Молекулярная биология клетки - Том 1 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
Введение в биологию клетки
Макромолекулы: структура, форма и информационные функции
Структура белка
Клетки в значительной степени состоят из белков, на долю которых приходится более половины их сухого вещества (см. табл. 3-1). Белки определяют структуру и форму клетки; кроме того, они служат инструментами молекулярного узнавания и катализа. ДНК, хотя и содержит всю необходимую для построения клетки информацию, оказывает незначительное прямое воздействие на клеточные процессы. Например, ген гемоглобина сам не переносит кислород: это свойство белка, кодируемого им. Используя компьютерную терминологию, можно сказать, что ДНК и мРНК представляют собой «программное обеспечение» - инструкции, полученные клеткой от родительской клетки. Белки и молекулы каталитических РНК составляют «аппаратное обеспечение» - физические механизмы, осуществляющие хранящуюся в памяти программу.
ДНК и РНК представляют собой цепи, построенные из нуклеотидов, химически очень похожих друг на друга. Напротив, молекулы белков собраны из 20 очень разных аминокислот, каждая из которых обладает ярко выраженной химической индивидуальностью. Это разнообразие лежит в основе необычайной универсальности химических свойств различных белков, и, по-видимому, эволюция выбрала именно белки, а не молекулы РНК в качестве катализаторов большинства реакций в клетке.
3.3.1. Форма белковой молекулы определяется ее аминокислотной последовательностью [21]
В длинной полипептидной цепи возможно свободное вращение атомов вокруг многих связей, что делает остов белковой молекулы очень гибким. Поэтому любая белковая молекула в принципе может принимать почти бесконечно большое число различных форм (конформаций). Однако большинство полипептидных цепей существуют лишь в одной из этих конформаций, определяемой последовательностью аминокислот. Это обусловлено тем, что боковые группы аминокислот взаимодействуют друг с другом и с водой с образованием слабых нековалентных связей (см. схему 3-1). В этом случае соответствующие боковые группы оказываются в ключевых местах цепи, между ними образуются сильные связи, что делает определенную конформацию очень стабильной.
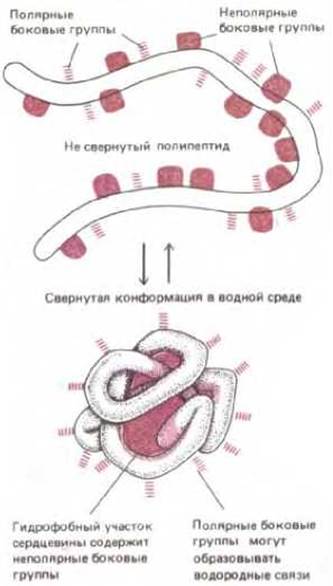
Рис. 3-22. Схематически показано, как белок свертывается в глобулу. Полярные боковые группы аминокислот стремятся расположиться на наружной поверхности белка, где они могут взаимодействовать с водой. Неполярные боковые группы аминокислот расположены внутри, где образуют «спрятанное» от воды гидрофобное «ядро».
Полипептидная цепь большинства белков самопроизвольно сворачивается с образованием правильной конформаций. При обработке определенными агентами белок можно развернуть, или денатурировать; при прекращении действия денатурирующего агента белок обычно самопроизвольно возвращается к исходной конформаций. Это указывает на то, что вся необходимая информация для определения формы белка содержится в самой последовательности аминокислот.
Одним из важнейших факторов, направляющих свертывание полипептидной цепи, является расположение полярных и неполярных боковых групп. Многочисленные гидрофобные боковые группы стремятся собраться внутри белковой молекулы, что позволяет им избежать контакта с водным окружением (точно так же сливаются механически диспергированные в воде капельки масла). В то же время все полярные группы стремятся, наоборот, расположиться на поверхности молекулы белка, где они могут взаимодействовать с водой и другими полярными группами (рис. 3-22). Именно таким путем происходит спаривание почти всех полярных групп, оказывающихся внутри белковой глобулы. Таким образом, водородные связи играют главную роль во взаимодействии разных участков одной полипептидной цепи в свернутой молекуле белка; кроме того, они имеют исключительно важное значение для многих взаимодействий, происходящих на поверхности белковых молекул (рис. 3-23).
