку дві фази. У фазі залежної диференційовки його доля залежить від дії індуктора і зовнішнього середовища - пересадка в нове місце приведе до трансдиференціювання (пересадка губи бластпора в будь-яке місце викликає там розвиток нервової трубки); у фазі незалежної диференційовки - не приведе до зміни шляху розвитку. Подальші дослідження проблеми ембріональної індукції йшли в напрямку визначення природи індукторів. Х. Тидеман у 1970 р. виділив із зародків курчат індуктори білкової природи. Один з них індукує структури нервової системи і органів чуттів, інший - м'язову систему. Індуктори входять в ядро і призводять до прояву активності відповідних генів.
Генетичний контроль розвитку
Загальні закономірності онтогенезу багатоклітинних пояснюються гіпотезою диференціальної активності генів. На різних етапах розвитку активними є різні структурні гени. Вони зумовлюють диференційовку клітин. Головний механізм клітинної диференційовки зводиться до вибіркового блокування - деблокування (інактивація- активація) окремих генів або їх груп, що добре можна спостерігати на велетенських політенних хромосомах личинок дрозофіли, інших двокрилих. Активні зони таких хромосом - здуття (пуфи) - в залежності від стадії розвитку пересуваються по довжині хромосоми, число їх змінюється. Ці ділянки - місця активного синтезу іРНК; вони відповідають генам, які включаються на певних стадіях життєвого циклу.
З 40 тисяч генів морського їжака на стадії бластули активні 30 тисяч, гаструли і личинки - 12-15 тисяч, у дорослих тварин - 3-5 тисяч генів. У клітинах дорослої людини приблизно 90 % генів не "працюють". Багато з них виконали свою функцію на різних етапах онтогенезу і знаходяться в заблокованому стані. Гени в процесі диференційовки соматичних клітин не втрачаються. Окремі групи генів втрачають лише свою активність. Гени, які при спеціалізації клітини втратили свою активність, можуть знову "запрацювати" після пересадки ядра в яйцеклітину, про що свідчать досліди англійського ембріолога Дж. Гердона.
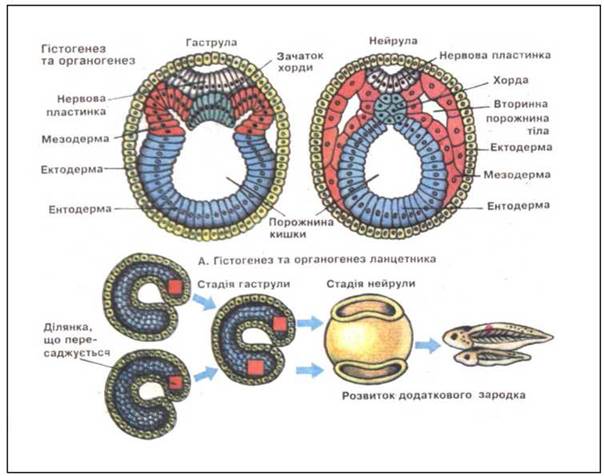
Рис. 40. Взаємодія частин зародка, що розвивається (явище ембріональної індукції)
Особливості пренатального періоду розвитку людини, критичні періоди
У пренатальному (до народження) розвитку людини виділяють три періоди: 1) початковий (преембріональний); 2) зародковий (ембріональний) і 3) плодовий (фетальний). Початковий періодтриває після запліднення протягом першого тижня вагітності. Зародковий період (2-8-й тижні) називають періодом органогенезу. Протягом цього періоду три зародкові листки (екто-, енто- і мезодерма) дають початок всім тканинам і системам органів. Процеси органогенезу дуже чутливі до ушкоджувальної дії генетичних факторів і факторів зовнішнього середовища. Протягом цього періоду виникає більшість великих вад розвитку. Наприкінці цього періоду ембріон стає плодом з характерними для дитини обрисами тіла. Плодовий період (від 9-го тижня вагітності до народження) характеризується швидким ростом тіла і дозріванням систем органів.
Критичні періоди - це періоди, коли зародок найбільш чутливий до впливу несприятливих факторів зовнішнього середовища. Поняття про критичні періоди розвитку було введене в 1921 р. К.Стоккардом і розвинуте П.Г. Свєтловим. У розвитку людини виділяють наступні критичні періоди: 1) прогенез (гаметогенез); 2) запліднення; 3) імплантація (6-7 доба після зачаття); 4) плацентація (у людини 3-6-й тиждень вагітності); 5) гісто- і органогенез (3-4-й місяць ембріогенезу); 6) перинатальний (пологи). З останнім критичним періодом пов'язані різкі зміни умов існування і перебудова діяльності всіх систем організму новонародженого (змінюється характер кровообігу, газоообміну, живлення). Крім цього, встановлені критичні періоди розвитку окремих органів протягом життя людського ембріона.
