Фізіологія людини - Вільям Ф. Ґанонґ 2002
Фізіологія нервових і м'язових клітин
Збудлива тканина: м'язова
Скелетні м'язи - Властивості м'язів у інтактному організмі
Вплив денервації
У нормі в інтактних людей і тварин скелетні м’язи, якщо нема подразнення з боку рухових нервів, не скорочуються. Порушення іннервації спричинює атрофію м’язів, і, крім того, призводить до патологічної збудливості м’язової тканини та підвищення її чутливості до ацетилхоліну, що міститься в кровоплині (денерваційна гіперчутливість, див. Розділ 4). Унаслідок цього з’являються слабкі невпорядковані скорочення окремих м’язових волокон (фібриляції). Це класична ознака враження нижнього рухового нейрона. Якщо руховий нерв регенерує, то фібриляції припиняються. Як звичайно, такі скорочення не видно неозброєним оком; важливо не плутати їх з фасцикуляціями - видимими посіпуваннями груп м’язових волокон унаслідок патологічної імпульсації рухових нейронів спинного мозку.
Рухова одиниця
Аксон спинномозкового рухового нейрона розгалужується на декілька гілок, кожна з яких відходить до одного м’язового волокна. Отже, в нормі скорочення окремого м’язового волокна in vivo неможливе. Внаслідок подразнення одного рухового нейрона скорочується декілька волокон.
Сукупність рухового нейрона та волокон, які він іннервує, називають руховою одиницею. Кількість волокон, що припадає на одну рухову одиницю, різна і залежить від функції конкретного м’яза. В м’язах кисті та очного яблука, тобто тих, що забезпечують точні, висококоординовані рухи, одна рухова одиниця містить три-шість м’язових волокон. У литкових м’язах кота на одну рухову одиницю припадає 120-165 волокон, а великі м’язи спини у людини містять їх навіть більше.
Кожен спинномозковий руховий нейрон іннервує волокна лише одного типу, тому всі м’язові волокна рухової одиниці мають ідентичні характеристики. Залежно від типу м’язових волокон (див. табл. 3-3) і тривалості поодинокого скорочення рухові одиниці поділяють на швидкі та повільні. Повільні рухові одиниці містять невеликі за розміром нейрони із низькою швидкістю проходження імпульсу, а швидкі, відповідно, - великі нейрони із високою швидкістю проходження імпульсу (принцип розміру). У великих м’язах кінцівок під час виконання переважної більшості рухів повільні (малі) одиниці залучені першими, вони резистентні до втоми і фактично найчастіше застосовні. Швидкі одиниці, які легко і швидко втомлюються, як звичайно, беруть участь у рухах, що потребують значної сили м’язового скорочення.
Відмінності між різними типами рухових одиниць не лише генетично детерміновані, а й значно залежать від інших чинників, зокрема, від ступеня їхньої активності (див. нижче). Якщо нерв, що проходить до повільного м’яза, перерізати і на його місце імплантувати нерв, який до цього іннервував швидкий м’яз, то після регенерації пересадженого нерва він згодом почне виконувати іннервацію м’яза. Внаслідок цього м’яз поступово набуде характеристик швидкого з відповідними змінами ізоформ м’язових білків та активності міозинової АТФ-ази. Такі зміни відбуваються після перебудови профілю активності м’яза. В дослідах зі стимуляцією цих м’язів, лише модифікуючи профіль електричного стимулювання, можна спричинити зміни експресії генів та ізоформ ВЛМ.
Електроміографія
Активування рухових одиниць можна вивчати за допомогою електроміографії - реєстрації електричної активності м’яза, яка графічно відображається на екрані осцилоскопа. Це дослідження є абсолютно безболісним, його виконують, накладаючи на шкіру над досліджуваним м’язом невеликі металеві диски або вводячи підшкірні голкові електроди. Графічний запис змін електричної активності м’яза називають електроміограмою (ЕМГ). Типова ЕМГ зображена на рис. 3-12.
