Фізіологія людини - Вільям Ф. Ґанонґ 2002
Функції нервової системи
Рефлекси
Моносинаптичні рефлекси: рефлекс розтягування
Розтягнутий скелетний м’яз без порушень іннервації скорочується. Цю реакцію називають рефлексом розтягування (міотатичним). Подразником, що спричинює цей рефлекс, є розтягування м’яза, а відповіддю - скорочення розтягнутого м’яза. Функцію чутливого рецептора у цьому разі виконує м’язове веретено. Імпульси, що виникають у м’язовому веретені, проходять до ЦНС по швидких чутливих волокнах безпосередньо до мотонейронів, які іннервують цей м’яз. Нейротрансмітером у центральних синапсах є глютамат. Рефлекси розтягування - найвідоміші і найліпше вивчені моносинаптичні рефлекси організму.
Клінічні приклади
Ударяння молоточком по сухожилку нижче від колінної чашечки спричинює колінний рефлекс - рефлекс розтягування чотириголового м’яза стегна, оскільки вдаряння по його сухожилку приводить до скорочення м’яза. Подібне скорочення можна спостерігати і в разі розтягування чотириголового м’яза руками. Рефлекси розтягування можна спровокувати також у більшості інших великих м’язів тіла. Вдаряння по сухожилку триголового м’яза плеча, наприклад, спричинює розгинання в лікті, що виникає внаслідок скорочення цього м’яза; вдаряння по сухожилку Ахілла спричинює рефлекс розтягування, зумовлений скороченням литкового м’яза; а вдарянням збоку обличчя зумовлює рефлекс розтягування в жувальному м’язі. Інші рефлекси розтягування описані в підручниках з неврології.
Структура м’язового веретена
Кожне м’язове веретено містить приблизно 10 м’язових волокон, охоплених сполучнотканинною капсулою. Ці волокна більше наближені за структурою до волокон ембріонального типу і менше посмуговані, ніж інші волокна м’яза. їх називають внутрішньоверетенними волокнами, на відміну від зовнішньоверетенних волокон, що є регулярними скоротливими одиницями м’яза. Внутрішньоверетенні волокна розміщені паралельно щодо інших волокон м’яза, оскільки кінці капсули веретена прикріплені до сухожилків на кожному з кінців м’яза або з боків зовнішньоверетенних волокон.
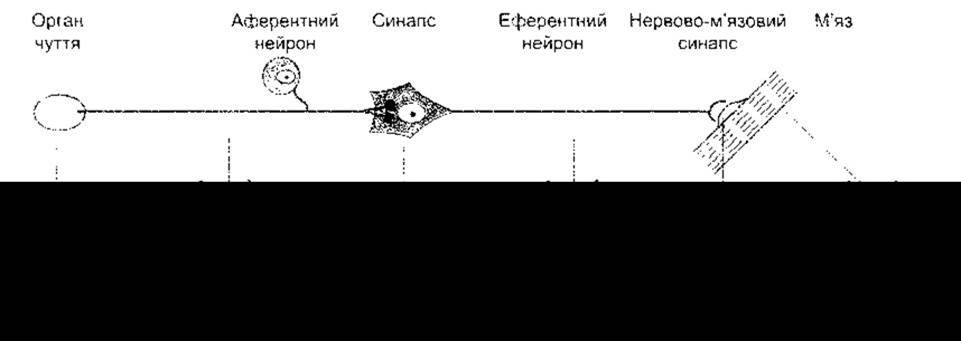
Рис. 6-1. Рефлекторна дуга. Зауважте, що в рецепторі та ЦНС виникає непоширювальна градуйована реакція. Вона пропорційна до сили подразника. Реакція в нервово-м’язовому синапсі теж Градуйована, тому за нормальних умов вона достатня, щоб спричинити реакцію скелетного м’яза. З іншого боку, в частинах рефлекторної дуги, призначених для проведення імпульсів (аферентний та еферентний нейрони, м’язова мембрана), потенціали дії виникають за принципом «усе або нічого».
