Фізіологія людини - Вільям Ф. Ґанонґ 2002
Функції нервової системи
Зір
Кольоровий зір
Характеристики кольору
Колір має три головні характеристики: відтінок, інтенсивніть та насиченість (її визначають як ступінь відсутності білого кольору). Для кожного кольору є комплементарний колір, змішування з яким у відповідній пропорції дає білий колір. Сприйняття чорного кольору зумовлене відсутністю світла, однак воно, ймовірно, несе позитивну інформацію, оскільки сліпе око не бачить чорного: воно не бачить нічого. Відомі явища послідовних і паралельних контрастів, оптичних ілюзій, що дають відчуття кольору, коли його нема, негативні та позитивні залишкові образи, а також різноманітні психологічні аспекти кольорового зору. Детальна характеристика цих явищ не є метою цієї книги.
Іншим чинником, що має важливе значення, є змога отримання білого або будь-якого іншого кольору спектра і навіть позаспектрального пурпурного кольору шляхом змішування у різних пропорціях червоного (довжина хвилі 723-647 нм), зеленого (575-492 нм) та синього (492- 450 нм) світла. Саме тому червоний, зелений і синій називають первинними кольорами. Ще одним важливим чинником є спостереження, зроблене Ландом, відповідно до якого сприйняття кольору частково залежить також від кольору інших об’єктів у полі зору. Наприклад, червоний предмет сприймається як червоний, якщо його освітлено зеленим або синім світлом, проте він набуває блідо-рожевого або навіть білого кольору у випадку освітлення червоним світлом.
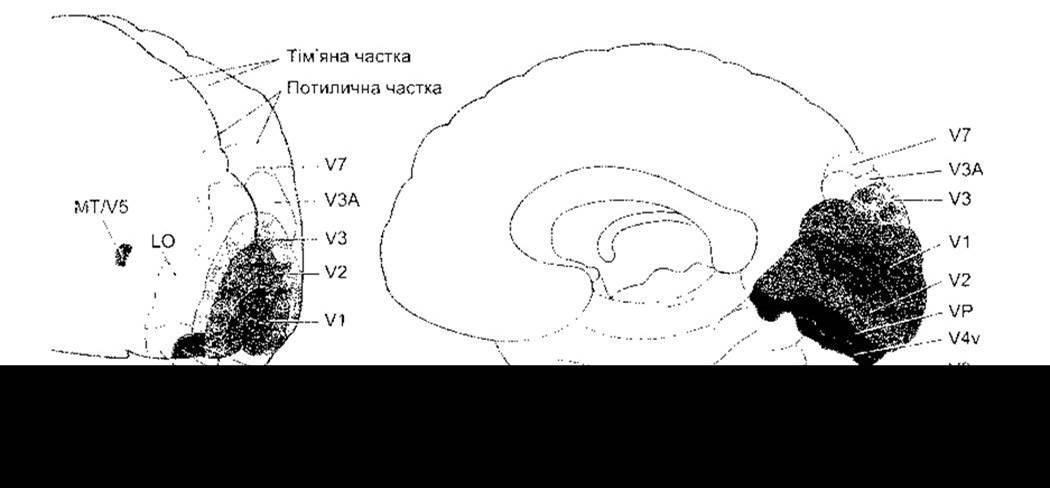
Рис. 8-25. Частина головних зон первинної зорової кори (V1) мозку людини. Вигляд збоку і зсередини. Див. також табл. 8-1 (модифіковано з Logothetis N: Vision: a window on consciousness. Sсi Am [Nov] 1999;28:99).
Сітківкові механізми
Теорія кольорового зору Юнґа-Гельмгольца передбачає наявність у людини трьох типів колбочок, кожен з яких містить особливий світлочутливий пігмент з максимальною чутливістю до одного із трьох первинних кольорів, а відчуття будь-якого кольору визначене відносною частотою імпульсів від кожного типу колбочок. Правильність цієї теорії доведена за допомогою ідентифікації та хімічної характеристики кожного з трьох пігментів. Один із них (синьочутливий, або короткохвильовий пігмент) має максимум поглинання у синьо-фіолетовій частині спектра (рис. 8-26); інший (зеленочутливий, або середньохвильовий) - у зеленій, а ще інший (червоно-чутливий, або довгохвильовий) - у жовтій частині спектра. Колбочки з максимальною чутливістю у жовтій частині спектра достатньо чутливі також у його червоній частині, маючи для червоного світла нижчий поріг збудження, ніж для зеленого. Усі ці експериментальні дані задовольняють вимоги теорії Юнґа-Гельмгольца.
