Молекулярная биотехнология. Принципы и применение - Глик Б., Пастернак Дж. 2002
Молекулярная биотехнология микробиологических систем
Молекулярная генетика человека
Обнаружение и оценка генетического сцепления у человека
До появления в начале 1980-х гг. технологии рекомбинантных ДНК обнаружение и оценка генетического сцепления у человека представляли собой сложную и очень трудоемкую процедуру, которая к тому же обычно оказывалась безуспешной. При этом исследователи сталкивались с целым рядом проблем. Во-первых, генетический статус родителей обычно бывает неизвестен, что затрудняет разграничение рекомбинантных и нерекомбинантных потомков. Во-вторых, немногочисленность большинства семей снижает статистическую достоверность полученных результатов. Наличие у мужчин одной Х-хромосомы значительно облегчает оценку генетических расстояний между генными локусами. В данном случае все аллели генов, расположенных на Х-хромосоме, проявляются фенотипически. Сыновья женщин, дигетерозиготных по Х-сцепленным локусам, получают рекомбинантную или нерекомбинантную Х-хромосому. Если фаза, в которой аллели двух генных локусов находятся у матерей, известна, то среди сыновей легко установить рекомбинантные и нерекомбинантные типы. Генотип із отцов в данном случае не имеют значения, поскольку сыновья наследуют только материнскую Х-хромосому. Иногда фазу аллелей у дигетерозиготной матери можно установить исходя из фенотипа ее отца. Например, если у отца матери (деда) два Х-сцепленных признака рецессивны, а у нее самой — доминантны, то мать дигетерозиготна, а рассматриваемые аллели находятся в цис-фазе, т. е. АВ/аb (рис. 20.8). Этот метод обнаружения сцепления основан на подсчете двухлокусных фенотипов у сыновей большого числа дигетерозиготных женщин с известной фазой аллелей. В этом случае доля хромосом, рекомбинантных по двум специфическим генным локусам (рекомбинационный индекс), будет равна сумме рекомбинантных хромосом (R), деленной на общее число хромосом — рекомбинантных и нерекомбинантных (NR):
![]()
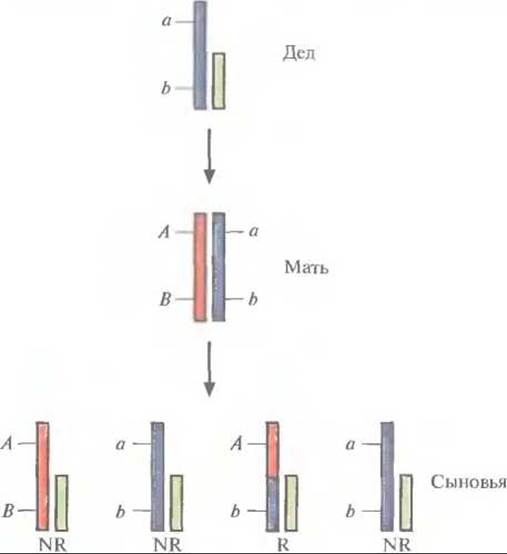
Рис. 20.8. Картирование X хромосомы. В этом случае генетическая фаза двух или большего числа Х-сцепленных локусов у дочери (Мать) устанавливается на основании данных о Х-сцепленных аллелях ее отца (Дед). Эту информацию в свою очередь используют для определения, какие из ее сыновей (Сыновья) получили рекомбинантную (R) и нерекомбинантную (NR) хромосому. В данном примере дед несет два рецессивных гена в локусах А и В Х-хромосомы, его дочь дигетерозиготна, а рассматриваемые аллели находятся у нее в цис-фазе. На Х-хромосоме показаны аллели локусов А и В, Y-хромосома изображена в виде более короткой полоски.