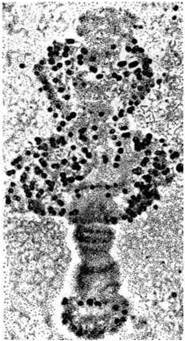
Рис. 41. Пуфи в хромосомах (за Дубиніним):
мікрофотографія хромосоми з клітини слинної залози дрозофіли, яка має на даній стадії три великі пуфи (здуття); чорні цятки - місця включення радіоактивного уридину, що свідчить про активний процес синтезу в цих місцях молекул РНК.
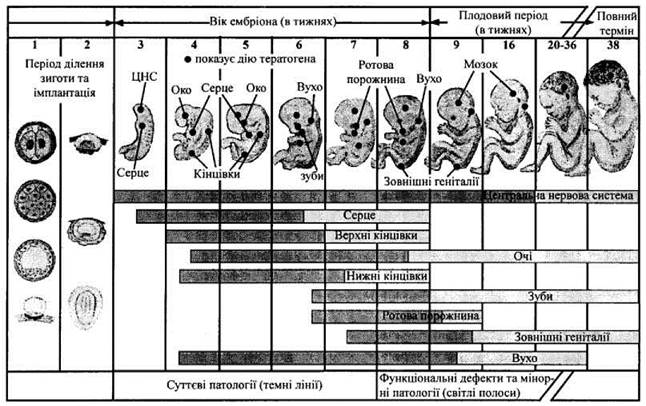
Рис. 42. Ефекти тератогенів на стадіях розвитку ембріонів та плодів
Тератогенні фактори
Тератогенні фактори, або тератогени (грец. teratos - виродок, потвора) - це фактори середовища, які порушують ембріогенез і зумовлюють природжені вади розвитку. Процес виникнення природжених вад розвитку називають тератогенезом, а науку про тератогени і тератогенез - тератологією. Розрізняють фізичні, хімічні та біологічні тератогени.
До фізичних тератогенів належать: іонізуюче випромінювання, висока і низька температура, механічні чинники (тиск, струс, механічні травми). До хімічних тератогенів відносять хімічні речовини (у тому числі деякі лікарські препарати): антагоністи фолієвої кислоти (аміноптерин), валпроєва кислота, андрогени, кумаринові антикоагулянти, ретинол (надлишок), свинець, органічні сполуки ртуті, тетрациклін, талідомід, триметадіон, алкоголь, кокаїн, ін. Прикладів впливу хімічних речовин на виникнення природжених вад розвитку багато. Так, в Японії народилося багато дітей з неврологічними симптомами, які нагадували церебральний параліч. Було встановлено, що риба, яку вживали матері цих дітей, містила високий рівень органічних сполук ртуті, які потрапляли з промисловими відходами в затоку Мінамата. Хвороба одержала назву хвороба Мінамата. Подібні випадки зафіксовані в США, коли зерном кукурудзи, обробленим фунгіцидом, що містив ртуть, годували свиней, а їхнє м'ясо споживали вагітні жінки. У 60-х роках у Західній Європі народилося понад 7000 дітей з амелією та меромелією (повна або часткова відсутність кінцівок), вадами серця. З'ясувалося, що причиною був талідомід (снодійний препарат і засіб проти нудоти), який матері цих дітей приймали на ранніх стадіях вагітності. В усіх розвинутих країнах після трагічної історії з талідомідом були прийняті законодавчі акти, згідно з якими кожний новий лікарський препарат перед тим, як його застосовувати в клініці, повинен пройти випробування на зародках тварин на тератогенність.
Також причиною природжених вад є вживання матір'ю алкоголю. Комплекс цих вад описаний як алкогольний синдром плода (вузькі очні щілини, гіпоплазія верхньої щелепи, серцеві вади, розумова відсталість). Частота алкогольного синдрому плода - 1-2 випадки на 1000 новонароджених. Шкідливим є навіть помірне вживання спиртних напоїв під час вагітності. Небезпеку становить тютюнопаління. Компоненти тютюнового диму пригнічують розвиток плода, призводять до зменшення маси тіла новонародженого, передчасним пологам, а деякі з них є тератогенами. Шкідливим є і "пасивне паління": у сім'ях, де палить батько, вади розвитку в дітей реєструються в 2 рази частіше, ніж у родинах, в яких батько не палив.
Біологічні тератогени - це збудники інфекційних та інвазійних захворювань (TORCH-інфекція): вірус краснухи (rubeola), цитомегаловірус, вірус простого герпесу, вірус вітряної віспи, токсоплазма (Toxoplasma gondii), збудник сифілісу, вірус вітряної віспи. Так, під впливом вірусу краснухи виникає катаракта, глаукома, серцеві вади, глухота. Цитомегаловірус спричинює мікроцефалію, сліпоту, розумову відсталість, смерть плода. При зараженні збудником токсоплазмозу розвивається гідроцефалія (водянка головного мозку), мозкові кальцифікати, мікрофтальмія (малий розмір ока). Тератогенний вплив мають деякі порушення метаболізму в матері (цукровий діабет, фенілкетонурія). Ризик виникнення природжених вад розвитку в дітей від матерів, хворих на цукровий діабет, фенілкетонурію, вищий порівняно з потомством від здорових матерів.
