Фізіологія людини - Вільям Ф. Ґанонґ 2002
Функції нервової системи
Стан тривоги, сон та електрична активність мозку
Біоелектричні потенціали кори головного мозку
Біоелектричні реакції, що відбуваються в корі після подразнення органів чуття, можна перевіряти за допомогою електрода, що розташований на певній відстані і має зв’язок з іншим електродом в індиферентній точці. Характерна реакція простежується у тварин під барбітуратною анестезією, яка значно послаблює основну біоелектричну активність. Якщо досліджувальний електрод міститься над первинною зоною сприймання для відповідного відчуття, то поверхнево позитивна хвиля має латентний період тривалістю 5-15 мс. Далі надходить мала негативна хвиля, у цьому разі виникає більший, видовженіший позитивний зубець з латентним періодом 20-80 мс. Першу позитивно-негативну хвильову послідовність називають первинним збудженим потенціалом, другу - вторинною дифузною реакцією.
Первинний збуджений потенціал надзвичайно специфічний з огляду на розташування. Його можна виявити тільки на закінченнях шляхів від конкретного органа чуття. Електрод на поверхні м’якої мозкової оболонки кори фіксує активність на глибині лише до 0,3-0,6 мм. Первинна реакція є швидше негативною, ніж позитивною, якщо її фіксувати мікроелектродом, уведеним вглиб між шарами 2-6 кори головного мозку, а після негативної хвилі в корі настає позитивна. Негативно-позитивна послідовність свідчить про деполяризацію на дендритах і тілах клітин у корі, після якої настає гіперполяризація. Позитивно-негативна хвильова послідовність, записана з поверхні кори, виникає тому, що поверхневі шари кори є позитивними порівняно з внутрішньою негативністю і негативними порівняно з глибокою гіперполяризацією. У тварин чи людей, що не були піддані анестезії, первинний збуджений потенціал значно затінений спонтанною активністю мозку, однак його наявність є наслідком, що виявляється поза середнім рівнем головної біоелектричної активності. Цей потенціал дещо більше дифузійний у тварин, не підданих анестезії, однак порівняно з дифузійною вторинною реакцією добре локалізований.
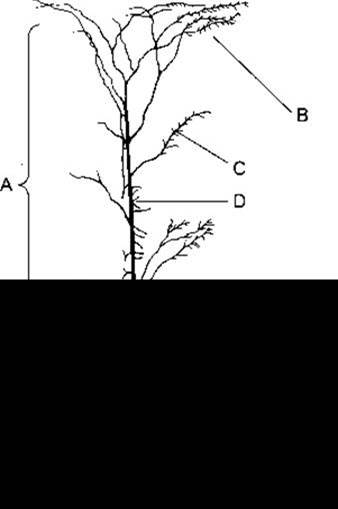
Рис. 11-2. Пірамідальна клітина нової кори, показано поширення нейронів, що закінчуються на ній. А - неспецифічні аференти з ретикулярної формації і таламуса; В - повторювані колатералі аксонів пірамідальної клітини; С - спайкові волокна з ділянок дзеркального відображення у протилежній півкулі; D - специфічні аференти з сенсорних ядер таламуса (відтворено за дозволом з Chow KL, Leiman AL: The structural and functional organization of the neocortex. Neurosci Res Program Bull. 1970; 8: 157).
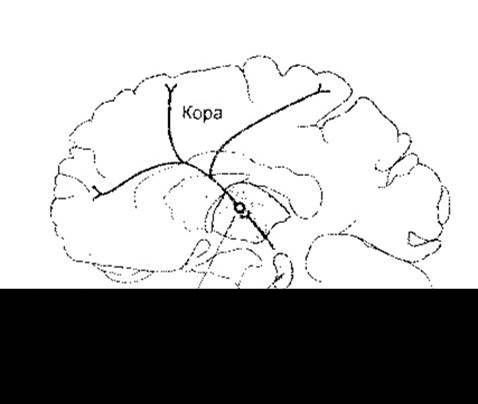
Рис. 11-3. Схематичне зображення висхідної ретикулярної системи у середньому мозку людини, її проекції до внутрішньоламінарних ядер таламуса та вихід внутрішньоламінарних ядер до багатьох частин кори головного мозку. Активування цих ділянок показано за допомогою ПЕТ сканування у разі переходу суб’єкта зі стану розслаблення у випадку неспання до стану концентрації уваги під час виконання завдання (дані з Kinomura S et al: Activation by attention of the human reticular formation and thalamic intralaminar nuclei. Science 1996; 271: 512).
Вторинна поверхнево-позитивна дифузійна реакція, на відміну від первинної, не є чітко локалізованою. Вона з’являється одночасно майже по всій корі головного мозку і відповідає активності в проекціях з серединних та споріднених таламічних ядер.