Однако данный подход имеет ряд недостатков. Во-первых, не всегда можно определить генотип деда, а следовательно, фаза, в которой находятся аллели у предположительно дигетерозиготной матери, остается неизвестной. Во-вторых, не все матери в большой выборке семей будут гетерозиготны по одним и тем же двум локусам. Несмотря на все усилия, до 1980-х гг. не удавалось построить достаточно протяженную однозначную карту сцепления Х-хромосомы человека, основанную на подсчете рекомбинантных и нерекомбинантных хромосом. В то время было известно всего несколько локусов и слишком мало аллелей было идентифицировано.
Анализ сцепления методом максимального правдоподобия: логарифм соотношения шансов (лод-балл)
Кроме метода, в котором определяется частота рекомбинаций между двумя локусами на основании прямого подсчета рекомбинантных и нерекомбинантных хромосом, необходимо было разработать более общий, непрямой метод, который: 1) мог бы строго различать независимое распределение и сцепление; 2) не обязательно опирался бы на данные о фазе аллелей дигетерозиготных родителей; 3) мог суммировать информацию, полученную от большого количества различных семей; 4) позволял оценить рекомбинационный индекс в том случае, когда сцепление обнаружено. Такой метод, широко использующийся в настоящее время, был создан в 1955 г. Мортоном.
При изучении сцепления рекомбинационный индекс обозначается греческой буквой тета (θ). В методе Мортона сравнивается вероятность L(θ) того, что у братьев и сестер (сибсов) два локуса сцеплены (т. е. локализованы на одной хромосоме и находятся близко друг от друга), с вероятностью L(0.50) того, что два локуса не сцеплены (т. е. находятся на разных хромосомах или далеко друг от друга в пределах одной хромосомы), для любого рекомбинационного индекса 0. В случае сцепления, поскольку рекомбинационный индекс неизвестен, он может принимать любое значение в интервале от 0 до 0,5 (0 ≤ θ < 0,50). Если же два локуса распределяются независимо, то 0 = 0,50 по определению. Другими словами, в том случае, когда половина гамет, полученных от дигетерозиготного родителя, содержит новые генетические комбинации, два локуса находятся либо на негомологичных хромосомах, либо настолько далеко друг от друга на одной хромосоме, что это выглядит так, будто они расположены на разных хромосомах. Следовательно, если L(θ) = L(0,50), то два локуса не сцеплены. Десятичный логарифм отношения этих двух вероятностей, т.е. log [L(θ )/L((0,50)], представляет собой логарифм соотношения шансов (log-of-odds ratio), называемый лод-баллом (LOD). Лодд-балл обозначают буквой Z; Z(θ) — это лод-балл для данного значения θ, где 0 ≤ θ < 0,50.
L(θ) можно определить, если известна вероятность получения конкретного сочетания рекомбинантных и нерекомбинантных хромосом для сибсов каждой изучаемой семьи. Вероятность того, что потомки получат от дигетерозиготного родителя нерекомбинантную хромосому, равна 1/2(1—θ) + 1/2(1—θ), или 1—θ, а вероятность того, что они получат рекомбинантную хромосому, — 1/2θ + 1/2θ, или θ. Например, в семье с пятью детьми вероятность для каждого из них получить нерекомбинантную хромосому от дигетерозиготного родителя составляет К(1-θ)5, где (1—θ) — вероятность получения нерекомбинантной хромосомы, показатель степени 5 — число сибсов с нерекомбинантной хромосомой, точнее, число нерекомбинантных хромосом у сибсов, К — коэффициент. Если все хромосомы одинаковы, т. е. все нерекомбинантные или все рекомбинантные, то К = 1 (т. е. 5!/5!0!, или n!/n!0!, где n — число сибсов в данной семье). В семье с четырьмя детьми вероятность того, что все они получат рекомбинантную хромосому от дигетерозиготного родителя, составляет θ4. Далее, вероятность того, что в семье с девятью детьми пять получат нерекомбинантные хромосомы и четыре — рекомбинантные, равна K(1-θ)5(θ)4, где К= 126, т. е. 9!/5!4!. Лод-балл выражается как отношение величин, имеющих одинаковые коэффициенты. Эти коэффициенты, стоящие в числителе и знаменателе, сокращаются, а потому при анализе сцепления не учитываются.
