Фізіологія людини - Вільям Ф. Ґанонґ 2002
Функції нервової системи
Слух та рівновага
Слух
Звукові хвилі
Звук є відчуттям, яке виникає під час потрапляння на барабанну перетинку поздовжніх коливань, а точніше - фаз згущення і розрідження молекул зовнішнього середовища. Результатом цих рухів є зміна тиску на барабанну перетинку за одиницю часу, що має вигляд серії хвиль (рис. 9-11). Сукупність таких рухів зовнішнього середовища називають звуковими хвилями. Звукові хвилі поширюються у повітрі зі швидкістю 344 м/с при 20°С на рівні моря. Швидкість хвиль зростає з підвищенням температури та висоти над рівнем моря. Інші середовища, у які може потрапляти людина, також проводять звукові хвилі, проте з іншою швидкістю. Наприклад, швидкість звуку у прісній воді при 20°С становить 1450 м/с, у морській воді вона ще вища. Вважають, що свист голубого кита досягає сили 188 децибел (див. нижче), і його чути на відстані 900 км.
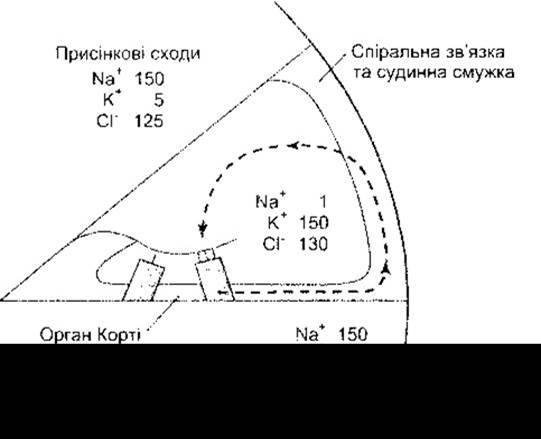
Рис. 9-9. Йонний склад перилімфи присінкових та барабанних сходів, а також ендолімфи середніх сходів. Штриховою лінією показаний шлях рециклізації К+: від волоскових клітин через підтримувальні клітини до спіральної зв’язки і через клітини судинної смужки до ендолімфи.
Сила звуку корелює з амплітудою звукової хвилі, а висота тону - з її частотою (кількістю коливань за одиницю часу). Чим більша амплітуда, тим голосніший звук, чим вища частота коливань - тим вищий тон. Окрім частоти, на висоту тону впливають і деякі інші, не зовсім зрозумілі фактори; частота дещо впливає і на силу звуку, оскільки поріг звукового подразнення для певних частот нижчий, ніж для інших (див. нижче). Звукові хвилі, що повторюються, навіть у разі їхньої комплексності, сприймаються як музичні звуки; неперіодичні, неповторювані коливання дають відчуття шуму. Більшість музичних звуків складається з хвилі головної частоти, що визначає висоту звуку, на яку накладається низка гармонійних коливань (обертонів), що надають звукові характерного тембру (якість звуку). Відмінності тембру дають змогу розрізняти звучання різних музичних інструментів, навіть якщо виконують звуки однакової висоти.