Секретируемые белки, или белки клеточной поверхности, часто образуют дополнительные ковалентные связи между разными участками одной и той же полипептидной цепи. Например, образование дисульфидных связей (называемых также —S—S-мостиками) между двумя SH-группами цистеина, оказавшимися по соседству в свернутой полипептидной цепи, стабилизирует пространственную структуру внеклеточных белков (рис. 3-24). Для правильного свертывания белков эти связи не нужны, поскольку оно происходит нормально в присутствии восстанавливающих агентов, препятствующих образованию —S—S-мостиков. В самом деле, —S—S-мостики образуются редко (если образуются вообще) у белковых молекул в цитозоле, где высока концентрация агентов, восстанавливающих SH-группы и разрушающих такие мостики (см. разд. 8.6.11).
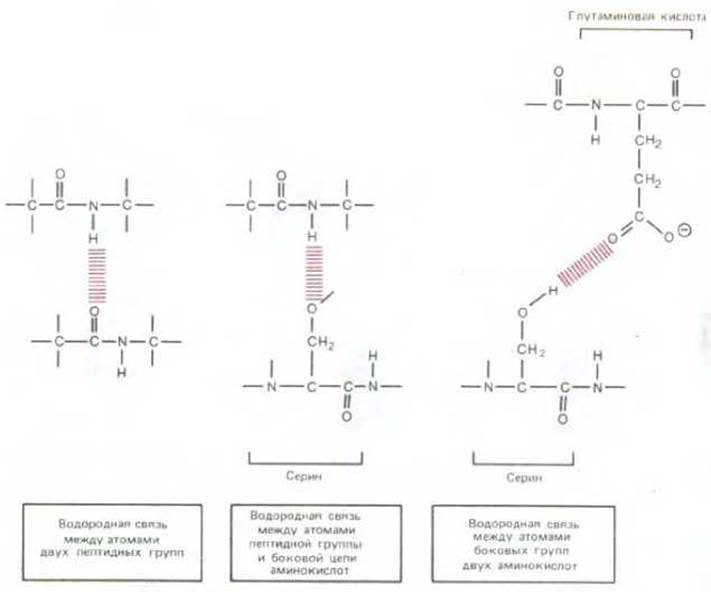
Рис. 3-23. Водородные связи (выделены цветом), которые могут образовываться между аминокислотами в белках. Пептидные связи обозначены серым.
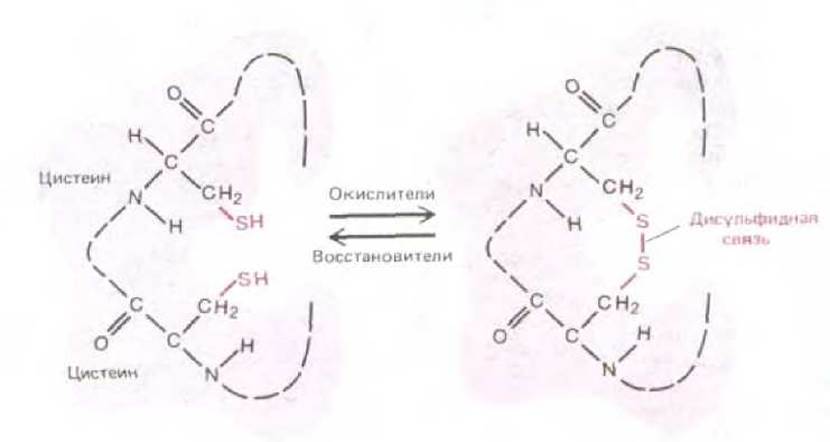
Рис. 3-24. Образование ковалентной дисульфидной связи между соседними остатками цистеина белка.
Общий результат всех индивидуальных взаимодействий аминокислот состоит в том, что большинство молекул белка спонтанно принимает характерную для них конформацию: обычно компактную глобулярную, но изредка и вытянутую фибриллярную. Сердцевина глобулы образована плотно упакованными, почти как в кристалле, гидрофобными боковыми группами, а полярные боковые группы формируют сложную и нерегулярную наружную поверхность. Специфичность связывания белка с малыми молекулами и другими макромолекулярными поверхностями определяется расположением и химическими свойствами различных атомов на этой сложной поверхности (см. ниже). С химической точки зрения белки - наиболее сложные из известных молекул.