Проиллюстрируем подсчет лод-балла на примере сибсов одной семьи (рис. 20.9). Обозначения В и О на рис. 20.9 соответствуют аллелям АВО* В и АВO*O групп крови системы АВО. Закрашенными символами обозначено аутосомно-доминантное заболевание с полной пенетрантностью — наследственный онихоартроз (NPS, nail-patella syndrom). Основные признаки NPS — нарушение роста ногтей на пальцах рук и ног и редукция или отсутствие надколенника. Ген NPS обозначается NPS1, а его рецессивный («нормальный») и доминантный («патологический») аллели — NPS1*N и NPSl*D соответственно. NPS представляет собой подходящий для изучения сцепления признак, так как он рано диагностируется, не влияет на жизнеспособность и репродуктивную функцию и присутствует при рождении.
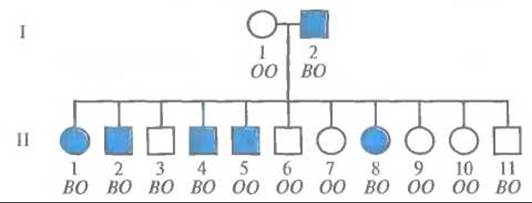
Рис. 20.9. Наследование генов онихоартроза и генов групп крови системы АВО. Закрашенными символами обозначены лица с наследственным онихоартрозом, незакрашенными — лица, у которых признаки данного заболевания отсутствуют. Буквы под каждым символом обозначают аллели групп крови системы АВО (использованы сокращенные обозначения: О соответствует АВO*O, В — АВО*В).
Отец 1-2 (рис. 20.9) гетерозиготен по локусу NPS, поскольку среди его детей есть как больные, так и здоровые. Он гетерозиготен и по локусу АВО (АВО*В/АВO*O), так как у его детей встречаются фенотипы О и В, а генотип его супруги (1-1) — АВO*O/АВO*O. Следовательно, отец дигетерозиготен по этим двум аутосомным локусам (NPS1*N/NPS1*D; АВO*В/АВO*O). Если локусы АВО и NPS1 сцеплены, то фаза, в которой находятся их аллели у отца, неизвестна (состояние с неизвестной фазой). Она может быть как АВО*В NPS1*D/ABO*O NPS1*N (фаза 1), так и АВО*В NPS1 *N/ABO*O NPS1*D (фаза 2). Или, в сокращенном виде, В D/O N (фаза 1) или В N/О D (фаза 2).
Если предположить, что локусы АВО и NPS сцеплены и их аллели у отца находятся в фазе 1 (АВО*В NPS1 *D/ABO*D NPS1*N), то дети II-1, 11-2, II-4, И-6, И-7, II-8, II-9 и II-10 получили от него нерекомбинантную хромосому АВО*В NP1*D или АВО*O NPS1*N (рис. 20.10). Все дети получили от матери (I-!) хромосому АВО*O NPS1*N, поскольку она гомозиготна по двум локусам (АВО*O NPS1*N/ABO*O NPS1*N). В данном случае генетический вклад матери известен и не влияет на анализ сцепления. Исходя из того, что рассматриваемые аллели у отца находятся в фазе 1, каждый из его детей II-З, II-5 и II-11 получил рекомбинантную хромосому. Следовательно, вероятность такого сочетания нерекомбинантных и рекомбинантных хромосом для данной семьи равна (1-θ)8(θ)3.