Фактори, що визначають м’язову активність
За допомогою електроміографії з’ясовано, що скелетні м’язи здорових осіб у стані спокою практично не виявляють спонтанної активності. У разі довільних рухів малої інтенсивності нервові імпульси надходять з невеликої кількості нейронів рухових одиниць, а зі збільшенням навантаження кількість задіяних рухових одиниць збільшується. Цей процес деколи називають залученням рухових одиниць. Отже, сила м’язової відповіді залежить від кількості активованих рухових одиниць. Крім того, значну роль відіграє частота нервових імпульсів, адже напруження, що розвивається під час тетанічного скорочення, є набагато більшим, ніж під час поодиноких скорочень. Певним визначним фактором є й довжина м’яза. І, зрештою, нервові імпульси в рухових одиницях утворюються асинхронно, тобто із зсувом за фазою один щодо іншого. Внаслідок асинхронності роботи нейронів відповіді окремих м’язових волокон зливаються у плавне суцільне скорочення м’яза.
Сила скелетних м’язів
Скелетні м’язи людини можуть створювати напруження 3-4 кг на 1 см2 площі перерізу. Цей показник ідентичний до того, який одержують у багатьох дослідах на тваринах, і є, ймовірно, сталим для усіх видів ссавців. Оскільки багато м’язів у людини мають порівняно велику площу перерізу, то вони можуть розвивати досить значне напруження.
Наприклад, литковий м’яз не лише підтримує масу всього тіла під час пересування угору, а й витримує в декілька разів більше навантаження, коли стопа стикається з поверхнею землі під час бігу або стрибків. Ще більше вражає приклад із великим сідничним м’язом, який може розвивати силу до 1200 кг. Сила, яку б розвинули всі м’язи тіла здорової людини, якби скоротились одночасно, дорівнює приблизно 22 т.
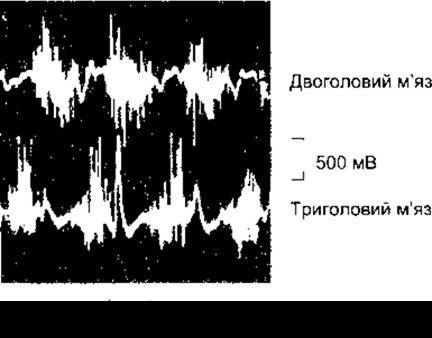
Рис. 3-12. Електроміографічний запис біопотенціалів двоголового та триголового м’язів людини під час почергового згинання та розгинання у ліктьовому суглобі (за ВС Garoutte).
Роль м’язів у підтриманні пози і руху
Будь-які рухи відбуваються із максимальним використанням описаних вище фізіологічних принципів. Наприклад, точки прикріплення м’язів на скелеті розташовані так, що більшість з них на початку скорочення мають довжину спокою.
У м’язах, що поширюються більше ніж на один суглоб, рухи в одному суглобі можуть компенсувати рухи в іншому, завдяки чому під час скорочення м’яза його довжина лише незначно змінюється. Таке скорочення, що є майже ізометричним, дає змогу розвинути максимальне напруження. Зокрема, кравецькі м’язи проходять з таза через кульшовий і колінний суглоби до велико- та малогомілкової кісток. Скорочення цього м’яза спричинює згинання стегна.
У разі згинання стегна в кульшовому суглобі подовження частини кравецького м’яза, що проходить через кульшовий суглоб, компенсує скорочення тієї його частини, що проходить через колінний суглоб. Під час складних рухів із залученням великої кількості м’язів їхня взаємна координація відбувається так, щоб забезпечити оптимальні умови скорочення для принаймні деяких з них. Точки прикладання м’язової сили часто так розташовані щодо суглобів, що утворюють важелі, за допомогою яких за мінімального напруження м’яза створюється максимальна сила. Це важливо не лише для самих м’язів, а й для кісток та сухожилків. Скорочення м’язів організовані так, що навантаження на кістки та суглобово-зв’язковий апарат зрідка перевищує 50% їхньої міцності, а це захищає їх від пошкодження.
Під час ходіння кожна кінцівка ритмічно проходить через фазу опори, коли стопа дотикається землі, і фазу підіймання, коли стопа перебуває у повітрі. Фази опори обох ніг накладаються, отже, під час кожного циклу ходи є два періоди подвійної опори.
