Фізіологія людини - Вільям Ф. Ґанонґ 2002
Кровообіг
Природа серцевого скорочення й електричної активності серця
Виникнення і поширення імпульсів у серці
Анатомічні особливості
У серці людини СА-вузол міститься біля впадіння верхньої порожнистої вени у праве передсердя. АВ-вузол розміщений у правій задній ділянці міжпередсердної перегородки (рис. 28-1). Від СА-вузла відходять три пучки передсердних волокон типу волокон Пуркіньє, які сполучають СА-вузол з АВ-вузлом: передній міжвузловий пучок Бахмана, середній міжвузловий пучок Венкебахата задній міжвузловий пучок Тореля. Однак триває дискусія про те, які саме структури забезпечують передавання імпульсів від СА- до АВ-вузла: названі пучки чи типові передсердні кардіоміоцити. В нормі АВ-вузол - це єдиний провідний шлях між передсердями і шлуночками. АВ-вузол продовжений пучком Гіса. У верхній ділянці міжшлуночкової перегородки пучок Гіса розділений на праву і ліву ніжки. Ліва ніжка, відповідно, має передню і задню гілки. Ніжки та гілки йдуть субендокардіально з обох боків перегородки і переходять у волокна Пуркіньє, які поширюються до всіх ділянок міокарда шлуночків.
Гістологія серцевого м’яза описана у Розділі 3. Провідна система утворена атиповими кардіоміоцитами, які мають менше виражену посмугованість і нечіткі краї. СА-вузол та менше АВ-вузол містять також маленькі округлі клітини з малою кількістю органел, що з’єднані за допомогою щілинних контактів. Можливо, що це справжні пейсмекерні клітини, тому їх називають P-клітинами. М’язові волокна передсердь відокремлені від волокон шлуночків кільцем із фіброзної тканини. В нормі єдиною провідною тканиною між передсердями та шлуночками є пучок Гіса.
СА-вузол розвивається зі структур правого боку ембріона, а АВ-вузол - зі структур лівого боку. Тому в дорослих людей правий блукаючий нерв іннервує переважно СА-вузол, а лівий - АВ-вузол. Подібно до цього симпатичні нерви правого боку підходять до СА-вузла, а симпатичні нерви лівого боку - до АВ-вузла. З обох боків більшість симпатичних волокон відходить від зірчастого вузла. Норадренергічні волокна є епікардіальними, тоді як волокна блукаючого нерва - ендокардіальні. Однак є сполучення між парасимпатичними і симпатичними нервами серця, які забезпечують реципрокні гальмівні ефекти. Наприклад, ацетилхолін діє пресинаптично, пригнічуючи виділення норадреналіну симпатичними нервами; а нейропептид Y, що його виділяють норадренергічні нервові закінчення, може пригнічувати виділення ацетилхоліну.
Властивості серцевого м’яза
Електрична відповідь серцевого м’яза і тканини вузлів, а також переміщення йонів, які є в основі цієї відповіді, детально описано в Розділі 3. Волокна міокарда мають потенціал спокою близько -90 мВ (див. рис. 3-15). Залежність між потенціалом дії і скороченням зображено на рис. 3-14. Хоча волокна відокремлені мембранами, однак завдяки щілинним контактам деполяризація поширюється по них радіально, як у синцитії. Трансмембранний потенціал дії окремих кардіоміоцитів має фази швидкої деполяризації, плато і повільної реполяризації (див. рис. 3-15). Початкова деполяризація відбувається внаслідок надходження Na+ всередину клітини через швидкі Nа+-канали (Na+-струм, INa). Надходження Са2+ через повільні Са2+ - канали (Са2+-струм, ІСа) зумовлює фазу плато. Реполяризація спричинена виходом К+ через К+-канали трьох типів (див. Розділ 3). ЕКГ - це позаклітинно зареєстрована, сумована електрична активність усіх м’язових волокон серця. Співвідношення між потенціалами дії окремих ділянок серця та записом ЕКГ показано на рис. 28-1.
