Фізіологія людини - Вільям Ф. Ґанонґ 2002
Функції нервової системи
Шкірна, глибока та вісцеральна чутливість
Провідні шляхи
За гістологічними характеристиками задні роги спинного мозку поділяють на пластинки I—VI; пластинка І є найближче до поверхні, а VI - найглибша. Пластинка II і частина пластинки IV утворюють драглисту речовину - слабко забарвлену ділянку біля верхівки кожного заднього рогу. Розрізняють три типи первинних чутливих волокон, які забезпечують шкірну чутливість: великі мієлінові Aß- волокна, які передають імпульси, що генеруються у відповідь на механічні подразнення; малі мієлінові Аß-волокна, з яких частина передає імпульси від холодових рецепторів і ноцирецепторів, що опосередковують швидкий біль (див. нижче), а інша частина - імпульси від механорецепторів; малі безмієлінові волокна С, які пов’язані головно з відчуттями болю і температури. Є також незначна кількість волокон С, які передають імпульси від механорецепторів. Розподіл цих волокон у задніх канатиках і різні шари заднього рогу показані на рис. 7-1.
Головні прямі провідні шляхи шкірної чутливості до кори головного мозку зображені на рис. 7-2. Волокна, які забезпечують відчуття дотику і пропріорецепцію, проходять через задні канатики до довгастого мозку, де закінчуються синапсами у ніжному та клиноподібному ядрах. Нейрони другого порядку від цих ядер перетинають середню лінію і піднімаються у складі медіальної петлі до заднього вентрального ядра і пов’язаних з ним специфічних сенсорних перемикальних ядер таламуса (див. Розділ 11). Цю висхідну систему часто називають дорсальним стовпом, або петельною системою.
Інші тактильні волокна разом із тими, що передають відчуття температури і болю, закінчуються синапсами на нейронах дорсального стовпа. Аксони цих нейронів перетинають середню лінію і проходять у передньолатеральному квадранті спинного мозку, де вони утворюють передньо-латеральну систему висхідних волокон. Інші волокна ідуть дорсальнільше. Загалом відчуття дотику пов’язане із вентральним спіноталамічним трактом, тоді як болю і температури - з латеральним, однак жорсткої локалізації функцій нема. Деякі волокна передньолатеральної системи закінчуються у специфічних перемикальних ядрах таламуса, інші проектовані на ядра середньої лінії, а також внутрішньопластинчасті неспецифічні проекційні ядра. Є значний потік імпульсації від передньолатеральної системи до ретикулярної формації середнього мозку. Отже, сенсорні імпульси активують ретикулярну активаційну систему, яка, відповідно, підтримує тонус кори мозку (див. Розділ 11). Колатералі від волокон, які є в складі задніх канатиків, проходять через задні роги. Вони можуть змінювати вхідну інформацію до інших шкірних сенсорних систем, у тім числі до больової системи. Задній ріг - це так звані ворота, у яких імпульси сенсорних нервових волокон перетворюються в імпульси висхідних трактів і, ймовірно, що проникність цих воріт залежить від природи і характеру імпульсів, які досягають губчастої речовини та прилеглих ділянок. Ці ворота також перебувають під впливом імпульсів, що йдуть по низхідних трактах від головного мозку. Зв’язок воріт з больовими імпульсами розглянуто нижче.
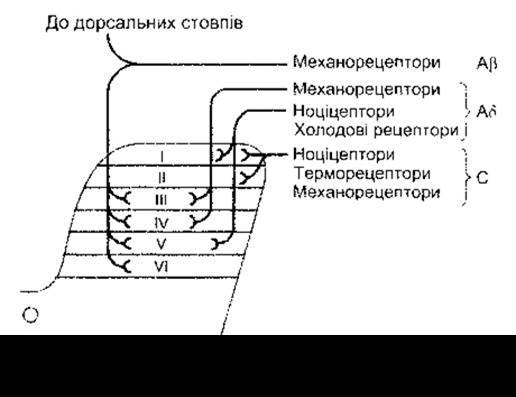
Рис. 7-1. Схематичне відтворення закінчень трьох типів первинних аферентних нейронів у різних шарах заднього рогу спинного мозку.
