МЕДИЧНА БІОЛОГІЯ, АНАТОМІЯ, ФІЗІОЛОГІЯ ТА ПАТОЛОГІЯ ЛЮДИНИ - Я.І.Федонюк 2010
БІОЛОГІЯ
РОЗДІЛ 1. БІОЛОГІЧНІ ОСНОВИ ЖИТТЄДІЯЛЬНОСТІ ЛЮДИНИ
1.3. МОЛЕКУЛЯРНО-ГЕНЕТИЧНИЙ І КЛІТИННИЙ РІВНІ ОРГАНІЗАЦІЇ ЖИТТЯ
1.3.2. Структурно-хімічна функціональна організація еукаріотичних клітин
Органели цитоплазми
Органели - постійні структури цитоплазми, які мають певну будову і виконують спеціалізовану функцію. Одні органели (мікроскопічні) видно в світловий мікроскоп, інші - лише в електронний (субмікроскопічні). За наявністю біологічної мембрани їх поділяють на мембранні і немембранні. До мембранних органел належать: ендоплазматична сітка, мітохондрії, лізосоми, комплекс Гольджі, пероксисоми; до немембранних - рибосоми, центросома, мікрофіламенти, мікротрубочки. Ці дев'ять органел називають органелами загального призначення затією ознакою, що вони присутні у всіх клітинах. Виділяють також органели спеціалізованих клітин — органели спеціального призначення: міофібріли м'язових клітин і волокон, нейрофібрили нервових клітин, війки епітелію трахей, бронхів та ін.
Ендоплазматична сітка (вакуолярна сітка,ретикулум) - субмікроскопічна органела загального призначення, яка являє собою систему мембран, що формує взаємопов'язану сітку канальців, мішечків і цистерн, які пронизують гіалоплазму в усіх напрямках, контактують з мембранами ядра і плазматичною мембраною, утворюючи єдину замкнену циркуляторну систему. Основні функції ендоплазматичної сітки - синтез і транспортування органічних речовин. Розрізняють шорстку (гранулярну) і гладку (агранулярну) ендоплазматичну сітку. На мембранах гранулярної ендоплазматичної сітки з боку гіалоплазми розміщені рибосоми, на мембранах гладкої ендоплазматичної сітки вони відсутні. На рибосомах синтезуються білки, які по каналах гранулярної ендоплазматичної сітки транспортуються в апарат Гольджі, де зазнають хімічних перетворень і мембранної упаковки. На мембранах гладкої ендоплазматичної сітки синтезуються ліпіди, вуглеводи
(глікоген). Вважають, що на агранулярному ретикулумі починається процес утворення біологічних мембран.
Рибосоми - субмікроскопічні немембранні органели загального призначення, в яких відбувається біосинтез білка. Діаметр рибосом - близько 20 нм. Вони вільно розміщуються в цитоплазмі або прикріплюються до мембран гранулярної ендоплазматичної сітки та ядра. Рибосома складається з двох субодиниць: великої і малої, які здатні до зворотної дисоціації (можуть саморозпадатися і самозбиратися). Кожна субодиниця побудована з єдиного рибонуклеопротеїного тяжа. Під час синтезу білка рибосоми об'єднуються на молекулі ІРНК у комплекси - полісоми (полірибосоми).
Лізосоми (гр. lisis - розчинення, soma - тіло) - субмікроскопічні органели загального призначення, основною функцією яких є розщеплення біополімерів різного хімічного складу (клітинне травлення). Мають вигляд одномембранних замкнених мішечків діаметром 0,2-0,4 мкм з набором гідролітичних ферментів. У лізосомах перетравлюються чужорідні часточки, які надходять в клітину (гетерофагія), а також структури самої клітини, які відмерли або втратили своє функціональне призначення (аутофагія). У нормі процес травлення здійснюється всередині лізосом. У випадку руйнування стінки лізосом лізосомні ферменти виходять у цитоплазму і тоді вся клітина зазнає самоперетравлення і гине (аутоліз). Лізосоми бувають: прелізосоми, власне лізосоми і постлізосоми. Прелізосоми (первиннілізосоми) - це неактивні лізосоми, які містять ферменти і не містять субстрату для перетравлення. Первинні лізосоми формуються в комплексі Гольджі. Власне лізосоми (вторинні лізосоми) - це активні лізосоми, які містять ферменти і субстрат для розщеплення. Вони утворюються при злитті прелізосом і фагосом. Фагосомами називають піноцитозні, фагоцитозні та інші пухирці; усі вони містять субстрат для перетравлення. Вторинні лізосоми, в яких завершився процес травлення, називають постлізосомами, або залишковими тільцями; вони містять лише неперетравлені рештки. Одні залишкові тільця переміщуються до плазматичної мембрани, зливаються з нею і випорожнюються, інші лишаються в клітині. Недостатність чи відсутність того чи іншого лізосомного ферменту призводить до нагромадження в клітині неперетравлених речовин, що зумовлює розвиток так званих лізосомних хвороб нагромадження, вони переважно мають спадкову природу. З дефіцитом лізосомних ферментів пов'язують розвиток атеросклерозу, ожиріння.
Мітохондрії (грец. mitos - нитка, chondros - зернятко) - мікроскопічні мембранні органели загального призначення, основною функцією яких є вилучення енергії з органічних речовин шляхом їх окислення і нагромадження енергії, що звільнилася, в молекулах АТФ. У формі АТФ енергія використовується в життєдіяльності клітини. Розмір мітохондрій -1-10 мкм. За формою вони бувають округлими, паличкоподібними, можуть змінювати форму, зливатися, переміщуватися, зосереджуючись при патологічних станах переважно біля ядра клітини.
