Фізіологія людини - Вільям Ф. Ґанонґ 2002
Кровообіг
Кровообіг в окремих ділянках організму
Церебральний кровообіг - Гематоенцефалічний бар'єр
Наявність щільних контактів між ендотеліальними клітинами капілярів мозку та епітеліальними клітинами судинного сплетення ефективно запобігає проникненню білків у мозок дорослих та сповільнює пенетрацію менших за розміром молекул. Прикладом може бути повільне проникнення сечовини (рис. 32-5). Рівень проходження молекул обернено пропорційний до її розміру та прямо пропорційний їхній розчинності у жирах. Цей унікальний обмежений обмін речовин називають гематоенцефалічним бар’єром. Деякі фізіологи використовують цей термін лише для опису бар’єру в капілярній стінці, тоді як для опису бар’єру у судинному сплетенні застосовують термін гемато-СМР бар’єр. Та оскільки обидва бар’єри мають багато спільного, то доцільно використовувати один термін до обох структур. Крім того, незважаючи на те, що везикулярне транспортування через мозкові капіляри порівняно невелике, є багато транспортних систем із залученням носіїв. Отже, говорячи про ступінь проходження будь-якої речовини через гематоенцефалічний бар’єр, мають на увазі проникнення через описані бар’єри і транспортування за допомогою систем ендотеліальних клітин.
Проходження речовин з головного мозку у кров відрізняється від руху у протилежному напрямі і є набагато вільнішим завдяки основному потоку СМР у венозне русло через ворсинки павутинної оболонки (див. вище). До того ж, у стінці капілярів є трансферні системи, опосередковані носіями, які забезпечують переважний рух речовин з мозку у кров.
Проникнення речовин у головний мозок
Вода, СО2 та О2 легко проникають у мозок. Те ж можна сказати про вільні жиророзчинні сполуки та стероїдні гормони, тоді як їхні форми, зв’язані з білками, та загалом усі білки і поліпептиди не можуть туди проникнути. Легка пенетрація СО2 протистоїть повільному проникненню Н+ та HCО3-, що має фізіологічне значення для регулювання дихання (див. Розділ 36).
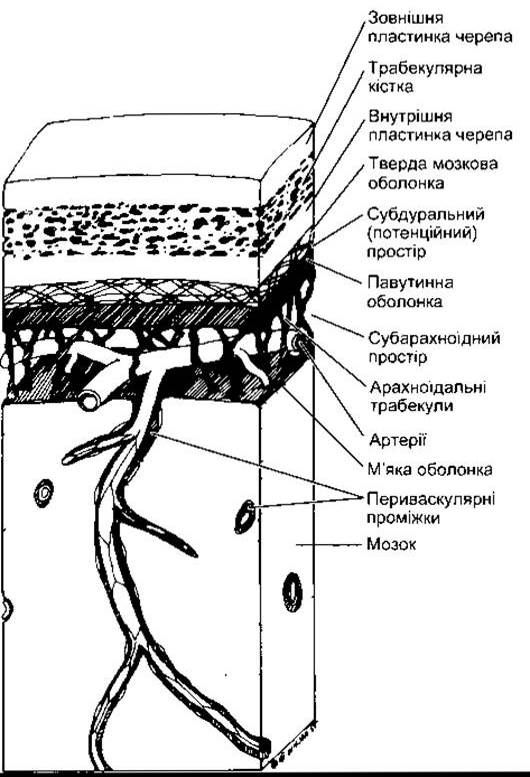
Рис. 32-4. Розміщення оболонок головного мозку та їхнє співвідношення з черепом і тканиною мозку (відтворено за дозволом з Wheater PR et al: Functional Histology. Churchill Livingstone, 1979).
Глюкоза є головним джерелом енергії для нервових клітин. Вона повільно проходить через гематоенцефалічний бар’єр, водночас її транспортує через капілярні стінки транспортна молекула GLUT 1. У головному мозку міститься дві молекули GLUT 1: GLUT 1 55 К та GLUT 1 45 К. Обидві вони кодовані тим самим геном, однак відрізняються за ступенем глюкозування. GLUT 1 55 К є у високій концентрації в мозкових капілярах (рис. 32-6). У дітей з вродженою недостатністю GLUT 1 простежується низький рівень глюкози в СМР за нормального її рівня в плазмі, що супроводжується судомами та сповільненим розвитком.
Іншою транспортною системою у мозкових капілярах є унікальні Nа+-К+-2Сl--котранспортери, стимульовані ЕТ-1 і ЕТ-3 та, імовірно, індуковані гуморальними факторами астроцитів. Це допомагає підтримувати концентрацію К+ у мозку на низькому рівні. Крім того, є транспортери для тиреоїдних гормонів, декількох органічних кислот, холіну, попередників нуклеїнових кислот та нейтральних, лужних і кислих амінокислот.
