Фізіологія людини - Вільям Ф. Ґанонґ 2002
Функції нервової системи
Регулювання постави і рухів
Система регулювання постави - Роль довгастого мозку
В експериментальних тварин, у яких унаслідок перетинання стовбура головного мозку на рівні верхнього краю мосту задній і спинний мозок відокремлені від решти ЦНС, головним проявом ушкодження є виражена спастичність мускулатури тіла. Це оперативне втручання називають децеребрацією, а характерну спастичність, що виникає після нього, - децеребраційною ригідністю. Децеребрація спричинює наслідки, що відрізняються від спінального шоку, оскільки ригідність настає одразу ж після перетинання стовбура головного мозку.
Механізм децеребраційної риґідності
Проявом децеребраційної риґідності є спастичність, що виникає внаслідок дифузного посилення рефлексів розтягування. Це явище зумовлене двома факторами: зростанням загальної збудливості пулу мотонейронів і відповідної зміни потенціалів у у-еферентних нейронах.
Супраспінальне регулювання рефлексів розтягування
Ділянки головного мозку, з яких надходять імпульси, що посилюють або гальмують рефлекси розтягування, зображені на рис. 12-6. Ці імпульси, здебільшого, або підвищують, або знижують чутливість м’язових веретен (рис. 12-7). Значного розміру стимулювальна ділянка ретикулярної формації стовбура головного мозку діє спонтанно, очевидно, під впливом аферентних імпульсів, подібно до РАС. Однак меншого розміру ділянка стовбура головного мозку, що інгібує збудження в у-еферентних нейронах, діє не спонтанно, а під впливом імпульсів, які надходять з кори великого мозку і мозочка. Інгібувальна ділянка в базальних ядрах або діє безпосередньо через низхідні шляхи, як це показано на рис. 12-6, або ж стимулює кірковий інгібувальний центр. Імпульси з інгібувальної і стимулювальної ділянок ретикулярної формації надходять донизу в складі латерального канатика спинного мозку. У випадку перетинання стовбура головного мозку на рівні верхнього краю мосту, дві з трьох інгібувальних ділянок, що керують інгібувальним центром ретикулярної формації, залишаються вище перетину і їхній вплив припиняється. Вплив стимулювальної ділянки залишається і, отже, баланс між стимулювальними та інгібувальними імпульсами, що впливають на у-еферентні нейрони, зміщується в бік стимулювання. Потенціал в у-еферентних нейронах збільшується і, як наслідок, рефлекси розтягування стають гіперактивними. Екстирпація мозочка в децеребраційних тварин, і тим самим вилучення ще одної інгібувальної ділянки, призводить до підвищення рівня ригідності. Механізм впливу мозочка складний. Наприклад, деструкція мозочка в людини призводить більше до гіпотонії, ніж до спастичності.
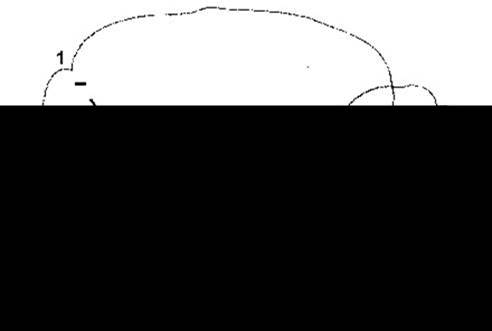
Рис. 12-6. Ділянки головного мозку кота, подразнення яких чинить стимулювальний (знак плюс) або інгібувальний (знак мінус) вплив на рефлекси розтягування: 1 - моторна кора; 2 - базальні ядра; 3 - мозочок; 4 - інгібувальна ділянка ретикулярної формації; 5 - стимулювальна ділянка ретикулярної формації; 6 - присінкові ядра (відтворено за дозволом з Lindsley DB, Schreiner LH, Magoun HW: An electromyographic study of spasticity J Neurophysiol 1949; 12 197).
Присінково-спинномозковий і ще деякі інші низхідні шляхи теж посилюють рефлекси розтягування, спричинюючи ригідність; на відміну від шляхів з ретикулярної формації, вони проходять переважно в складі переднього канатика спинного мозку, і ригідність, зумовлену збудженням у них, не можна зняти шляхом деаферентації м’язів. Це означає, що ця ригідність спричинена безпосереднім впливом на а-мотонейрони, який підвищує їхню збудливість, більше, аніж дією через малу рухову нервову систему, що, звичайно, могла б бути заблокованою шляхом деаферентації.