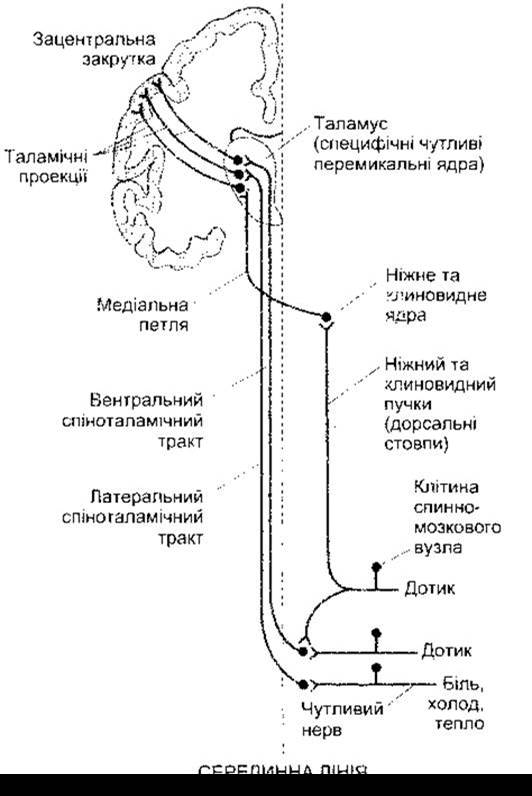
Рис. 7-2. Дотикові, больові та температурні шляхи від тулуба та кінцівок. Передньолатеральна система (вентральний і латеральний спіноталамічні, а також споріднені висхідні тракти) також проектується на ретикулярну формацію середнього мозку та неспецифічні таламічні ядра.
Аксони спіноталамічних трактів від крижових і поперекових сегментів зміщені латерально аксонами, які перетинають середню лінію на рівнях, що послідовно підвищуються. З іншого боку, волокна дорсального стовпа зміщені медіально волокнами вищих сегментів (рис. 7-3). З огляду на це обидві висхідні системи розділені на пластинки з відповідними шийними, грудними, поперековими і крижовими сегментами, що орієнтовані у медіально-латеральному напрямі в передньолатеральних шляхах, і від крижових до шийних сегментів, орієнтованих у медіально-латеральному напрямі у дорсальних стовпах. Завдяки такому поділу на пластинки пухлини, що виникають ззовні від спинного мозку, спочатку перетискають спіноталамічні волокна крижових і поперекових ділянок, спричинюючи ранній симптом втрати больової і температурної чутливості у крижовій ділянці. Пухлини, що виникають усередині спинного мозку, спочатку зумовлюють втрату чутливості у вищих сегментах.
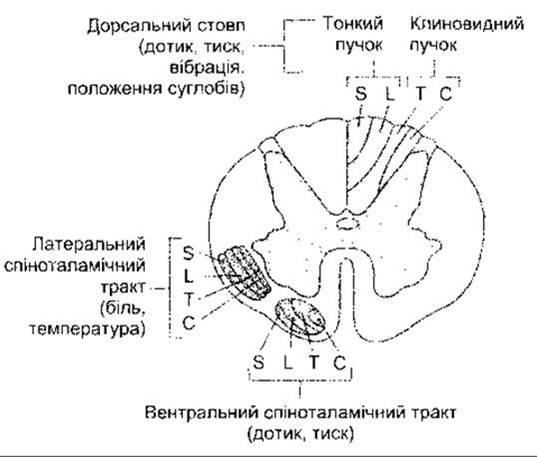
Рис. 7-3. Поперечний зріз спинного мозку. Показана локалізація висхідних чутливих шляхів. Зверніть увагу на пластинки у складі кожного з них: S - крижові; L - поперекові; Т - грудні; С - шийні.