На початку кожного кроку у м’язах-згиначах ноги простежується короткочасне стрибкоподібне підвищення напруження, а потім нога переходить до періоду із незначним зростанням напруження. Отже, під час кожного кроку м’язи перебувають в активному стані лише упродовж короткого проміжку часу, тому тривале ходіння викликає лише порівняно незначну втому.
Здорова молода особа під час ходіння в зручному для неї темпі рухається зі швидкістю близько 80 м/хв, у цьому разі виникає потужність 150-175 Вт.
У дослідженнях, які проводили з молодими волонтерами, їх просили ходити у найзручнішому для них темпі. Виявилось, що всі вони обрали швидкість близько 80 м/хв, за якої витрати енергії мінімальні. Ходіння із швидкістю, що більша або менша від оптимальної, потребує більше енергії.
Незважаючи на те, що ходіння є складним рефлекторно-руховим актом, воно відбувається більш-менш автоматично. За допомогою дослідів на тваринах з’ясовано, що такий процес запрограмований у певним чином організованих провідних шляхах спинного мозку і, відповідно, активований конкретними командними сигналами. Це досить подібне до процесу ініціювання профілів активності внаслідок імпульсації командних нейронів у безхребетних тварин.
Патологія м’язів
Як уже зазначено, мутації генів, що кодують різноманітні компоненти дистрофін-глікопротеїнового комплексу, спричинюють м’язову дистрофію - синдром, що супроводжується м’язовою слабкістю, яка прогресує. Деякі з багатьох форм цього захворювання можуть зумовлювати стійку інвалідність або смерть. М’язову дистрофію Дюшена, пов’язану з Х-хромосомою, виявляють в одного з близько 3000 новонароджених хлопчиків, як звичайно, вона призводить до смерті у віці до 30 років. Спричинена ця дистрофія мутаціями гена дистрофіну, внаслідок яких дистрофіну в м’язах зовсім нема. У випадку легшої форми цього захворювання - м’язової дистрофії Бекера, - дистрофін у м’язах наявний, проте він модифікований або міститься у недостатній кількості. Різноманітні дистрофії м’язів пояса кінцівок пов’язані із мутацією генів, які кодують саркоглікани.
У ділянках нервово-м’язових з’єднань дистрофін замінений утрофіном - подібним за структурою білком, що кодований іншим геном. У генетично модифікованих мишей із дефіцитом дистрофіну, у яких утрофін продукований у надлишкових кількостях, простежується значне поліпшення м’язової функції; а це доводить, що дистрофін значно заміщений утрофіном. Тому сьогодні триває активний пошук медикаментозних або інших засобів стимулювання гена утрофіну у людей, що хворіють на м’язову дистрофію Дюшена.
Мутації генів, що кодують ензими, які беруть участь у метаболізмі вуглеводів, жирів, білків та утворенні АТФ у м’язовій тканині, спричинюють метаболічні міопатії. Одна з них - синдром Мак-Кардля, що описаний у Розділі 17.
Метаболічні міопатії мають різноманітні прояви залежно від конкретних генетичних порушень, однак їхньою загальною ознакою є швидка втомлюваність м’язів і, ймовірно, розпад м’язової тканини внаслідок накопичення токсичних продуктів обміну.
У разі різноманітних форм міотонії збільшується тривалість розслаблення м’язів після їхнього довільного скорочення. Міотонії спричинені аномаліями генів, що містяться в 7, 17 та 19 хромосомах і призводять до порушення структури Na+- або Сl- -каналів.
Сьогодні тривають інтенсивні дослідження щодо можливостей імплантації нормальних генів, які відповідають за розвиток м’язової тканини у хворих на м’язові дистрофії, проте в цьому напрямі ще багато невирішених проблем.
Розвиток м’язів
Хоча розгляд ембріогенезу м’язів не є метою цієї книги, та варто зазначити, що останніми роками з’ясовано чимало раніше невідомих механізмів генетичного контролювання розвитку м’язів.
Провідну роль у цьому процесі відіграє білок міогенін, що є фактором транскрипції відповідних генів. Під його впливом фібробласти перетворюються на м’язові клітини. Піддослідні миші, гомозиготні за мутантним геном міогеніном, помирають невдовзі після народження внаслідок недостатнього розвитку м’язів, у тому числі дихальної мускулатури.