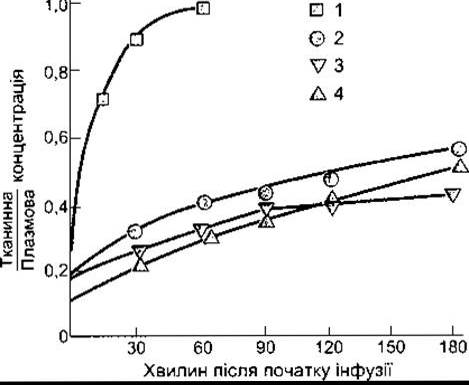
Рис. 32-5. Проникнення сечовини у м’яз (1), головний мозок (2), спинний мозок (3) та спинномозкову рідину (4). Сечовину вводили шляхом постійної інфузії (модифіковано та відтворено за дозволом з Kleeman CR, Davson Н, Levin Е: Urea transport in the central nervous system. Am J Physiol 1962;203: 739).
Навколошлуночкові органи
Після введення у плазму білків, зв’язаних з барвниками, забарвлюється багато тканин, однак речовина головного мозку не є забарвленою. Це спостереження, зроблене багато років тому, стало основою для створення концепції гематоенцефалічного бар’єру. Однак чотири невеликі ділянки у стовбурі мозку або біля нього забарвлюються так само, як і тканини поза мозком. До них належать задня частка гіпофіза (нейрогіпофіз) та прилегла вентральна частина серединного горба гіпоталамуса, задня ділянка, судинний орган кінцевої пластинки (СОКП, супраоптичний перехрест) та субфорнікальний орган (СФО).
Ці відділи разом утворюють навколошлуночкові органи (рис. 32-7). Всі вони містять фенестровані капіляри і з огляду на їхню проникність уважають, що вони перебувають «поза гематоенцефалічним бар’єром». Деякі з них діють як нейрогемальні органи, тобто ділянки, де поліпептиди, секретовані нейронами, потрапляють у кровоплин. Наприклад, окситоцин та вазопресин виділяються у системний кровообіг у задній частці гіпофіза, а гіпофізотропні гормони гіпоталамуса секретують у портальну гіпофізарну систему у серединному горбі. Інші навколошлуночкові органи містять рецептори для різних пептидів та інших речовин і діють як хеморецепторні зони, тобто ділянки, де речовини із крові, що циркулює, можуть зумовити зміни функції мозку, обминаючи гематоенцефалічний бар’єр. Задня ділянка є хеморецепторною зоною, що спричинює блювання у відповідь на зміни хімічного складу плазми (див. Розділ 14). Вона також пов’язана з регулюванням серцево-судинної системи, і у багатьох випадках циркулювальний ангіотензин II діє на задню ділянку, зумовлюючи підвищення артеріального тиску. Ангіотензин II діє також на СФО та, можливо, на СОКП, посилюючи споживання води. Крім того, виявилось, що СОКП є ділянкою розміщення осморецепторів, які контролюють секрецію вазопресину (див. Розділ 14); є дані про те що ІЛ-1 крові спричинює гарячку, впливаючи на навколошлуночкові органи.
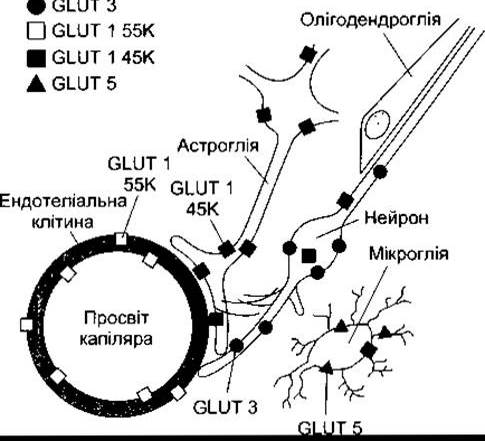
Рис. 32-6. Розміщення різних транспортерів GLUT у головному мозку (відтворено за дозволом з Maher F, Vannucci SJ, Simpson IA: Glucose transporter proteins in brain. FASEB J 1994;8:1003).
Субкомісуральний орган (див. рис. 32-7), щільно зв’язаний з епіфізом, за гістологічною будовою нагадує навколошлуночкові органи. Однак у ньому нема фенестрованих капілярів, не зафіксовано високої проникності, і основна функція не визначена. Шишкоподібна залоза та передній гіпофіз містять фенестровані капіляри та перебувають поза гематоенцефалічним бар’єром, однак обидва органи належать до ендокринних залоз і не є частиною головного мозку.