У мозковому стовбурі до волокон петельної і передньо-латеральної систем долучаються волокна, що забезпечують чутливість голови. Перемикання больових та температурних імпульсів відбувається у спинномозковому ядрі трійчастого нерва, а тактильні і пропріорецептивні імпульси переважно перемикаються через головні чутливі та середньомозкові ядра цього нерва.
Кіркове представництво
Карти сенсорних зон кори складають за допомогою експериментів на тваринах і під час нейрохірургічних операцій у людей. Новітні способи картування кори головного мозку людини без хірургічних втручань ґрунтуються на використанні методів позитронної емісійної томографії (ПЕТ) та ядерно-магніторезонансних (ЯМР) зображень. Ці методи, описані у Розділі 32, стали основою більшості досягнень не тільки у сенсорній фізіології, а й у всіх аспектах досліджень нормальних кіркових функцій людини.
Від спеціальних сенсорних ядер таламуса нейрони, що переносять сенсорну інформацію, утворюють високоспецифічні проекції на дві соматосенсорні ділянки кори: соматосенсорну ділянку I (SI) у зацентральній закрутці і соматосенсорну ділянку II (SII) у Сильвієвій борозні. Окрім того, SI проектується на SII; SI відповідає 1-, 2- та 3-му полям Бродмана. Бродман був гістологом, який ретельно розподілив кору мозку на численні поля за особливостями їхньої гістологічної будови.
Таламічні волокна у ділянці SI організовані так, що частини тіла представлені впорядковано вздовж зацентральної закрутки - ноги на її вершині, а голова біля основи закрутки
(рис. 7-4). У зацентральній закрутці лише сфокусована точна топографія волокон від різних частин тіла, а й розміри ділянки кори, які отримують імпульси від певної частини тіла, пропорційні до кількості рецепторів у цій частині тіла. Відносні розміри кіркових ділянок показані на рис. 7-5, де пропорції гомункула змінені відповідно до розмірів цих ділянок кори. Сенсорні ділянки кори, які сприймають імпульси від тулуба і спини порівняно малі, тоді як ділянки, що сприймають імпульси від руки і частин рота, пов’язаних з мовою, - дуже великі.
Вивчення сенсорних ділянок засвідчує дискретну природу надзвичайно точної локалізації периферійних ділянок тіла у корі мозку і дає додаткові докази важливості теорії специфічності нервової енергії (див. Розділ 5). Стимуляція різних частин зацентральної закрутки зумовлює відчуття, які проектуються на відповідні частини тіла. Зазвичай таким способом можна створити відчуття оніміння поколювання або руху, однак за допомогою достатньо тонких електродів можна спричинити порівняно чіткі відчуття дотику, тепла, холоду або болю. Клітини у зацентральній закрутці організовані у вигляді вертикальних колонок, подібно до клітин зорової кори (див. Розділ 8). Усі клітини певної колонки активуються висхідними імпульсами від певної частини тіла й усі відповідають певній формі чуття.
SII локалізована у верхній частині Сильвієвої борозни, яка відокремлює скроневу частку від лобової та тім’яної; голова у нижньому кінці зацентральної закрутки, а ноги - на дно Сильвієвої борозни. Частини тіла тут представлені не так повно і детально, як у зацентральній закрутці.
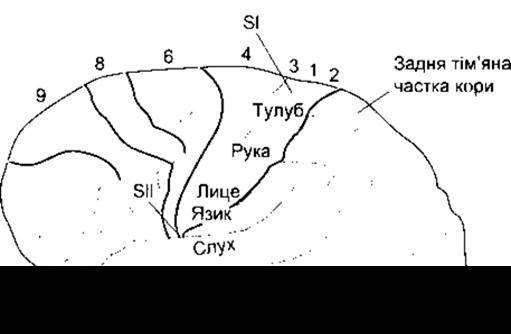
Рис. 7-4. Відповідність полів кори головного мозку соматичній чутливості, а також деякі ділянки кіркового сприйняття для інших типів чуття мозку людини. Цифрові позначки відповідають нумерації кіркових полів Бродмана. Первинна слухова ділянка локалізована у Сильвієвій борозні на вершині верхньої скроневої закрутки, і в нормі її не видно.