У м’язових веретенах ссавців є два типи внутрішньоверетенних волокон. Один тип волокон містить у розширеній центральній ділянці багато ядер, тому його називають волокном ядерної сумки (рис. 6-2). Переважно у веретені містяться два волокна ядерної сумки: волокно ядерної сумки 1 з низьким рівнем активності міозин АТФ-ази і волокно ядерної сумки 2 з високим рівнем активності міозин АТФ-ази. Інший тип волокон - волокна ядерного ланцюжка - тонші і коротші, не мають окресленої сумки. Таких волокон у веретені чотири або й більше, їхні кінці прикріплені з боків до волокон ядерної сумки. Кінці внутрішньоверетенних волокон скоротливі, тоді як їхні центральні частини, вірогідно, не скорочуються.
У кожному веретені є два типи чутливих закінчень. Первинні (кільцево-спіральні) закінчення - це закінчення швидкопровідних аферентних волокон групи Іа (див. табл. 2-2). Одна гілка волокна Іа іннервує волокно ядерної сумки 1, тоді як інша - волокна ядерної сумки 2 і волокна ядерного ланцюжка. Ці чутливі волокна спіраллю оточують центральні ділянки волокон ядерної сумки і волокон ядерного ланцюжка. Вторинні (гроноподібні) закінчення є закінченнями чутливих волокон групи II. Вони розміщені поблизу кінців внутрішньоверетенних волокон ядерного ланцюжка.
Веретена мають власну рухову іннервацію. Ці нерви, 3-6 мкм у діаметрі, становлять приблизно 30% волокон переднього корінця і належать до групи Ерлянґера і Ґассера А у. Завдяки такому розміру їх названо у-еферентами Лекселя, або малою руховою нервовою системою. Ці волокна прямують винятково до веретен. Крім того, як внутрішньо-, так і зовнішньоверетенні волокна іннервовані більшими ß-мотонейронами. Закінчення еферентних волокон бувають двох гістологічних типів: рухові кінцеві пластинки (пластинчасті закінчення) волокон ядерної сумки і закінчення у вигляді сіток (протяжні закінчення), що містяться головно на волокнах ядерного ланцюжка.
У веретенах є два типи чутливих волокон - динамічні та статичні (див. нижче), а у- та ß-мотонейрони забезпечують два функційні типи відповідей. Подразнення першого типу посилює динамічне реагування (динамічні веретено-моторні аксони), а подразнення другого типу - статичне за сталої довжини (статичні веретеноподібні аксони).
Центральні зв’язки аферентних волокон
За допомогою експерименту можна з’ясувати таке: волокна Іа, що прямують від первинних закінчень м’яза, закінчуються безпосередньо на мотонейронах, які іннервують зовнішньоверетенні волокна цього ж м’яза. Час, що минає між початком подразнення і відповіддю, називають часом рефлексу. В організмі людини час рефлексу розтягування, наприклад колінного рефлексу, становить 19-24 мс. Слабке подразнення чутливого нерва, який відходить від м’яза, що, як відомо, призводить до збудження лише волокон Іа, спричинює скорочувальну реакцію, з такою ж латентністю. Оскільки швидкість передавання імпульсів в аферентних та еферентних нейронах відома, а відстань від м’яза до спинного мозку легко виміряти, то можна обчислити частку часу реакції, потрібну для передавання збудження від м’яза до спинного мозку і в зворотному напрямі. Якщо це значення відняти від загального часу реакції, то залишок, який називають центральною затримкою, відповідатиме часу, необхідному для рефлекторної активності в межах спинного мозку. В організмі людини центральна затримка розвитку колінного рефлексу становить 0,6-0,9 мс. Подібні значення отримані також в експериментах на тваринах. Оскільки мінімальна синаптична затримка становить 0,5 мс (див. Розділ 4), то зрозуміло, що рефлекторна дуга такого типу рефлексу містить лише один синаптичний контакт.