Таблиця 8-1. Функція ділянок зорових проекцій мозку людини1
V1 |
Первинна зорова кора; отримує імпупьси з бічних копінчастих ядер, розпочинає опрацювання інформації стосовно орієнтації, країв об’єктів тощо |
V2, V3, VP |
Продовжують опрацювання зображень, мають більші зорові поля |
V3A |
Рух |
V4v |
Функція невідома |
MT/V5 |
Контролювання руху |
LO |
Розпізнавання великих об’єктів |
V7 |
Функція невідома |
V8 |
Кольоровий зір |
1 Модифіковано з Logothetis N: Vision: a window on consciousness. Sсi Am (Nov) 1999,281:99.
У людини ген родопсину локалізований на 3-й хромосомі, а ген, що кодує синтез синьочутливого S-пігменту колбочок, - на 7-й. Два інші пігменти колбочок кодовані двома спареними генами, що розміщені на q-плечі X- хромосоми. Зеленочутливий пігмент М та червоночутливий пігмент L подібні за будовою: їхні опсини виявляють 96% гомології амінокислотного складу, тоді як кожен з цих пігментів має лише 43% гомології з опсином синьочутливого пігменту, й усі три колбочкові пігменти виявляють близько 41% гомології з родопсином. Багато ссавців є дихроматами, тобто мають лише два типи колбочкових пігментів - коротко- та довгохвильовий. Мавпи Старого Світу і люди є трихроматами з окремими середньо- та довгохвильовими пігментами, що, ймовірно, було зумовлене дублікацією довгохвильового гена предків з наступною дивергенцією.
Недавно виявлено, що в межах людської популяції є певні варіанти довгохвильових (червоночутливих) пігментів. Віддавна відомо, що відповідь на пропорцію Рейлі, тобто співвідношення кількості червоного і зеленого світла, яке різні індивіди змішують для отримання монохроматичного оранжевого, є бімодальною.
Цей феномен добре пояснюють нові дані про те, що 62% осіб з нормальним кольоросприйняттям у 180-му положенні ланцюга довгохвильового опсину колбочок мають залишок серину, тоді як решта 38% - аланіновий залишок.
Пік поглинання пігменту в осіб з сериновим залишком у 180-му положенні відповідає довжині хвилі 556,7 нм, тобто цей варіант пігменту є чутливішим до червоного світла, тоді як крива осіб з аланіновим залишком має пік поглинання 552,4 нм.
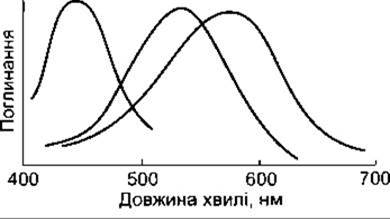
Рис. 8-26. Спектри поглинання трьох типів колбочкових піґментів сітківки людини. Піґмент S з піком поглинання 440 нм забезпечує сприйняття синього, а піґмент М з піком поглинання 535 нм - зеленого кольорів. Піґмент L з піком поглинання 565 нм (жовтий колір спектра) сприймає також довгі хвилі червоної частини спектра (відтворено за дозволом з Michael CR: Color vision. N Engl J Med 1973;288:724).
Нервові механізми
Інформацію про колір передають гангліонарні клітини, які віднімають або додають імпульси, що надходять від колбочок різних типів. Опрацювання інформації у гангліонарних клітинах та нейронах бічного колінчастого тіла забезпечує формування імпульсів, що трьома різними шляхами надходять до V1: червоно-зелений шлях передає сигнали, які відображають різницю між відповідями L- та М-колбочок; синьо-жовтий шлях - сигнали, що відображають різницю між відповіддю S-колбочок та сумарною відповіддю L- і М-колбочок; третій шлях - сумарну відповідь L- та М-колбочок. Ці три шляхи проектуються на зону плямок та глибоку частину шару 4С поля V1. Від зони плямок та шару 4 інформація стосовно кольору передається до поля V8. Механізм подальшої трансформації сигналів, що надходять до V8, у відчуття кольору, до цього часу не з’ясований.