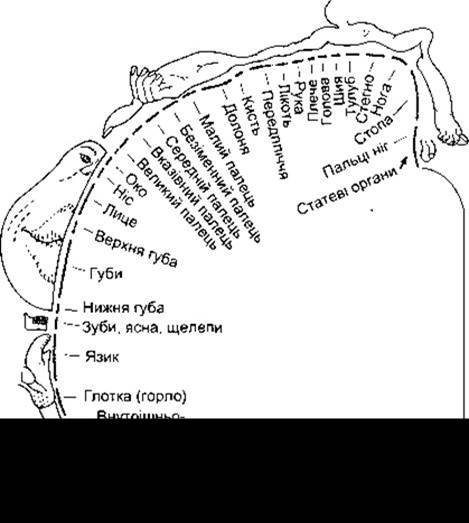
Рис. 7-5. Чутливий гомункул, що лежить на вінцевому розрізі через зацентральну закрутку (відтворено за дозволом з Рег- field W, Rasmussen G: The Cerebral Cortex of Man. Macmillan, 1950).
Кіркова пластичність
Сьогодні відомо, що численні нервові зв’язки, описані вище, не є вродженими і незмінними, і принаймні у сенсорних ділянках їх можна порівняно швидко змінити шляхом набуття досвіду, який передбачає використання відповідної ділянки. Наприклад, якщо у мавпи ампутувати палець, то кіркове представництво сусідніх пальців пошириться на ділянку, яка до цього відповідала ампутованому пальцю. І навпаки, якщо видалити ділянку кори, яка відповідає за той чи інший палець, то соматосенсорна карта цього пальця пересунеться до прилеглої кори. Широка довготривала деаферентація кінцівок призводить до ще більших зрушень соматосенсорного представництва у корі, коли, наприклад, кіркова ділянка кінцівки відповідає на дотик до обличчя. Крім того, якщо мавпа тренована розрізняти найдетальніші відмінності між різними вібраційними відчуттями на одному пальці однієї руки, то кіркове представництво цього пальця розширюється. Такі пластичні процеси відбуваються як упродовж розвитку, так і у дорослих. Наприклад, рецепторні поля окремих нейронів частини поля 3 сенсорної кори для пальців у нормі розташовані на одному пальці (рис. 7-6, ліворуч). Однак якщо дорослу мавпу постійно піддавати дії стимулів, які подразнюють лише дистальні частини першого, другого і третього пальців, то рецепторні поля окремих нейронів поширяться на дистальні частини усіх трьох пальців (див. рис. 7-6). Пояснення цих зрушень, очевидно, полягає у тому, що кіркові зв’язки сенсорних одиниць кори мають велику здатність до конвергенції і дивергенції зі зв’язками, які можуть ставати слабкими від невикористання і сильними від використання. У щурів цей процес пов’язаний з базальною частиною кінцевого мозку: імпульси з цієї ділянки кори збільшуються у разі використання слухових подразників. Розміри слухової сенсорної ділянки стають більшими, ніж тоді, коли подразнення поєднують з низьким рівнем імпульсів із базальної частини головного мозку.
Пластичність цього типу простежується в разі надходження вхідних імпульсів не лише від шкірних рецепторів, а також від інших сенсорних систем. Наприклад, у котів із невеликими ушкодженнями сітківки кіркова ділянка сліпої плями починала відповідати на світлове подразнення інших ділянок сітківки. Розвиток проекцій сітківки дорослого на зорову кору є іншим прикладом цієї пластичності. В екстремальних випадках експериментальне скерування зорової інформації у слухову кору під час розвитку створювало зорові рецепторні поля у слуховій системі.