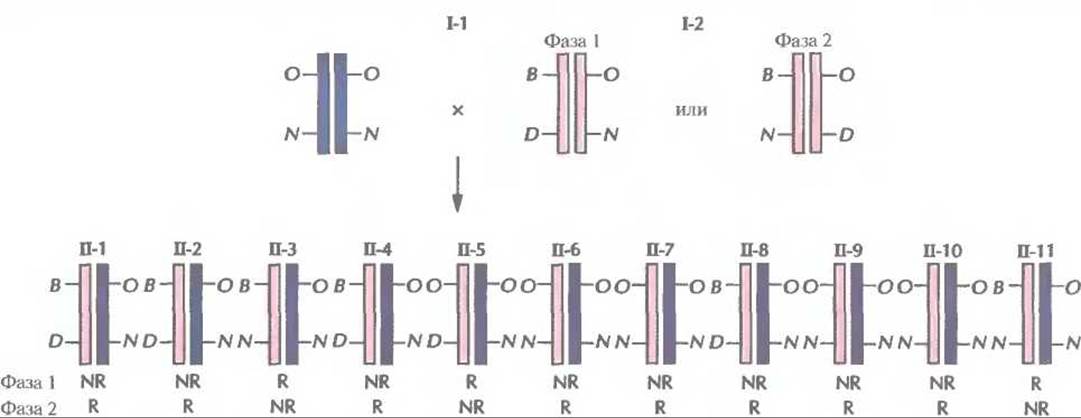
Рис. 20.10. Генетическая организация аллелей генов онихоартроза и групп крови АBО у членов родословной, приведенной на рис. 20.9, при условии сцепления этих двух локусов. Использованы сокращенные обозначения аллелей групп крови системы АВО: О соответствует АВO*O, а В — АВО*В. Рецессивный («нормальный») и доминантный («патологический») аллели локуса наследственного онихоартроза обозначены N и D соответственно. Генотип отца (1-2) может находиться в любой из двух фаз (фаза I, фаза 2). Хромосомы отца и хромосомы, унаследованные от него детьми, выделены синим цветом, хромосомы матери (1-1) и хромосомы, унаследованные от нее, — светло-коричневым. Отмечено, какие из хромосом, полученных от отца, являются нерекомбинантными (NR) или рекомбинантными (R) для фазы I и фазы 2.
Рассматриваемые аллели у отца с такой же вероятностью могут находиться в фазе 2, т. е. АВО*В NPS1 *N/AВO*O NPS1*D. Тогда дети II-3, II-5 и II-11 получили от него нерекомбинантные хромосомы, а каждый из оставшихся детей унаследовал рекомбинантную хромосому (рис. 20.10). Вероятность такой комбинации для данной семьи равна (1—θ)3(θ)8.
Поскольку для генотипа отца обе фазы равновероятны, общая вероятность L(θ) наблюдаемой в родословной комбинации хромосом у его детей равна 1/2( 1—θ)8(θ)3 + 1/2(1—θ)3(θ)8. Далее находят значение данного выражения для разных θ. Обычно используют следующий набор значений θ : 0; 0,001; 0,05; 0,10; 0,2; 0,3; 0,4 и 0,50, а если нет ограничений во времени, можно взять весь спектр значений θ от 0 до 0,50. Затем вычисляют логарифм отношения вероятности для каждого θ, кроме 0,50, к вероятности для θ = 0,50. Например, для θ = 0,10 отношение L(0,10)/L(0,50) равно
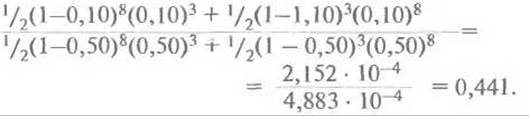
Десятичный логарифм 0,441 равен —0,356; это и есть лод-балл для данного отношения. Другими словами, Z(0,10) = —0,356.