Кольорова сліпота
Є численні тести для виявлення кольорової сліпоти. Найпоширеніші - це підбір ниток відповідного кольору і таблиці Ішігара (в Україні використовують поліхроматичні таблиці Рабкіна). У першому варіанті обстежуваному дають набір ниток різного кольору і пропонують виокремити нитки подібних кольорів. Таблиці Ішігара та подібні до них містять цифри, що складаються з різноколірних плямок, фоном для яких слугують подібні за формою плямки іншого кольору. Цифри навмисне намальовані кольорами, що зливатимуться з фоном для особи, що має кольорову сліпоту.
Частина осіб з кольоровою сліпотою не здатні розрізняти певні кольори (анопія), тоді як інші мають лише ослаблене сприйняття кольорів (аномалія). Префікси прот-, дейтер- та трит - стосуються дефектів червоної, зеленої та синьої колбочкових систем, відповідно. Осіб з нормальним кольоросприйняттям, а також осіб з протаномалією, дейтераномалією та тританомалією називають трихроматами: у них наявні усі три колбочкові системи, одна з яких може бути ослабленою. Дихроматами є особи, що мають лише дві колбочкові системи: у них може бути протанопія, дейтеранопія або тританопія. Монохромати мають тільки одну колбочкову систему. Дихромати можуть урізноманітнювати спектр свого кольоросприйняття шляхом комбінування двох первинних кольорів, монохромати можуть лише змінювати інтенсивність одного кольору.
Кольорова сліпота найчастіше є спадковим захворюванням. Іноді вона простежується в осіб з ушкодженнями поля V8 (див. вище), у яких виникає ахроматопсія. Окрім того, в чоловіків, які уживають віагру для лікування еректильної дисфункції, виникає тимчасова слабкість сприйняття синьо-зеленого кольору, оскільки цей лікарський засіб пригнічує активність фосфодіестерази сітківки (див. Розділ 23).
Успадкування кольорової сліпоти
Популяційні дослідження осіб, що походять з кавказького регіону дали змогу виявити, що аномалії кольоросприйняття успадковують 8% чоловіків та 0,4% жінок. Тританомалія та тританопія порівняно мало поширені і не виявляють статевої вибірковості. Близько 2% чоловіків з кольоровою сліпотою є дихроматами з протанопією або дейтеранопією, близько 6% є аномальними трихроматами з порушеною спектральною чутливістю червоно- або зеленочутливого пігментів. Ці аномалії успадковані за рецесивним типом і пов’язані з Х-хромосомою, тобто з’являються за наявності аномального гена у складі Х-хромосоми. Оскільки всі соматичні клітини чоловічого організму додатково до 44 аутосом містять одну X- та одну У-хромосому (див. Розділ 23), то кольорова сліпота виникає у чоловіків за наявності дефектного гена в складі їхньої X- хромосоми. Соматичні клітини жіночого організму містять дві Х-хромосоми - по одній від кожного з батьків. З огляду на рецесивний характер успадкування цієї патології кольорова сліпота виявляється у жінок тільки за наявності дефектних генів в обох Х-хромосомах. Половина дівчаток, народжених від чоловіків з Х-зумовленою кольоровою сліпотою, є носіями прихованої кольорової сліпоти і передають цей дефект половині своїх синів. Тому пов’язана з Х-хромосомою кольорова сліпота перескакує через покоління і виявляється у чоловіків кожного другого покоління. Гемофілія, міодистрофія Дюшенна та багато інших спадкових захворювань зумовлені мутаціями генів Х-хромосоми.
Порівняно значне поширення дейтер- та протаномалії, ймовірно, пов’язане з молекулярною організацією генів, що кодують синтез зелено- та червоночутливих пігментів колбочок. Вони розміщені один біля одного (спарені «головою до хвоста» у складі q-плеча Х-хромосоми) і схильні до нерівнозначної гомологічної рекомбінації (нерівнозначний кросинговер) у процесі розвитку статевих клітин. Це призводить до виникнення гібридних пігментів зі зміненою просторовою чутливістю; значна кількість таких гібридів уже досліджена й описана.