Збудження від м’язових веретен, яке спричинює м’язове скорочення, поширюється полісинаптичними шляхами, і воно, ймовірно, надходить від вторинних закінчень. Волокна групи II, що утворюють моносинаптичні контакти з мотонейронами, роблять невеликий внесок у рефлекси розтягування.
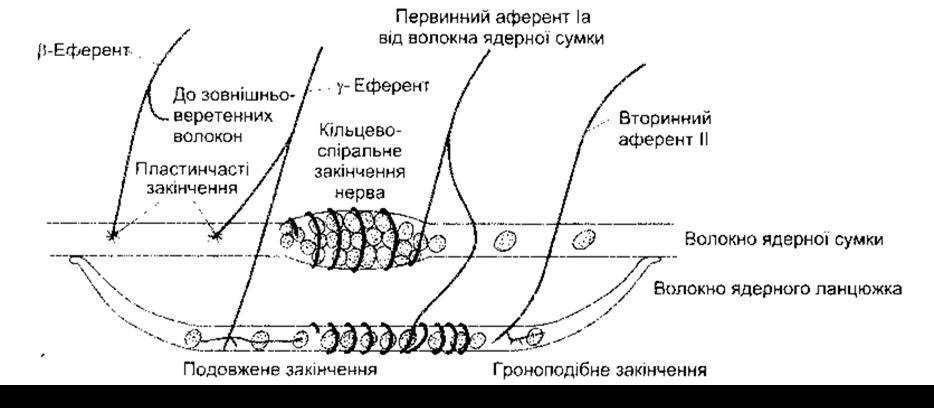
Рис. 6-2. Схематичне зображення головних компонентів м’язового веретена ссавців. Кожне веретено охоплене капсулою і містить два волокна ядерної сумки та чотири або більше волокна ядерного ланцюжка.
Функція м’язових веретен
Під час розтягування м’язового веретена його чутливі закінчення деформуються, і в них генеруються рецепторні потенціали. Це, відповідно, зумовлює виникнення потенціалів дії в чутливих волокнах, що за частотою пропорційні до ступеня розтягування. Веретена розміщені паралельно щодо зовнішньоверетенних волокон, і коли м’яз пасивно розтягується, то вони теж підлягають розтягуванню. Внаслідок цього виникає скорочення зовнішньоверетенних волокон м’яза. З іншого боку, аференти веретена характерним способом припиняють збудження, якщо м’яз скорочується внаслідок електричного подразнення нервових волокон, що надходять до зовнішньоверетенних волокон, оскільки тоді м’яз скорочується, а веретено - ні (рис. 6-3).
Отже, веретено і його рефлекторні зв’язки є механізмами зворотного зв’язку, що діє зі збільшенням довжини м’яза. Коли м’яз розтягується, то зростає збудження у веретені і виникає рефлекторне скорочення м’яза. З іншого боку, коли м’яз скорочується без зміни збудження в еферентах, то збудження у веретені зменшується, і м’яз розслаблюється.
У разі розтягування веретена подразнюються первинні закінчення волокон як ядерної сумки, так і ядерного ланцюжка, проте тип відповіді в цьому випадку відрізняється. Нерви, що відходять від закінчень ділянки ядерної сумки, зумовлюють динамічну реакцію, тобто імпульсація в них відбувається швидше, коли м’яз розтягнутий, і повільніше, якщо розтягування тривале (рис. 6-4). Нерви, що відходять від первинних закінчень волокон ядерного ланцюжка, створюють статичну реакцію, тобто імпульсація в них відбувається з високою швидкістю протягом усього періоду, доки м’яз розтягнутий. Отже, первинні закінчення реагують на зміну як довжини м’яза, так і ступеня його розтягування. Реакція первинних закінчень на фазові та статичні явища, що відбуваються в м’язі, має важливе значення, оскільки швидке фазове реагування допомагає зменшувати амплітуду осциляцій, спричинених затримками проведення в дузі зворотного зв’язку, що регулює довжину м’яза. Невеликі осциляції в дузі зворотного зв’язку, звичайно ж, виникають. Цей фізіологічний тремор відбувається з частотою приблизно 10 Гц. Проте тремор був би сильнішим, якби не змінювалась чутливість веретена до швидкості розтягування.