Амплітуду звукової хвилі можна виразити як максимальну зміну тиску на барабанну перетинку, однак зручнішою є відносна, так звана децибельна шкала. Інтенсивність звуку в белах є записом логарифмічної залежності значення інтенсивності досліджуваного звуку до певного звукового стандарту: 1 децибел (дБ) становить 0,1 бела. Отже,
![]()
Інтенсивність звуку пропорційна до площі тиску звуку. Тому
![]()
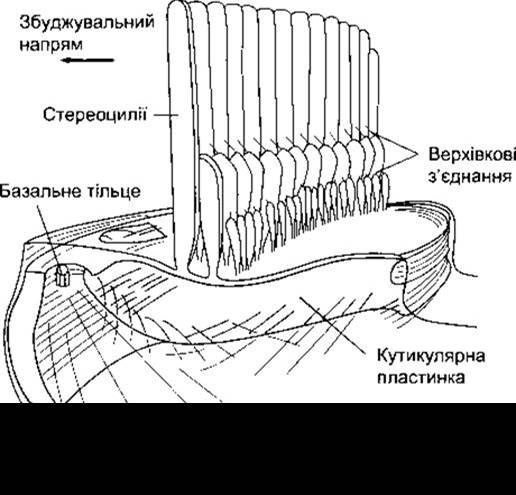
Рис. 9-10. Будова апікальної частини волоскової клітини. Зверніть увагу на верхівкові з’єднання між рядами стереоцилій. Стрілкою показаний напрям, за яким зміщення стереоцилій призводить до збільшення потоку йонів усередину волосків (відтворено за дозволом з Hackney CM, Furness DN: Mechanotransduction in vertebrate hair cells: Structure and function of the stereociliary bundle. Am J Physiol 1995;268:C1).
Відповідно до стандарту Американського Акустичного Товариства за 0 децибел прийнятий тиск 0,000204 Н/см2, що є порогом слухових відчуттів середньостатистичної людини. На рис. 9-12 показані значення у децибелах найпоширеніших звуків. Треба пам’ятати, що шкала децибел є логарифмічною. Тому 0 децибел не означає, що звуку нема, а відображає наявність звуку, інтенсивність якого відповідає стандартові. Крім того, діапазон між 0 та 140 децибелами, який охоплює рівні від нижнього порогу слухового відчуття до такого, що може пошкодити орган Корті, практично відображає десятимільйонну (107) різницю в тискові, який чинять хвилі на барабанну перетинку. Іншими словами, якщо атмосферний тиск на рівні моря становить 1 бар, то діапазон інтенсивності від звуку ледь чутного до потенційно руйнівного для завитки, охоплює тиск від 0,0002 до 2000 мікробарів.
Людина може чути звукові частоти від 20 до 20 000 Гц (герц, або коливань за секунду). Деякі тварини, особливо кажани та собаки, чують звуки набагато вищої частоти. Пороги відчуттів людського вуха відрізняються для тонів різної висоти, досягаючи максимальної чутливості в діапазоні 1-4 кГц. Для порівняння, тон чоловічого голосу становить близько 120 Гц, тон голосу середньо статистичної жінки - 250 Гц. Пересічна людина може розрізнити до 2 000 тонів, однак музичні вправи дають змогу цю кількість значно збільшити. Найлегше розрізняють тони у діапазоні 1-3 кГц, значно важче - тони вищих або нижчих частот (рис. 9-13).
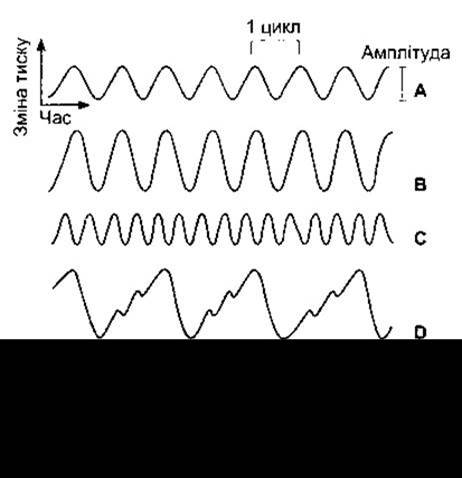
Рис. 9-11. Характеристика звукових хвиль: А - запис простого тону; В - запис звуку з більшою амплітудою і голоснішого, ніж на фрагменті А; С - звук з такою ж амплітудою, що й на фрагменті А, проте більшої частоти і, відповідно, вищого тону; D - складна звукова хвиля, що періодично повторюється. Фрагменти А-D є графічним відтворенням музичних звуків, тоді як хвилі на фрагменті Е не упорядковані і мають характер шуму.