Дія тератогенів залежить: 1) від генотипу матері і плода; 2) від стадії розвитку: більшість великих вад розвитку виникає протягом третього-восьмого тижнів вагітності (тератогенний період), коли формуються органи. Проте зародок чутливий до тератогенів до і після нього. Тому практично не існує періодів вагітності, безпечних щодо дії на ембріон і плід тератогенів; 3) від дози і тривалості дії тератогенів.
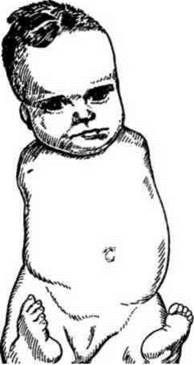
Рис. 43. Талідомідна ембріопатія як наслідок впливу екзогенного фактору на вагітну жінку (вживання талідоміду).
Природжені вади розвитку, їх класифікація
Природжені вади розвитку - це стійкі морфологічні відхилення в будові органа або цілого організму, які виникають внаслідок порушення морфогенезу у внутрішньоутробному періоді (інколи після народження дитини). За даними ВООЗ, частота природжених вад розвитку в окремих країнах складає від 2,7 до 16,3 %. За різними даними, природжені вади розвитку становлять 20 % серед причин дитячої смертності. Серед дітей першого року життя від природжених вад гине в Україні понад 1000 малюків щорічно. Більшість вад розвитку виникають при моногенних і мультифакторіальних захворюваннях. До ВВР відносять наступні порушення розвитку:
Аплазія (агенезія) — природжена відсутність органу.
Гіпоплазія (гіпотрофія) — природжене недорозвинення органу (нестача маси або розмірів органу, який перевищує два стандартних відхилення від середніх для даного віку показників).
Гіперплазія (Гіпертрофія) — природжене збільшення маси і розмірів органів внаслідок збільшення кількості (гіперплазія) або об’єму (гіпертрофія) клітин.
Макросомія (гігантизм) — збільшення довжини тіла.
Гетеротопія — наявність клітин або тканин одного органу в іншому, або в тих ділянках того ж органу, де вони повинні бути.
Ектопія — розташування органу в незвичному місці.
Дуплікація — подвоєння, а також збільшення у кількості органів або їх часток (подвоєння матки, подвійна доза аорти, поліспленія).
Стеноз звуження каналу або отвору.
Атрезія — відсутність природного каналу або отвору.
Нерозділення органів (синдактилія, симподія, сирепомелія) або двох симетрично, або асиметрично з’єднаних близнюків.
Персистування — залишки ембріональних структур (наприклад, зона метонедорогенної ділянки у нирці новонародженого, баталова протока у дитини старше 3 місяців).
Дизрафія (арафія) — незрощення ембріональних щілин (розщелина губи, незмикання кісток черепа і хребта, частіше супроводжується черепно- і спинномозковими грижами).
Порушення лобуляції— зміна кількості долей (легень, печінки, нирок).
Утворення несправжніх водянок (гідроцефалія, гідронефроз).
Інверсія — зворотнє (дзеркальне) розташування органів.
Залежно від причини всі природжені вади поділяють на спадкові, екзогенні і мультифакторіальні. Спадкові вади розвитку зумовлені мутаціями в гаметах батьків, екзогенні - дією на ембріон і плід факторів зовнішнього середовища (тератогенів), мультифакторіальні — поєднанням екзо- та ендогенних чинників. Залежно від стадії, на якій виникають порушення ембріогенезу, розрізняють гаметопатії, бластопатії, ембріопатії та фетопатії. Гаметопатії виникають на стадії утворення гамет, бластопатії - на стадії бластули, ембріопатії - у період від двох до 8 тижнів, фетопатії - від 9-го тижня внутрішньоутробного розвитку. Вади розвитку бувають первинними і вторинними. Первинні вади зумовлені безпосередньою дією тератогенного фактора, вторинні є ускладненням первинних і завжди патогенетично з ними пов'язані.
Усі природжені вади розвитку поділяють на філогенетично зумовлені і нефілогенетичні. Філогенетично зумовлені - це природжені вади розвитку, які за будовою нагадують ознаки предків або сучасних груп хордових тварин. До них належать: незростання дужок хребців, шийні або поперекові ребра, незарощення твердого піднебіння. Філогенетично зумовлені вади розвитку показують генетичний зв'язок людини з іншими хребетними. Нефілогенетичні - це такі вади розвитку, які не мають аналогів у сучасних хребетних тварин або їх предків: наприклад, зрощені (сіамські) близнята.
Анцестральні (атавістичні) вади розвитку, або атавізми - прояв ознак далеких предків. До таких вад належать: надмірне оволосіння, багатососковість, збереження подовженого куприка (хвоста). Вивчення тератогенних факторів і природжених вад розвитку вказує на необхідність охорони матері і дитини від шкідливих факторів середовища, особливо в тератогенному періоді (другий - восьмий тижні вагітності).