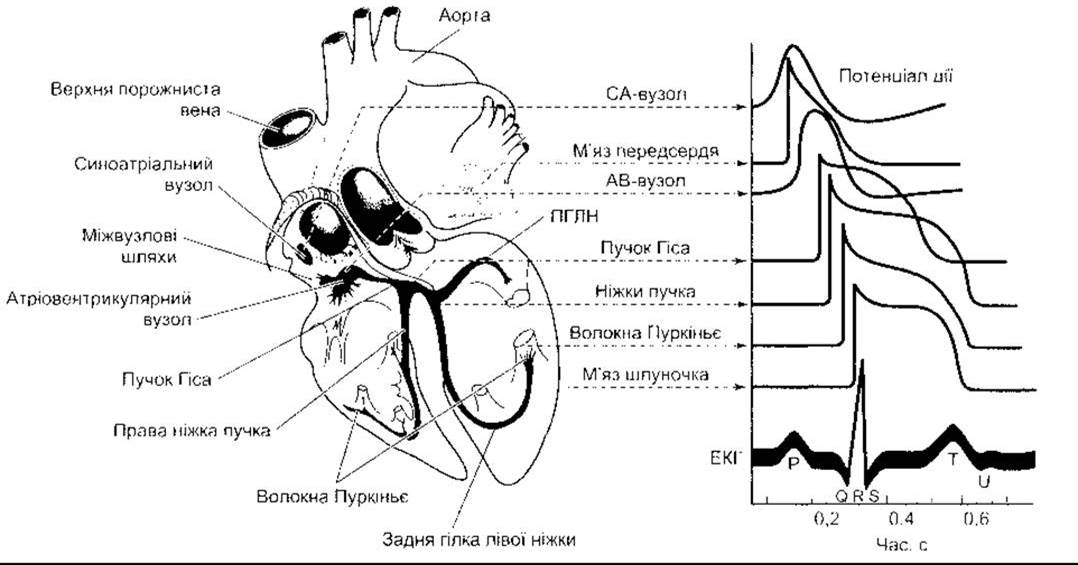
Рис. 28-1. Провідна система серця. Типові трансмембранні потенціали дії для СА- й АВ-вузлів та інших ділянок провідної системи, а також м’язів передсердь і шлуночків, співвіднесені з позаклітинно зареєстрованою електричною активністю, тобто електрокардіограмою (ЕКГ). Потенціали дії та ЕКГ зображено щодо однієї осі часу, проте з різними початками відліку на вертикальній осі; ПГЛН - передня гілка лівої ніжки.
Пейсмекерні потенціали
Після кожного імпульсу мембранний потенціал пейсмекерних клітин досягає порогового рівня. Отже, цей перед- потенціал, або пейсмекерний потенціал (рис. 28-2), зумовлює наступний імпульс. У момент піка кожного імпульсу виникає ІK, який спричинює реполяризацію. Потім ІK спадає, і зі зменшенням виходу К+ з клітин починається деполяризація мембрани, що зумовлює першу частину передпотенціалу. Далі відкриваються Са2+-канали. В серці розрізняють два типи цих каналів: Т-канали (від англ. transient - тимчасовий) і L-канали (від англ. Long-lasting -довготривалий). Кальцієвий струм ІСа завдяки відкриттю Т-каналів завершує передпотенціал, і ІСa) внаслідок відкриття L-каналів спричинює імпульс.
Потенціал дії в СА- і АВ-вузлах виникає головно завдяки надходженню Са2+, менш важливим є Na+. Унаслідок цього нема фази швидкої деполяризації, що характерна для інших ділянок провідної системи, а також для м’язів передсердь і шлуночків (див. рис. 28-1). У нормі передпотенціал простежується лише в СА- і АВ-вузлах. Однак є «латентні пейс- мекери» в інших ділянках провідної системи, які можуть виявити себе в умовах пригнічення СА- і АВ-вузлів або в разі порушення передавань імпульсів від них. М’язовим волокнам передсердь і шлуночків передпотенціал не притаманний, і вони здатні до спонтанної деполяризації лише тоді, коли є ушкодженими або неповноцінними.
У випадку стимулювання холінергічних волокон блукаючого нерва, які підходять до вузлів, мембрана гіперполяризується, і нахил передпотенціалу зменшується. Це пояснюють тим, що виділений нервовими закінченнями ацетилхолін підвищує К+-провідність вузлової тканини. Цей ефект відбувається через М2-мускаринові рецептори, які за участю ß-, у-субодиниць G-білка зумовлюють відкриття К+-каналів. Унаслідок виникає ІKAch, який протидіє зменшенню ІK. Крім того, активування М2-рецепторів знижує рівень цАМФ у клітинах, що веде до сповільненого відкриття Са2+-каналів, і, як наслідок, частота імпульсів зменшується. Значне стимулювання блукаючого нерва може на деякий час припинити спонтанне генерування імпульсів.