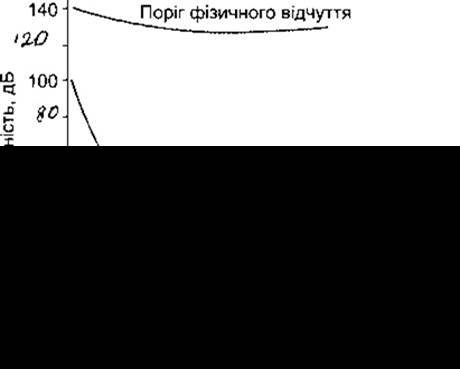
Рис. 9-13. Графіки звукосприйняття людини. Середня крива отримана внаслідок аудіометрії у звичайних умовах, нижня - в ідеальних умовах. У разі 140 дБ звуки чують, а також відчувають фізично.
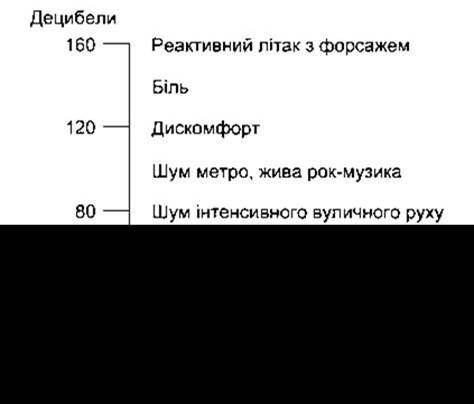
Рис. 9-12. Інтенсивність найпоширеніших звуків.
Маскування
Загальновідомо, що один звук може погіршувати здатність особи чути інші звуки. Це явище називають маскуванням. Причиною маскування вважають відносну або абсолютну рефрактерність попередньо стимульованих слухових рецепторів та нервових волокон до нових стимулів. Ступінь маскування залежить від тону. Маскувальний ефект шуму оточення, якщо нема надійного протишумового захисту, призводить до певного підвищення слухового порогу.
Передавання звуків
У вусі звукові хвилі, що надходять із зовнішнього середовища, перетворюються на потенціали дії слухових нервів. За участю барабанної перетинки та слухових кісточок звукові коливання передаються на основу стремінця, рухи якого спричинюють коливання рідини у внутрішньому вусі. Під дією цих коливань орган Корті генерує потенціали дії у нервових волокнах.
Функції барабанної перетинки та слухових кісточок
У відповідь на зміни тиску, спричинені дією звукових хвиль на її зовнішню поверхню, барабанна перетинка рухається всередину і назовні. Вона діє як резонатор, що відтворює коливання джерела звуку. Коливання барабанної перетинки припиняються майже одночасно із припиненням звуку, тобто вона має дуже короткий період критичного загасання. Рухи барабанної перетинки передаються на рукоятку молоточка, який обертається навколо осі, що проходить через точку сполучення його довгого і короткого відростків, унаслідок чого короткий відросток передає коливання на коваделко, а воно, відповідно, - далі на головку стремінця. Рухи головки стремінця змушують його основу рухатися вперед-назад подібно до шарнірних дверей, закріплених біля заднього краю овального вікна. Таким способом слухові кісточки діють як система важелів, що перетворює резонансні коливання барабанної перетинки у рухи стремінця, а через нього - у рухи перилімфиприсінкових сходів завитки (див. рис. 9-2). Ця система підсилює тиск звукових хвиль, що доходять до овального вікна, оскільки важільна система молоточка і коваделка збільшує силу поштовхів у 1,3 раза, а площа барабанної перетинки значно перевищує площу основи стремінця. Хоча інерційність та опір усієї системи і зумовлюють деяку втрату енергії звуків, та підраховано, що за частоти до 3 000 Гц близько 60% звукової енергії, що потрапляє на барабанну перетинку, досягає перилімфи присінкових сходів.