3.3.2. Одни и те же способы укладки цепи постоянно повторяются в разных белках [22]
Хотя аминокислотная последовательность полипептидной цепи и содержит всю необходимую для ее свертывания информацию, мы до сих пор не знаем, как эту информацию прочесть, чтобы по последовательности детально предсказать пространственную структуру белка. В результате нативную конформацию белка можно определить лишь с помощью очень трудоемкого метода рентгеноструктурного анализа белковых кристаллов. Этим методом к настоящему времени полностью проанализировано более 100 белков. Специфическая конформация каждого из них столь сложна, что для ее детального описания потребовалась бы целая глава.
При сравнении пространственной структуры различных белков выяснилось, что, хотя конформация каждого белка уникальна, несколько способов укладки цепи постоянно повторяются в отдельных частях макромолекул. Особенно часто встречаются два способа укладки, поскольку они обусловлены регулярным образованием водородных связей между самими пептидными группами, а не уникальными взаимодействиями боковых цепей. Оба способа были правильно предсказаны в 1951 г. с помощью моделей, основанных на результатах рентгеноструктурного анализа шелка и волос. Сейчас эти периодические структуры называют ß-кладчатым слоем и a-спиралью. В ß-складчатой конформации находится белок шелка фиброин, а-спираль обнаружена в а-кератине - белке кожи и ее производных (волосах, ногтях и перьях).
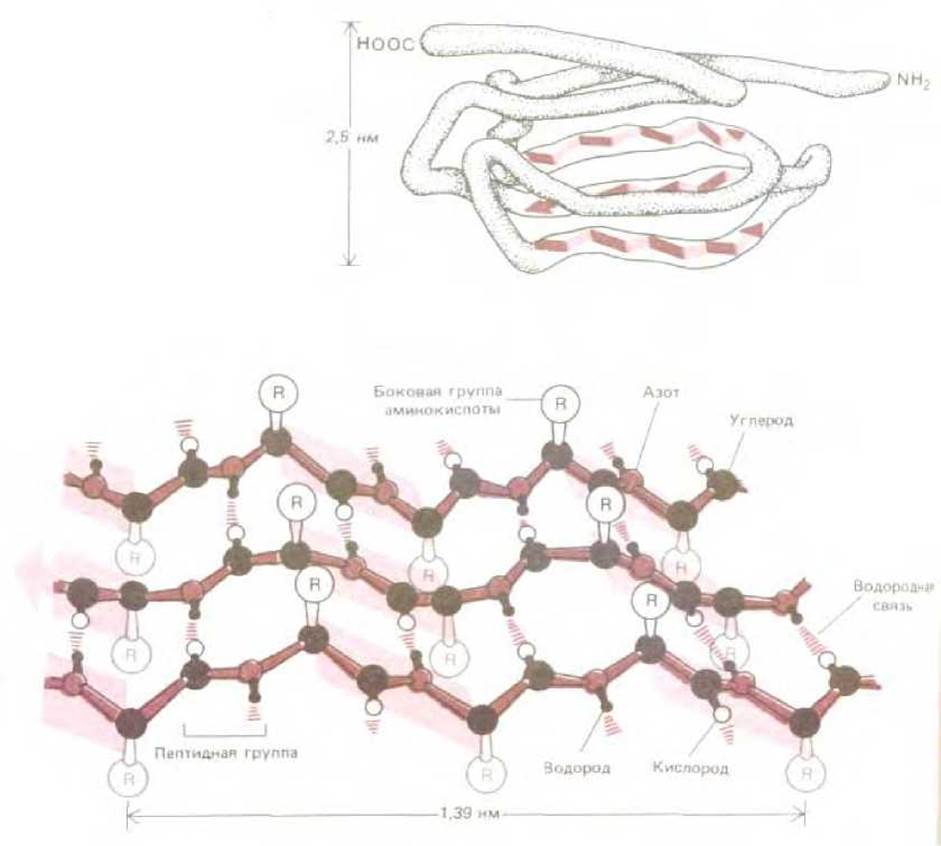
Рис. 3-25. ß-Слой - это обычная структура участков глобулярных белков. Сверху показан включающий 115 аминокислот домен молекулы иммуноглобулина. Он состоит из двух ß-слоев, уложенных наподобие сандвича, один из которых выделен цветом. Внизу более детально изображен совершенный антипараллельный ß-слой. Обратите внимание на то, что каждая пептидная группа образует водородные связи с соседними пептидными группами. ß-Слои, встречающиеся в глобулярных белках, обычно несколько менее регулярны, чем показанная здесь структура; часто ß-слои оказываются слабо скрученными (см. рис. 3-27).