Ефект референтної імпульсації
Подразнення у-еферентної системи зумовлює дуже різноманітні явища, що виникають під час скорочення зовнішньоверетенних волокон. Воно не призводить безпосередньо до помітного скорочення м’язів, оскільки внутрішньоверетенні волокна недостатньо потужні, кількість їх недостатня щоб спричинити скорочення. Проте у-еференти скорочують скоротливі кінці внутрішньоверетенних волокон, що зумовлює розтягування ділянки ядерної сумки веретен, деформуючи кільцево-спіральні закінчення й ініціюючи імпульси у волокнах Іа. Це, відповідно, може призводити до рефлекторного скорочення м’яза. Отже, м’яз може скорочуватись унаслідок подразнення а-мотонейронів, що іннервують зовнішньоверетенні волокна, або у-еферентних нейронів, що ініціюють м’язове скорочення опосередковано - шляхом рефлексу розтягування.
З посиленням у-еферентної імпульсації, внутрішньоверетенні волокна стають коротшими від зовнішньоверетенних. Коли ж цілий м’яз розтягується під час подразнення у-еферентів, то внаслідок додаткового розтягування ділянки ядерної сумки генеруються додаткові потенціали дії, і кількість імпульсів у волокнах Іа далі збільшується (див. рис. 6-3). Посилення у-еферентної імпульсації, отже, приводить до зростання чутливості веретена, а чутливість веретен до розтягування змінюється залежно від ступеня у-еферентної імпульсації. Є значна вірогідність посилення у-еферентної імпульсації одночасно з посиленням імпульсації а-мотонейронів, що ініціює рухи. Внаслідок цього а-зв’язку веретено скорочується разом з м’язом, і імпульсація веретена триває протягом скорочення. Отже, веретено зберігає здатність відповідати на розтягування і рефлекторно регулювати рівень збудження в мотонейронах упродовж скорочення.
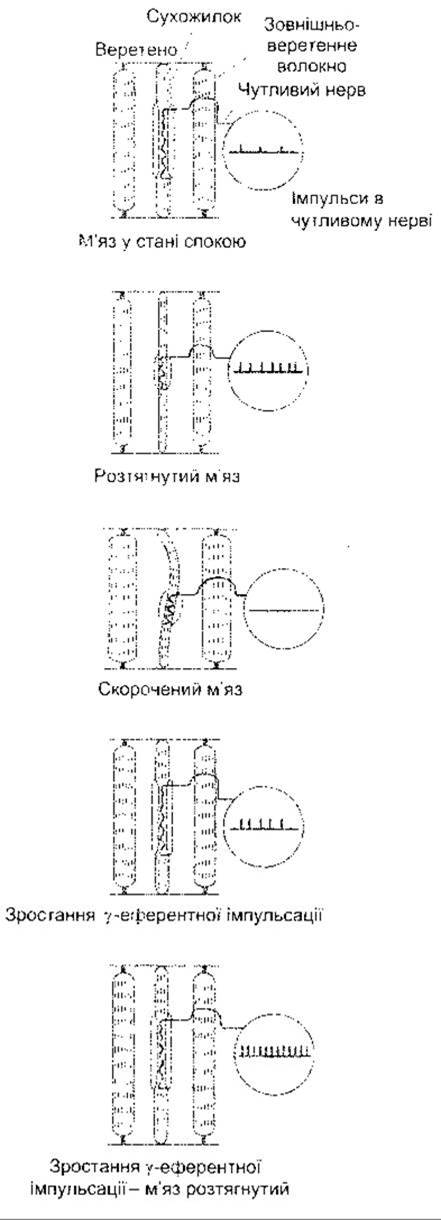
Рис. 6-3. Вплив різних умов на імпульсацію в м’язовому веретені.