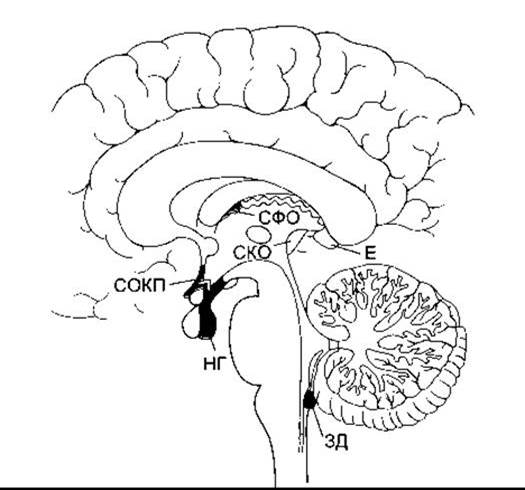
Рис. 32-7. Навколошлуночкові органи. Нейрогіпофіз (НГ); судинний орган кінцевої пластинки (СОКП, супраоптичний перехрест), субфорнікальний орган (СФО), та задня ділянка (ЗД) показані у саґітальному перерізі мозку людини; СКО - субкомісуральний орган; Е - епіфіз.
Функція гематоенцефалічного бар’єру
Гематоенцефалічний бар’єр, імовірно, забезпечує стабільність мікрооточення нейронів у центральній нервовій системі. Ці нейрони настільки залежать від концентрації R+, Са2+, Mg2+, Н+ та інших йонів у рідині, яка оточує їх, що навіть незначні зміни ведуть до серйозних наслідків. Сталий склад ПКР у всіх частинах тіла підтримують множинні гомеостатичні механізми (див. Розділи 1 та 39), однак з огляду на чутливість кіркових нейронів до змін йонного складу немає нічого дивного у наявності додаткового захисного механізму для їхнього збереження. Аналогічно, гематотестикулярний бар’єр захищає склад рідини, що оточує гермінативний епітелій, а плацентарний бар’єр - склад тканинної рідини плоду.
До інших можливих функцій гематоенцефалічного бар’єру можна зачислити захист мозку від ендо- та екзогенних токсинів у крові та запобігання потраплянню нейротрансмітерів у загальний кровообіг.
Розвиток гематоенцефалічного бар’єру
В експериментальних тварин багато дрібних молекул проникають у мозок легше в пери- та неонатальний періоди, ніж у зрілої особини. На цьому ґрунтується твердження про те, що гематоенцефалічний бар’єр у новонародженого є незрілим. Люди під час народження є зрілішими, ніж щурі та інші види експериментальних тварин, проте точних даних про пасивну проникність гематоенцефалічного бар’єру у людини немає. Однак у дітей з вираженою жовтяницею, високим рівнем вільного білірубіну у плазмі та незрілою системою кон’югації білірубіну у печінці вільний білірубін проникає в мозок і за наявності асфіксії вражає базальні ганглії (керніктерус, ядерна жовтяниця). Еквівалентом цього стану у пізнішому періоді життя є синдром Криглера-Найяра, за якого простежується вроджена недостатність глюкуронілтрансферази. У таких осіб може бути дуже високий рівень вільного білірубіну у крові з наступним розвитком енцефалопатії. У разі інших станів рівень вільного білірубіну недостатньо високий для спричинення ушкоджень тканини мозку.
Клінічне значення
Лікарі повинні знати ступінь проникнення ліків у головний мозок для того, щоб лікувати захворювання нервової системи грамотно. Проникнення медикаментів, як і інших речовин, залежить від багатьох факторів. Наприклад, деякі антибіотики проникають у мозок дуже обмежено, тоді як інші - досить легко. З клінічних спостережень відомо, що аміни дофамін та серотонін проникають у тканину мозку незначно, а їхні кислотні попередники - L-ДОФА та 5-гідрокситриптофан - порівняно легко (див. Розділи 12 та 15).
Інше важливе клінічне спостереження таке: гематоенцефалічний бар’єр може руйнуватись у зонах інфекційного та іншого ушкодження. Пухлинний ріст супроводжується утворенням нових судин, і новоутворені капіляри не мають контактів з нормальними астроцитами. Тому не формуються щільні контакти, і судини можуть бути навіть фенестрованими. Відсутність бар’єру допомагає визначити локалізацію пухлини; такі сполуки, як значений йодом альбумін, проникають у нормальну тканину мозку дуже повільно, а у пухлинну - швидко. Тому пухлина є ніби «островом радіоактивності» серед навколишньої нормальної тканини мозку. Гематоенцефалічний бар’єр може бути також тимчасово зруйнований у разі раптового значного підвищення артеріального тиску або внутрішньовенного введення гіпертонічних розчинів.