Барабанний рефлекс
Під час скорочення м’язи середнього вуха (м’яз стремінця та м’яз-натягач барабанної перетинки) тягнуть молоточок досередини, а основу стремінця - назовні. Це послаблює передавання звуку. Голосні звуки спричинюють рефлекторне скорочення означених м’язів, що називають барабанним рефлексом. Барабанний рефлекс має захисне значення, запобігаючи надмірному стимулюванню слухових рецепторів сильними звуками. Реакційний час барабанного рефлексу становить 40-160 мс, тому він не може захистити від коротких інтенсивних звуків, наприклад, звуків пострілів.
Кісткова і повітряна провідність
Поширення звукових хвиль через барабанну перетинку та слухові кісточки до рідини внутрішнього вуха - головний шлях нормального слуху - називають кісточковою провідністю. Звукові хвилі також спричинюють коливання вторинної барабанної перетинки, якою закрите кругле вікно. Це явище називають повітряною провідністю, воно не відіграє значної ролі у нормальному звукосприйнятті. Третій тип звукопоширення - кісткова провідність - полягає у передаванні коливань із зовнішнього середовища до рідини внутрішнього вуха кістками черепа. Виражена кісткова провідність виникає в разі прикладання камертона або іншого вібрувального предмета безпосередньо до черепа. Цей тип провідності також має значення в разі сприйняття дуже голосних звуків.
Пересувні хвилі
Рухи основи стремінця зумовлюють виникнення серії пересувних хвиль у перилімфі присінкових сходів. Графічно такі хвилі зображено на рис. 9-14. З пересуванням хвилі по завитці її висота досягає максимуму у певній точці, після чого швидко спадає. Відстань від основи стремінця до місця у завитці, де хвиля досягає максимальної висоти, залежить від частоти коливань, що зумовлюють її виникнення. Високочастотні звуки генерують хвилі, що досягають макимуму біля основи завитки; наслідком дії низькочастотних звуків є хвилі з максимумом біля верхівки завитки. Кісткові стінки вестибулярних сходів жорсткі, проте гнучкою є мембрана Рейсснера. Базилярна мембрана під впливом тику також може легко зміщуватися в барабанні сходи піками хвиль, що виникають у присінкових сходах. Коливання перилімфи барабанних сходів розсіюються у повітрі через кругле вікно. Таким способом звукові хвилі зумовлюють зміщення базилярної мембрани, у цьому разі точка максимального зміщення визначена частотою хвилі. Апікальну частину волоскових клітин органа Корті фіксує ретикулярна пластинка, волоски зовнішніх волоскових клітин включені у покривну мембрану (див. рис. 9- 4). Під час рухів стремінця обидві означені мембрани зміщуються в тому самому напрямі, та оскільки вони орієнтовані у просторі під різними кутами, то їхнє взаємозміщення призводить до згинання волосків. Волоски внутрішніх волоскових клітин не торкаються покривної мембрани, однак вони також згинаються під дією зміщень ендолімфи у проміжку між покривною мембраною та верхівками волоскових клітин.
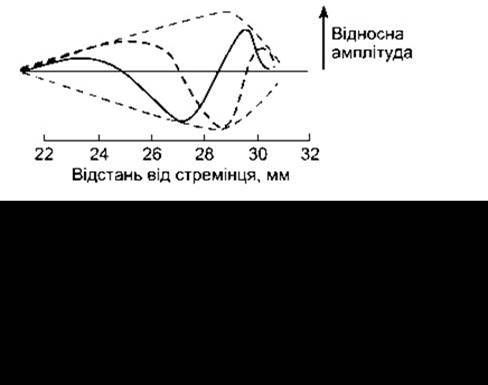
Рис. 9-14. Пересувні хвилі. Угорі: суцільна і короткоштрихова лінії відображають хвилі, зафіксовані в різні часові моменти. Довгоштрихова лінія відображає звуковий «коридор», що утворюється у разі сполучення лінією піків, які виникають у часовій прогресії. Внизу: зміщення базилярної мембрани хвилями, що утворюються внаслідок коливань стремінця з частотою, зазначеною біля піка кожної кривої.