У людей також відбуваються пластичні зміни, подібні до тих, що описані вище в експериментальних тварин. Наприклад, у деяких інвалідів із ампутованою рукою дотик до обличчя зумовлює відчуття, що проектуються на руку, якої нема. ПЕТ дала змогу також виявити пластичні зміни однієї форми чутливості на іншу. Наприклад, тактильні та слухові стимули збільшують метаболічну активність зорової кори у сліпих осіб. Навпаки, глухі швидше і точніше реагують на рухові стимули в ділянці периферійного зору порівняно з нормальними людьми. Пластичність характерна також для рухової кори (див. Розділ 12). Наведені приклади свідчать про гнучкість мозку і його здатність до адаптації.
Ефекти ушкоджень мозкової кори
Видалення соматичної сенсорної ділянки І у тварин зумовлює недостатність у відчутті пози та змозі розрізняти розмір і форму, а соматичної сенсорної ділянки II - порушує здатність до навчання розрізняти дотики. Видалення SI призводить до дефіциту опрацювання сенсорної інформації у SII, тоді як видалення SII незначно впливає на процес опрацювання інформації у SI. Очевидно, ділянки SI і SII опрацьовують сенсорну інформацію швидше послідовно, аніж паралельно. Ділянка SI також проектується на задню тім’яну кору (див. рис. 7-4), і ушкодження цієї асоціативної ділянки спричинює комплекс відхилень у просторовій орієнтації протилежної частини тіла (див. Розділ 16)
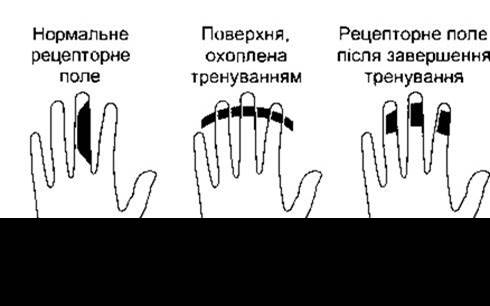
Рис. 7-6. Роль вхідних імпульсів в утворенні рецепторних полів кіркових нейронів. Ліворуч: нормальне рецепторне поле одного нейрона кори головного мозку на бічній поверхні середнього пальця мавпи. Посередині: тренування мавпи до виконання завдань, що потребують подразнення дистальних частин першого, другого та третього пальців. Праворуч: змінене внаслідок тренувань рецепторне поле кіркового нейрона (за Wang et аl, модифіковано з Sur М: Maps of time and space. Nature 1995;378:13).
Зазначимо, що і в експериментальних тварин, і у людей пошкодження кори не означає, що зникнуть тілесні відчуття. Найлегше уражаються внаслідок кіркових ушкоджень пропріорецепція і відчуття дотику. Менш вразливими є відчуття температури і лише злегка уражається больове відчуття. Отже, сприйняття можливе й тоді, якщо кори нема. Під час одужання, відчуття болю повертається першим, за ним відновлюється температурне відчуття, і, нарешті, пропріорецепція і легкий дотик.
Принципи сенсорної фізіології
Найважливіші загальні принципи, що стосуються фізіології сенсорних систем, детально описані у Розділі 5. Кожний орган чуття спеціалізований на перетворенні окремого виду енергії у потенціал дії сенсорних нервів. Кожна форма чуття має окремий шлях до мозку, а отримане відчуття, так само, як і частина тіла, у якій воно локалізоване, визначене окремою ділянкою активованого мозку.
Відмінності в інтенсивності того чи іншого відчуття передаються двома способами: зміною частоти потенціалів дії сенсорних нервів і зміною кількості активованих рецепторів. Збільшення інтенсивності подразнення органа чуття на якість відчуття впливає дуже слабко або не впливає зовсім.
Іншим принципом, який стосується органів чуття шкірної локалізації, є принцип точкового представництва. Якщо виконати детальне, міліметр за міліметром, картування шкіри за допомогою тонкого волоска, то можна з’ясувати, що відчуття дотику відходить з точок, які лежать над тактильними рецепторами. Його не можна викликати з ділянок, що лежать між цими точками. Подібно цього, відчуття болю і температури спричинені подразненням шкіри лише над тими точками, де розташовані відповідні рецепторні структури для цього подразника.