Мітохондрії складаються з матрикса (основної речовини) і двомембранної стінки. Зовнішня мемрана має рівні контури, а внутрішня утворює гребені (кристи), спрямовані всередину (рис. 1.32). Кристи збільшують поверхню мембран і є місцем розташування ферментів дихального ланцюга (ланцюга перенесення електронів) і окислювального фосфорилювання. Мітохондрії мають власну ДНК, розташовану в матриксі, і здатні до самовідтворення. Вони ніколи не виникають заново, а завжди утворюються лише в результаті поділу. Мітохондральна ДНК має форму кільця (як у прокаріотів). Гени мітохондріальної ДНК кодують будову деяких мітохондріальних білків, автономний синтез яких відбувається на мітохондріальних рибосомах. Проте більша частина мітохондральних білків синтезується поза мітохондріями під контролем ядерної ДНК. Рибосоми мітохондрій, як і рибосоми прокаріотів, мають константу седиментації 70S. Існує гіпотеза, що мітохондрії в далекі часи були вільноживучими прокаріотами, але за певних умов проникли в клітину і стали симбіонтами.
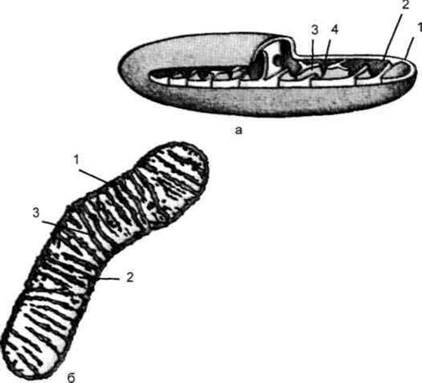
Рис. 1.32. Мітохондрія:
а - загальна схема будови: 1 - зовнішня мембрана; 2 - внутрішня мембрана; 3 - кристи; 4 - матрикс;
б - схема внутрішньої будови: 1 - двомембранна оболонка; 2 - матрикс; 3 - кристи.
Комплекс Гольджі- мікроскопічна мембранна органела загального призначення, яка побудована з диктіосом числом від кількох сотень до кількох тисяч на клітину. Диктіосома являє собою сукупність пов'язаних між собою цистерн товщиною близько 25 нм, сплющених у центральній частині і розширених на периферії. Від розширених країв цистерн відшнуровуються дрібні пухирці (пухирці Гольджі). Образно диктіосому порівнюють зі стосом тарілок, повернутих ввігнутим боком до ядра. У комплексі Гольджі нагромаджуються продукти внутрішньоклітинної секреції, які підлягають виведенню з клітини. Тут вони зазнають деяких хімічних перетворень, мембранної упаковки і в складі пухирців Гольджі транспортуються в цитоплазму або через плазматичну мембрану виводяться назовні (екзоцитоз). У комплексі Гольджі формуються первинні лізосоми, синтезуються полісахариди, утворюються комплекси цих сполук з білками (глікопротеїни) і жирами (гліколіпіди). Комплекс Гольджі розташований звичайно біля ядра.
Центросома (клітинний центр) - мікроскопічна немембранна органела загального призначення, яка бере активну участь у поділі клітини. В інтерфазі розміщується близько ядра на одній осі з центром ядра і центром клітини; складається з двох центріолей і центросфери навколо них. При підготовці клітини до поділу кожна центріоля подвоюється. Під час поділу клітини подвоєні центріолі (диплосоми) розходяться до
протилежних кінців і формують полюси клітини. Центріолі стимулюють тубулінових білків, що зумовлює утворення мікротрубочок веретена поділу. Центросому містять всі клітини тварин і людини, крім яйцеклітин, а також клітини нижчих рослин. Клітини вищих рослин центросоми не мають.
Пероксисоми - субмікроскопічні мембранні органели загального призначення, основною функцією яких є звільнення клітини від токсичних продуктів обміну речовин (детоксикація). Пероксисоми утилізують хімічно активний атомарний кисень, розщеплюють етиловий спирт, сечову кислоту, ін. Вони мають вигляд мішечків діаметром близько 0,2-0,5 мкм, заповнених ферментами, з них маркерним (визначальним) ферментом є каталаза.
Мікрофіламенти - субмікроскопічні немембранні органели загального призначення, які виконують роль цитоскелету, а також скоротливого апарату клітини. Являють собою тонкі волоконця діаметром близько 5 нм, які побудовані із скоротливих білків (актин, міозин, ін.). Вони розміщені переважно в кортикальній (підмембранній) зоні клітини та в складі її цитоплазматичних виростів. Функція -скоротлива-рухова. Існують також проміжні мікрофіламенти діаметром 10-15 нм. Білок, з якого вони побудовані, є суто специфічною ознакою клітин того чи іншого типу (наприклад, кератин є гістохімічним маркером клітин епітелію). Проміжні мікрофіламентии в основному відповідають за збереження клітиною своєї форми. В останні роки підтверджений їх зв'язок з регуляцією активності генома і процесами клітинної диференціації.
Мікротрубочки - субмікроскопічні немебранні органели загального призначення, які забезпечують рухливість органел клітини і входять до складу цитоскелету. Побудовані з білків тубулінів, молекули яких здатні до полімеризації і деполімеризації. Мікротрубочки є основою будови центросоми, веретена поділу, війок, джгутиків.
Включення, на відміну від органел, не є постійними структурними компонентами цитоплазми і не мають строго визначеної будови. Включеннями бувають переважно продукти клітинного метаболізму, які нагромаджуються у вигляді крапель, гранул, кристалів, вакуолей. За призначенням їх умовно поділяють на три групи: трофічні (жовток яйцеклітин, глікоген, краплі жиру), секреторні і спеціальні (гемоглобін).