Функції внутрішніх та зовнішніх волоскових клітин
Внутрішні волоскові клітини є головними сенсорними клітинами органа Корті, що зумовлюють генерування потенціалів дії у слухових нервах, правдоподібно, у відповідь на описані вище зміщення рідини.
Зовнішні волоскові клітини іннервовані холінергічними еферентними нервовими волокнами, що надходять з верхньооливних комплексів. Ці клітини стають нижчими в разі деполяризації і видовжуються у випадку гіперполяризації. Зовнішні волоскові клітини гіперполяризують під дією ацетилхоліну, що його виділяють еферентні нервові волокна. Функція цих клітин полягає у збільшенні амплітуди і загостренні піків вібрації базилярної мембрани, хоча тонкі механізми цього процесу вивчені недостатньо.
Потенціали дії слухових нервових волокон
Частота потенціалів дії окремих слухових нервових волокон пропорційна до сили звукових подразнень. За низької інтенсивності звуку кожен аксон розряджається лише під дією звуку певної частоти, відповідно до частини завитки, яку він репрезентує. У разі вищої інтенсивності звуку аксони розряджаються у відповідь на ширший спектр звукових частот (рис. 9-15), а точніше, реагують на хвилі, частота яких нижча від порога збудження.
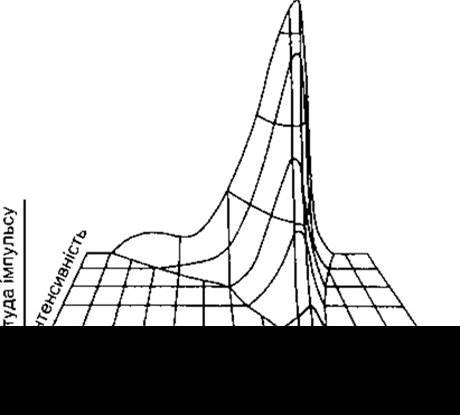
Рис. 9-15. Залежність амплітуди імпульсів у волокнах слухового нерва від частоти та інтенсивності звуку. Оскільки діаграма відображає загальний тип відповіді багатьох нервових волокон, то шкала не містить цифрових показників (модифіковано і відтворено за дозволом з Kiang NYS: Peripheral neural processing of auditory information. Handbook of physiology. Section 1, The Nervous System, vol 3, part 2. Brookhart JM, Mountcastle VB [editors] American Physiological Society, 1984).
Головним чинником, який визначає висоту сприйнятого звуку, є ділянка максимального стимулювання органа Корті. Пересувна хвиля, спричинена звуком певної частоти, зумовлює максимальне вгинання базилярної мембрани і, відповідно, максимальне збудження рецепторних клітин лише в одній точці. Як зазначено вище, відстань між цією точкою й основою стремінця обернено пропорційно залежить від висоти звуку: низькі тони зумовлюють максимальне стимулювання біля верхівки завитки, високі - біля її основи.
Провідні шляхи, що йдуть від різних частин завитки до мозку, відрізняються. Додатковим чинником сприйняття тонів з частотою до 2 000 Гц може бути структура потенціалів дії слухового нерва. У разі низької частоти звуку нервові волокна відповідають імпульсом на кожний цикл коливань звукової хвилі. Значення цього ефекту лавини імпульсів обмежене, оскільки частота потенціалів дії певного нервового волокна визначає більше силу, аніж висоту звуку.
Хоча висота тону головно залежить від частоти звукової хвилі, та деяку роль у його сприйнятті відіграє також сила звуку; низькі тони (до 500 Гц) здаються нижчими, а високі (понад 4 000 Гц) - вищими у разі збільшення сили звуку. Тривалість звуку також незначно впливає на сприйняття його тону. Тональність звуку неможливо визначити, якщо він триває до 0,01 с; у проміжку між 0,01 та 0,1 с сприйняття висоти тону зростає зі збільшенням тривалості звучання. Нарешті, висота комплексних звуків, що складаються з сукупності гармонійних частот, сприймається навіть без головної частоти (базового тону).