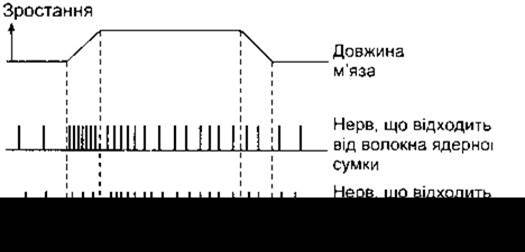
Рис. 6-4. Реакція аферентів веретена у відповідь на розтягування м’яза. Дві нижні лінії відображають кількість збудження в аферентних нервах, що відходять від первинних закінчення волокон ядерної сумки і ядерного ланцюжка, за умов, колі м’яз розтягнутий, а також коли він набуває попередньої ДОВжини.
Динамічні та статичні у- і ß-еференти описані раніше. Подразнення динамічних еферентів зумовлює зростання чутливості веретена до швидкості зміни довжини м’яза пі, час розтягування, а статичних - зростання чутливості веретена у разі стійкого тривалого розтягування. Отже, є змог роздільно регулювати реакції, що виникають у веретені відповідь на фазові або статичні явища.
Регулювання у-еферентної імпульсації
Рухові нейрони у-еферентної системи регульовані них хідними шляхами, що відходять з багатьох ділянок ГОЛОВного мозку. За допомогою цих шляхів може відбуватися регулювання чутливості м’язових веретен і, відповідне поріг рефлексів розтягування багатьох ділянок тіла, що важливо для постави (див. Розділ 12).
Інші чинники теж впливають на у-еферентну імпульсецію. Хвилювання призводить до посилення імпульсації цей факт, імовірно, пояснює гіперактивність сухожилкови рефлексів, що інколи простежуються в схвильованих пацієнтів. Крім того, з посиленням еферентної імпульсації пов’язані також несподівані рухи. Подразнення шкіри зокрема, небезпечними агентами, призводить до посилення еферентної імпульсації у веретенах іпсилатеральних м’язів згиначів та її послаблення у веретенах м’язів-розгиначів, а також виникнення протилежних явищ в іншій кінцівці. Добре відомо, що намагання розвести руки в разі зчеплених пальців обидвох кистей посилює колінний рефлекс (феномен Єндрасіка), і це теж може бути зумовлене посилення у-еферентної імпульсації, ініційованої аферентним імпульсами, що надходять від ділянок кистей.
Реципрокна іннервація
Під час рефлексу розтягування протилежні за функціє] м’язи щодо м’язів, які рефлексують, тобто їхні антагоністі розслаблюються. Цей феномен зумовлює реципрокни іннервація. Імпульси, що надходять по волокнах Іа від м’язових веретен м’язів-агоністів, спричинюють постсинаптичне гальмування в мотонейронах, які проходять до антагоністів. Шлях, що опосередковує цей ефект, містить два синаптичні контакти. Колатеральна гілка кожного волокна Іа прямує до гальмівного інтернейрона спинного мозку (пляшкоподібний нейрон Ґольджі), який утворює безпосередній синагітичний контакт з одним із мотонейронів, що іннервує м’яз-антагоніст. Приклад такого типу постсинаптичного гальмування наведено в Розділі 4, а його шлях зображений на рис. 4-9.
Зворотний рефлекс розтягування
Чим вище від точки сильного розтягування м’яза, тим сильніше виявляється рефлекторне скорочення. Проте коли ступінь напруження достатньо великий, то скорочення раптово припиняється і м’яз розслаблюється. Це розслаблення у відповідь на сильне розтягування називають зворотним інверсним рефлексом розтягування, або автогенним гальмуванням.