Если фаза, в которой находятся рассматриваемые аллели у отца, известна, то и значение вероятности L(θ) для данной семьи тоже будет известно. Например, если для генотипа 1-2 имеет место фаза 1 (АВО*В NPS1*D/AB0*0 NPS1*N), то, как отмечалось выше, вероятность L(θ) для данной семьи будет равна (1—θ)8(θ)3, и Z(θ = 0,10) составит

Если же для генотипа 1-2 имеет место фаза 2 (ABO*ВNPS1 *N/АВО*ONPS1*D, то Z(0,10) будет равен
![]()
Для состояния с неизвестной фазой значения Z для родословной, приведенной на рис. 20.9, варьируют от —5,993 при θ = 0,001 до +0,029 при θ = 0,45 (табл. 20.1). Если сибсы получили хотя бы одну рекомбинантную хромосому и θ = 0, то Z = -∞. Как видно из табл. 20.1, лод-балл максимален (Zmax) при в, близком к 0,30. Проведя дополнительные расчеты для в от 0,20 до 0,40, получим, что Zmax = +0,214 при 0= 0,276.
Таблица 20.1. Значения Z при разных θ для родословной, приведенной на рис. 20.9, в случае состояния с неизвестной фазой
|
θ |
0 |
0,001 |
0,01 |
0,05 |
0,10 |
0,20 |
0.30 |
0,40 |
0,45 |
|
Z |
-∞ |
-5,993 |
-3,025 |
-1,071 |
-0,356 |
0,138 |
0,209 |
0,095 |
0,029 |
Значение Zmax = +0,214 не позволяет с уверенностью говорить о сцеплении локусов АBО и NPS1. Условились, что два аутосомных локуса могут считаться сцепленными только в том случае, если значение максимального лод-балла большее или равно +3,000: вероятность сцепления в этом случае составляет 1000 к 1 или выше. В случае Х-cцепленных генов, заведомо находящихся на одной хромосоме, значение Zmax, при котором можно говорить о сцеплении, больше или равно +2,000; это соответствует шансам в пользу сцепления 100 к 1 или выше. Если Z= -2,000, то сцепление двух локусов исключается, поскольку в этом случае в пользу сцепления существует лишь 1 шанс из 100.
Чтобы выявить сцепление, необходимо подсчитать Z-балл при разных в для разных семей и найти максимальное его значение. Преобразование отношения правдоподобий для каждой из семей в десятичный логарифм позволяет суммировать полученные Z(∞). Для определения сцепления локусов АBО и NPS1 было проанализировано 25 родословных, в том числе несколько с большим количеством детей, и получено значение Z(0,10) = +31,235 (табл. 20.2); это больше, чем +3,000, следовательно, два указанных локуса сцеплены.
Значение в, при котором Z достигает максимума, дает грубую оценку рекомбинационного индекса для двух сцепленных локусов. В первой работе по определению сцепления локусов АВО и NPS1 точное значение Zmax не определялось, но Z-балл при θ = 0,10 был наибольшим из всех Z, подсчитанных для разных 0, из чего был сделан вывод, что, по-видимому, расстояние между этими двумя локусами составляет примерно 10 сМ. Необходимо подчеркнуть, что при анализе разных родословных с NPS обнаружилось, что с локусом NPS1 сцеплены разные аллели системы АВО. Другими словами, не существует специфического сцепления между конкретным аллелем системы АВО и локусом NPS1. Пока не доказано обратное, можно говорить только о генетическом сцеплении между локусами, а не между определенными аллелями. Следует также отметить, что метод подсчета лод-балла не позволяет определить аутосомную локализацию двух сцепленных локусов. Как мы увидим, для того чтобы установить, что локусы АВО и NPS1 расположены на длинном плече (q) хромосомы 9 между районами 34 и 34.2, т. е. 9q34—9q34.2, потребовались дополнительные исследования.
Таблица 20.2. Суммарные значения Z при разных ∞ для локусов ARO и NPSI1)
|
θ |
0,05 |
0,10 |
0,15 |
0,20 |
0,25 |
0,30 |
0,40 |
|
Z |
28,159 |
31,235 |
30,405 |
27,756 |
23,983 |
19,434 |
9,048 |
1) Из работы Renwick, Schulze, Ann. Hum. Genet. 28:379—392, 1965, с изменениями.