Слухові відповіді нейронів довгастого мозку
Відповідь окремих нейронів другого порядку ядра завитки на звукові подразнення подібна до відповідей окремих волокон слухового нерва. Частота, за якої звуки найнижчої інтенсивності зумовлюють відповідь, відрізняється між окремими сегментами; зі збільшенням сили звуку діапазон частот, на які генерується відповідь, зростає. Головна відмінність між відповідями нейронів першого та другого порядків полягає у гострішому «запиранні» мозкових нейронів з низькочастотного боку. Можливо, вища специфічність нейронів другого порядку зумовлена певними гальмівними процесами у мозковому стовбурі, однак механізм цього явища поки що не з’ясований.
Первинна слухова кора
Провідні шляхи від завитки до слухових ділянок кори мозку схарактеризовані на початку цього розділу. Висхідні імпульси проходять з дорсального та вентрального завиткових ядер по складних шляхах, частина з яких перетинається, інші - не перетинаються. У тварин є впорядкована система представництва різних тонів у первинній слуховій корі - ніби над нею розкручена завитка. У людини низькі тони сприймають передньолатеральні, а високі - задньо-медіальні ділянки слухової кори. В цьому випадку у мозковій корі кодується висота тону, а не власне звукова частота, бо під час звучання комплексного звуку без головного тону (див. вище) стимулюється та ділянка кори мозку, яка відповідає сприйнятій тональності. Тому перетворення частоти звуку на тональність повинні відбуватися на субкортикальному рівні.
Інші ділянки кори, пов’язані зі слухом
Використання зображень позитронної емісійної томографії та функційного ЯМР (див. Розділ 16) забезпечило швидке збільшення обсягу інформації стосовно асоціативних слухових зон мозку людини. Слухові шляхи мозкової кори нагадують зорові шляхи складністю процесів опрацювання інформації, а також наявністю двох головних шляхів. У випадку слухового сприйняття дорсально-тім’яний шлях, правдоподібно, пов’язаний з локалізацією джерела звуку (дає відповідь на питання «звідки»). Тім’яні зони пов’язані із переміщенням джерела звуків; цікавим є той факт, що у лобовій частці кори мавпи виявлені нейрони, які реагують лише на близькі звуки, що, очевидно, мають найбільше значення для виживання тварин. З іншого боку, скроневі частки людини містять вентральні шляхи, які відповідають на питання «що»; нейрони цих шляхів вибірково реагують на голоси. Ці вибіркові до голосів ділянки можуть бути подібні до ділянок зорової кори, що відповідають за розпізнавання обличь (див. Розділ 8).
Подібно до соматичних та зорових кіркових систем слухова система змінюється під впливом набутого досвіду та інших чинників. Збільшення розміру звукосприймальних ділянок кори під впливом слухових подразнень, доповнених високочастотним стимулюванням базальних шляхів переднього мозку, описане у Розділі 7. Свідченням слухової пластичності людини можуть бути результати спостережень за особами, що втратили слух перед набуттям мовних навичок: розглядання письмових мовних символів активує у цих індивідів слухові асоціативні зони за межами первинної слухової кори. Натомість, особи, які у ранньому дитинстві втратили зір, достовірно ліпше локалізують джерело звуку, аніж особи з нормальним зором. Іншими дослідженнями з’ясовано, що діти після досягнення ними шестимісячного віку швидко набувають здатності давати підвищені нейронні відповіді на звуки, специфічні для їхньої рідної мови, тоді як відповіді на неспецифічні звуки поступово зникають.