Рецептором зворотного рефлексу розтягування є сухожилковий орган Ґольджі (рис. 6-5) - сітчасте скупчення шишкоподібних нервових закінчень, розміщене серед пучків волокон сухожилка. Один сухожилковий орган відходить від 3 до 25 м’язових волокон. Нервові волокна, що відходять від сухожилкового органа Ґольджі, є мієліновими, швидкопровідними чутливими волокнами групи Іb (див. табл. 2-2). Подразнення цих волокон зумовлює ГПСП у мотонейронах, що іннервують м’язи, від яких відходять ці чутливі волокна. Волокна Іb закінчуються в спинному мозку на інтернейронах, а ті, відповідно, безпосередньо на мотонейронах (рис. 6-6). Ці волокна теж формують збуджувальні контакти з мотонейронами, які іннервують антагоністи м’яза.
Оскільки сухожилкові органи Ґольджі, на відміну від веретен, реєструють натяг серії м’язових волокон, то їхнє подразнення виникає внаслідок як пасивного розтягування, так і активного скорочення м’яза. Поріг збудження в сухо-жилкових органах Ґольджі невисокий. Рівень подразнення шляхом пасивного розтягування незначний, тому що більша частина м’язових еластичних волокон переймає на себе силу розтягування, і внаслідок цього сильне розтягування приводить не до скорочення, а до розслаблення. Проте скорочення м’яза зумовлює постійну імпульсацію, і сухожилковий орган Ґольджі, отже, функціонує як перетворювач у ланцюгу зворотного зв’язку, що регулює м’язову силу, подібно до того як ланцюг зворотного зв’язку веретена регулює довжину м’яза.
Про значення первинних закінчень веретен і сухожилкових органів Ґольджі в регулюванні швидкості м’язового скорочення, довжини м’яза і сили м’язового скорочення може свідчити той факт, що перерізання аферентних нервів, які прямують до ділянки плеча, спричинює вільне звисання верхньої кінцівки, як у разі геміпарезу. Структура описаної системи зображена на рис. 6-7, а взаємодія імпульсації веретена, імпульсації сухожилкового органа Ґольджі і реципрокної іннервації у визначенні рівня імпульсації в мотонейронах - на рис. 6-8.
М’язовий тонус
Резистентність м’яза стосовно розтягування часто характеризують як його тонус. Якщо руховий нерв м’яза перерізати, то м’яз виявлятиме дуже невелику резистентність і, як кажуть, стане розслабленим, кволим. Гіпертонічний (спастичний) м’яз - це м’яз, у якому резистентність щодо розтягування внаслідок гіперактивності рефлексу розтягування є високою. Десь між станом розслаблення й еластичності виникає нечіткий стан нормального тонусу. Гіпотонічність м’язів настає, насамперед, тоді, коли рівень у-еферентної імпульсації невисокий, а гіпертонічність - у протилежному випадку.
Реакція подовження
За умов гіпертонічності м’язів чітко простежується така послідовність: помірне розтягування → м’язове скорочення, сильне розтягування → м’язове розслаблення. У разі пасивного згинання в лікті, наприклад, фіксують негайну резистентність, що виникає внаслідок рефлексу розтягування триголового м’яза плеча. Подальше розтягування активує зворотний рефлекс розтягування. Резистентність щодо згинання раптово зникає і відбувається згинання в лікті. Продовження пасивного згинання знову розтягує м’яз, і послідовність повторюється. Така послідовність резистентності, коли кінцівка рухається пасивно, відома в клініці як ефект складаного ножа. Фізіологічний термін цього явища - реакція подовження, оскільки в цьому випадку відбувається реагування спастичного м’яза (у попередньому прикладі - триголового м’яза плеча) на подовження.