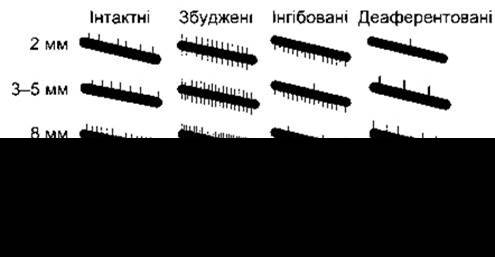
Рис. 12-7. Відображення ізольованих аферентних волокон, що проходять від м’язових веретен, на різного ступеня розтягування м’яза. Числа ліворуч позначають ступінь розтягування. Виступи догори відображають рівень потенціалів дії, виступи донизу - артефакти подразника Запис виконано перед подразненням головного мозку (перший стовпець), під час подразнення ділянок головного мозку, що стимулюють (другий стовпець) або інгібують (третій стовпець) рефлекси розтягування, і після перетинання рухових нервів (четвертий стовпець) (відтворено за дозволом з Eidred Е, Granit R, Merton PA: Supraspinal control of muscle spindles. J Physiol [Lond] 1953; 122:498).
Суть децеребраційної риґідності
У собак і котів спастичність, що виникає внаслідок децеребрації, найбільше позначається на функції м’язів-розгиначів. Шеррінгтон зазначав, що ці м’язи в тварин протидіють земному тяжінню; постава котів і собак після децеребрації, якщо їм надати стоячого положення, є «карикатурою на нормальне стояння».
Досліди з децеребрацією дали змогу також розкрити механізми тонічних статичних рефлексів, що сприяють підтриманню тіла тварини, протидіючи земному тяжінню. Додатковим аргументом щодо правильності інтерпретації цього феномену є дані спостережень над тваринами, що живуть на деревах. Більшість часу ці тварини висять догори ногами на гілках, і в них виникає ригідність м’язів-згиначів. У людей, як і в дослідних котів і собак, справжня децеребраційна ригідність супроводжується підвищенням тонусу м’язів-розгиначів. Очевидно, людина в своїй еволюції не так далеко відійшла від своїх чотириногих предків, щоб мати відмінні особливості в реакції м’язів верхніх кінцівок, навіть незважаючи на те, що основні м’язи, які протидіють тяжінню в положенні стоячи, - це згиначі. Однак децеребраційна ригідність зрідка трапляється в людини, а ушкодження, що спричинюють її, переважно призводять до смерті.
Найчастіше ригідність м’язів-розгиначів нижніх кінцівок і помірне згинання у верхніх кінцівках виникають як ознаки декортикаційної риґідності, зумовленої ушкодженням кори великого мозку, у разі неушкодженого стовбура головного мозку (рис. 12-8).
Тонічні лабіринтні рефлекси
У децеребраційних тварин тип ригідності в кінцівках залежить від положення, яке ці кінцівки займають. Рефлексів випрямлення нема, і тварини перебувають у тому положенні, у якому їх залишити. Якщо тварину покласти на спину, то простежується максимальне розгинання в усіх чотирьох кінцівках, у положенні на будь-якому боці ригідність зменшується, а якщо її покласти ниць, то ригідність стає мінімальною. Ці зміни риґідності - тонічні лабіринтні рефлекси - зумовлені впливом земного тяжіння на отолітові органи і відбуваються через присінково-спинномозкові шляхи. Такий ефект дещо несподіваний з погляду розуміння ролі ригідності в положенні стоячи, і його детальне фізіологічне значення не з’ясоване.
Переконливим доказом важливості вестибулярного апарату для регулювання постави є вплив лабіринтектомії в котів. У нормі кіт легко навчається пересуватись по балці, що обертається. Після односторонньої лабіринтектомії ця здатність зникає, проте після приблизно шести тижнів відновлюється внаслідок адаптивних змін у решті шляхів, що забезпечують регулювання постави. Однак після двосторонньої лабіринтектомії здатність пересуватись по балці, що обертається, не відновлюється.

Рис. 12-8. Декортикаційна риґідність (А-С) і справжня декортикаційна риґідність (D). А - пацієнт, що лежить навзнак з неповернутою головою; В та С - тонічний шийний рефлекс, зумовлений повертанням голови праворуч або ліворуч (відтворено за дозволом з Fulton JF [editor]: Textbook of Physiology, 17th ed. Saunders, 1955)
Тонічні шийні рефлекси
Зміна положення голови стосовно тулуба в децеребраційних тварин спричинює зміни типу ригідності. Якщо голова повернута в один бік, то кінцівки цього ж боку («щелепні кінцівки») випростовуються і стають більш ригідними, тоді як риґідність кінцівок контрлатерального боку зменшується. Це положення характерне і для інтактних тварин, які повертають голову, щоб дивитись убік. Згинання голови спричинює згинання передніх кінцівок і наступне розгинання задніх: виникає постава, характерна для тварини, що заглядає в нору. Розгинання голови спричинює згинання задніх кінцівок і розгинання передніх: виникає постава, характерна для тварини, що споглядає поверх перешкоди. Ці реакції називають тонічними шийними рефлексами. їх ініціюють пропріорецептори внаслідок розтягування м’язів у верхній ділянці шиї, і тривати вони можуть довго.