Музиканти дають додаткові приклади кіркової пластичності. Наприклад, унаслідок постійної активації музичними тонами розмір слухових зон у них збільшений. Окрім того, скрипалі мають дещо змінене соматосенсорне представництво ділянок, на які проектуються пальці, потрібні для гри на скрипці. У музикантів також достовірно більший мозочок порівняно з немузикантами з огляду на набуті навички до точних рухів пальцями.
Частина задньої верхньої скроневої закрутки, відома як скронева площина (рис. 9-16), як звичайно, більша ліворуч, аніж праворуч, особливо це стосується осіб із переважанням правої руки («праворукі»). Ця зона виявилася пов’язаною з мовно-слуховим сприйняттям. Цікавим є спостереження, яке до цього часу не пояснене: ліва скронева площина перевищує свої нормальні розміри у музикантів та осіб з музикальним слухом. Загальні питання кортикальної асиметрії розглянуто у Розділі 16.
Локалізація звуків
Визначення напряму джерела звуку у горизонтальній площині ґрунтується на визначенні різниці часу надходження подразнення до обох вух, наслідком чого є різниця у фазах звукових хвиль з обох боків голови. Має значення також той факт, що звук буде голоснішим з того боку, що ближчий до джерела звуку. Визначення часової різниці, яка може не перевищувати 20 мкс, уважають головним чинником визначення джерела звуку при частотах до 3 000 Гц, тоді як у разі вищих частот вирішальну роль відіграє різниця в силі звуку. Нейрони слухової кори, що отримують подразнення від обох вух, дають максимальну або мінімальну відповідь, коли надходження звукового сигналу до одного вуха відстає на деякий проміжок від часу надходження цього сигналу до іншого вуха. Цей проміжок відрізняється у різних нейронів.
Звуки, джерело яких міститься спереду від слухача, відрізняються від тих, що надходять ззаду, оскільки кожна вушна мушля трохи повернута до переду. Окрім цього, характер відбивання звукових хвиль від поверхні вушної мушлі змінюється зі зміщенням звуку вгору або вниз; зміни звукових хвиль є головним чинником локалізації звуків у вертикальній площині. Локалізація джерела звуків суттєво порушена у разі ушкоджень слухової кори.
Аудіометрія
Гостроту слуху звичайно вимірюють за допомогою аудіометра. Цей прилад слугує джерелом чистих тонів певної частоти, які обстежувана особа сприймає через навушники. Для кожної частоти визначають поріг чутності, значення якого наносять на діаграму у вигляді процентного співвідношення до нормальної чутності. Така діаграма дає об’єктивне відображення ступеня глухоти і якісну характеристику найураженіших діапазонів тональності.
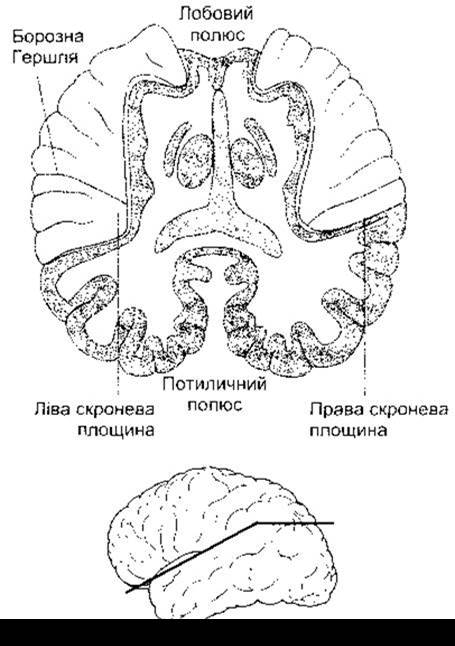
Рис. 9-16. Ліва та права скроневі площини півкуль головного мозку. Горизонтальний зріз уздовж Сильвієвої борозни (орієнтація площини зрізу показана на нижній частині рисунка) (відтворено за дозволом з Kandel ER, Schwartz JH, Jessel TM [editors]: Principles of Neural Science, 3rd ed. McGraw-Hill, 1991).