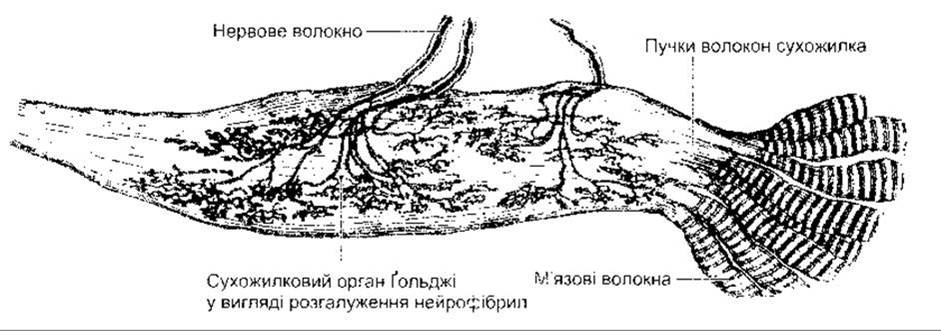
Рис. 6-5. Сухожилковий орган Ґольджі (відтворено за дозволом з Goss CM [editor]- Gray’s Anatomy of the Human Body, 29th ed. Lea & Febiger, 1973).
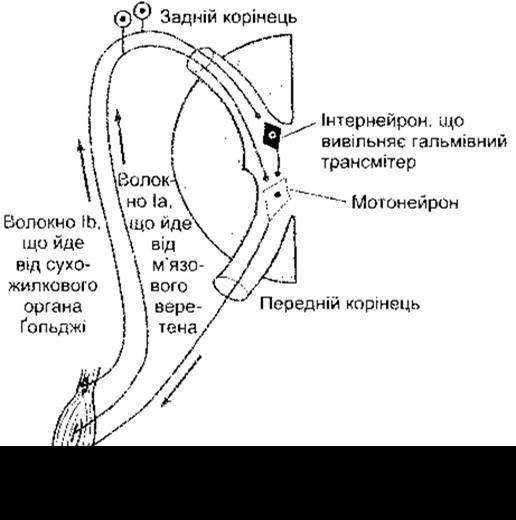
Рис. 6-6. Схема шляхів, що забезпечують рефлекс розтягування й інверсний рефлекс розтягування. Розтягування м’яза спричинює подразнення м’язового веретена, й імпульси, що поширюються волокнами Іа, спричинюють збудження в мотонейронах. Розтягування теж зумовлює подразнення сухожилкового органа Ґольджі, й імпульси, що поширюються волокнами Іb, спричинюють виділення інтернейроном гальмівного трансмітера - гліцину. В умовах сильного розтягування гіперполяризація, яка виникає в мотонейронах, є настільки значною, що імпульсація припиняється.
Клонус
Іншою ознакою станів, що супроводжуються зростанням у-еферентної імпульсації, є клонус. Цей неврологічний симптом полягає у регулярних ритмічних скороченнях м’яза, що піддається раптовому і тривалому розтягуванню. Типовим прикладом слугує гомілково-стопний клонус. Його ініціює різке і тривале тильне згинання стопи, а відповіддю є ритмічне згинання в гомілково-стопному суглобі. Основою реакції може бути описана вище послідовність: рефлекс розтягування - зворотний рефлекс розтягування. Проте реакція може відбуватися внаслідок синхронізованих збуджень у мотонейронах і без участі імпульсації з боку сухожилкового органа Ґольджі. М’язові веретена досліджуваного м’яза гіперактивні, і раптова поява імпульсів, ще поширюються від них, негайно зумовлює збудження в усіх мотонейронах, які іннервують м’яз. М’язове скорочення, що виникає внаслідок цього, припиняє імпульсацію веретен. Проте якщо продовжувати розтягування, то в розслабленому м’язі під час повторного розтягування знову виникає збудження веретен.