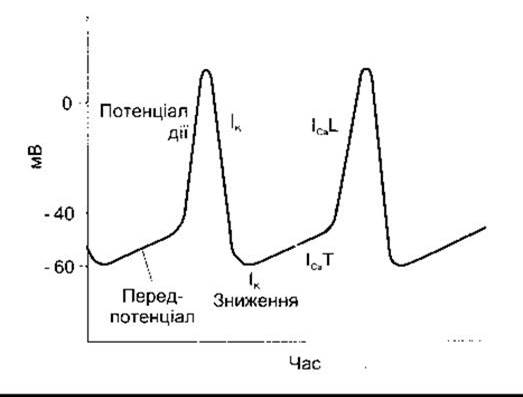
Рис. 28-2. Мембранний потенціал у пейсмекерній тканині Струм, що зумовлює кожну з фаз потенціалу, зазначено нижче або поряд; L-довготривалий; Т-тимчасовий. Зверніть увагу на те, що мембранний потенціал спокою пейсмекерноі тканини дещо нижчий, ніж у м’язах передсердь і шлуночків
Навпаки, стимулювання симпатичних нервів серця сприяє швидшому зменшенню мембранного потенціалу, і частота спонтанної генерації імпульсів збільшується (рис. 28-3). Виділений симпатичними нервовими закінченнями норадреналін зв’язується з ß1-рецепторами, унаслідок чого підвищується внутрішньоклітинний рівень цАМФ, відкриваються L-канали, збільшується ІСа і зменшується тривалість фази деполяризації потенціалу дії.
З урахуванням описаних вище особливостей іннервації серця стимулювання правого блукаючого нерва сповільнює частоту скорочення серця внаслідок пригнічення СА-вузла, тоді як стимулювання лівого блукаючого нерва головно сповільнює АВ-провідність. Подібно до цього, стимулювання правого зірчастого вузла прискорює роботу серця, а стимулювання лівого зірчастого вузла прискорює АВ- провідність і зменшує рефрактерність.
Частота генерації імпульсів у СА-вузлі та інших ділянках провідної системи може змінюватися під впливом температури і ліків. Вона збільшується з підвищенням температури, і це сприяє розвитку тахікардії в разі гарячки. Наперстянка пригнічує провідну систему і спричинює вплив, подібний до ефекту стимулювання блукаючого нерва (зокрема, на АВ-вузол).
Поширення збуджень у серці
Імпульси, що виникають у СА-вузлі, поширюються радіально через передсердя і сходяться в АВ-вузлі. Деполяризація передсердь завершується приблизно за 0,1 с. Оскільки передавання імпульсів через АВ-вузол повільніше (табл. 28-1), то є деяка затримка приблизно на 0,1 с (затримка в АВ-вузлі), перш ніж збудження пошириться на шлуночки. Ця затримка зменшується в разі стимулювання симпатичних нервів серця і збільшується у випадку стимулювання блукаючих нервів. Від вершини перегородки хвиля деполяризації з великою швидкістю поширюється волокнами Пуркіньє до всіх ділянок шлуночків (за 0,08-0,1 с). У людей деполяризація м’язів шлуночків розпочинається з лівого боку міжшлуночкової перегородки і спочатку поширюється праворуч через середню частину перегородки. Потім хвиля деполяризації поширюється вздовж перегородки до верхівки серця, а далі повертається вздовж стінок шлуночків до АВ-борозни, поширюючись з ендокардіальної поверхні до епікардіальної (рис. 28-4). Найнізніше імпульси проходять по задній стінці лівого шлуночка до основи серця, конуса легеневого стовбура і верхньої ділянки перегородки.
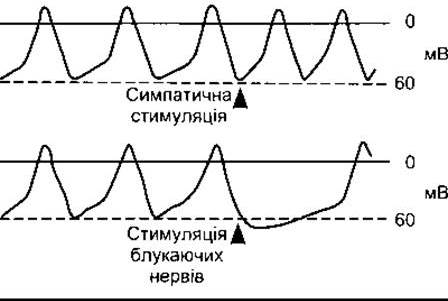
Рис. 28-3 Вплив стимулювання симпатичних (норадренер- гічних) і блукаючих (холінергічних) нервів на мембранний потенціал у СА-вузлі.