Глухота
Клінічна глухота може бути зумовлена порушеннями передавання звуків у зовнішньому або середньому вусі (глухота передавання, або кондуктивна), ушкодженнями волоскових клітин або провідних шляхів (сенсоневральна глухота). Ці дві форми патології звукосприйняття розрізняють за допомогою різноманітних простих тестів з камертоном. Три такі тести, названі за прізвищами їхніх авторів, наведені у табл. 9-1. Тести Вебера і Швабаха відображають маскувальний вплив шуму оточення на слуховий поріг.
Причинами глухоти передавання може бути закриття зовнішніх слухових каналів вушною сіркою або сторонніми тілами, ушкодження слухових кісточок, потовщення барабанної перетинки внаслідок повторних інфекцій середнього вуха, надто жорстке прикріплення основи стремінця до овального вікна. Аміноглікозидні антибіотики, такі як стрептоміцин та гентаміцин, закривають механочутливі йонні канали у стереоциліях волоскових клітин і можуть зумовити дегенерацію волоскових клітин, спричинюючи сенсоневральну глухоту та порушення вестибулярних функцій. Руйнування зовнішніх волоскових клітин унаслідок тривалої дії сильного шуму також пов’язане з втратою слуху. Іншими причинами є пухлини присінково-завиткового нерва та мосто-мозочкового кута, ушкодження мозку судинного генезу. Стареча глухота - поступова, пов’язана з віком, втрата слуху, від якої страждає більше третини осіб віком понад 75 років, правдоподібно, зумовлена поступовим відмиранням як волоскових клітин, так і нейронів.
Глухота, спричинена генними мутаціями, вражає близько 0,1 % усіх новонароджених. У 30% випадків вона супроводжуються аномаліями інших систем (синдромна глухота), у решти 70% - це єдиний явний дефект (несиндромна глухота). Є дані про те, що зумовлена мутаціями несиндромна глухота спершу виявляється у дорослих і охоплює до 16% осіб зі значними порушеннями слуху. Протягом останніх років описано багато генних мутацій, які призводять до глухоти. Це не лише розширило уявлення про патофізіологію глухоти, а й дало цінну інформацію про фізіологію слуху. Сьогодні відомо, що для нормального слуху потрібні продукти транскрипції понад 100 генів, і локуси, що можуть зумовити глухоту, описані у складі 19 з 24 хромосом людини.
Цікавим прикладом білків, мутація яких призводить до втрати слуху, може бути конексин 26. Спричинені цим білком дефекти конексонів (див. Розділ 1), імовірно, перешкоджають нормальній рециклізації К+ через підтримувальні клітини (див. рис. 9-9). Глухоту також зумовлюють мутації трьох нем’язових міозинів (див. Розділ 1). До них належить міозин VIІa, що у зв’язаній з актином формі локалізований у відростках волоскових клітин; міозин Іb, який, правдоподібно, є одним із компонентів «адаптаційного мотора», що регулює силу натягу верхівкових з’єднань волосків (див. вище); а також міозин IV, який має суттєве значення для формування нормальних війок. Глухота може бути пов’язана і з мутантними формами а-тектину, одного із головних білків покривної мембрани.
Приклад синдромної глухоти - синдром Пендреда, у разі якого мутація транспортного білка сульфатів зумовлює глухоту та зоб. Іншим прикладом може слугувати синдром
продовженого інтервалу QT, за якого простежується мутація одного з білків К+-каналів, а саме: KVLQT1. У складі клітин судинної смужки цей білок забезпечує підтримання високої К+-концентрації в ендолімфі, тоді як у механізмі серцевих скорочень його роль полягає у забезпеченні нормального QT інтервалу. Особи, гомозиготні за мутантним KVLQT1-геном, глухі і мають схильність до шлуночкових аритмій та раптової смерті, що є найхарактернішими ознаками синдрому продовженого інтервалу QT (див. Розділ 28).