Структура ß -складчатого слоя составляет существенную часть сердцевины (core) большинства (хотя и не всех) глобулярных белков.
На рис. 3-25 для примера показана часть молекулы антитела; антипараллельный ß - слой этой молекулы образован в результате многократного изгибания полипептидной цепи на 180°, так что направление каждого прямого участка цепи противоположно направлению ближайших соседних участков. Такая структура обладает высокой прочностью, обусловленной образованием водородных связей между пептидными группами соседних участков цепи. Поэтому антипараллельный ß -слой часто служит каркасом, на котором собирается глобулярный белок.
а-Спираль образуется при закручивании полипептидной цепи вокруг себя с образованием жесткого цилиндра, в котором каждая пептидная группа связывается водородными связями с ближайшими пептидными группами цепи. Многие глобулярные белки содержат короткие участки таких а-спиралей (рис. 3-26); участки трансмембранного белка, который проходит сквозь липидный бислой, также почти всегда являются а-спиралями в силу сжатия, испытываемого им со стороны гидрофобного липидного окружения (см. разд. 6.2.1). В водной среде изолированная а-спираль обычно неустойчива. Однако две одинаковые а-спирали, имеющие повторяющиеся участки неполярных групп цепей, могут последовательно обвиваться вокруг друг друга с образованием чрезвычайно устойчивой структуры. Такие длинные стержневидные структуры обнаружены во многих фибриллярных белках, в частности во внутри-клеточных волокнах а-кератина, обусловливающих прочность кожи. Пространственные модели а-спирали и ß-складчатого слоя белков с боковыми группами и без них представлены на рис. 3-27.
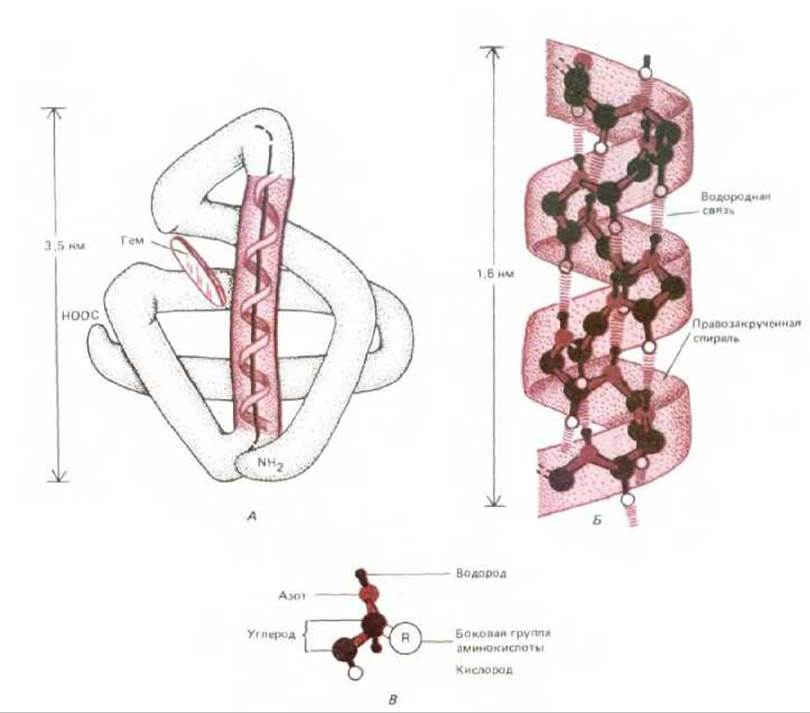
Рис. 3-26. а-Спираль - еще одна общая структура, обычно образующаяся в отдельных участках полипептидной цепи белков. А. Показана переносящая кислород молекула миоглобина (длиной 153 аминокислоты); один из а-спиральиых участков выделен цветом. Б. Детальное изображение совершенной а-спирали. Как и в ß-слое, каждая пептидная группа связана с соседними пептидными группами водородными связями. В. Атомы в аминокислотном остатке. Заметим, что в Б боковые группы аминокислот для упрощения опущены (они расположены на наружной поверхности спирали. В они обозначены как R на атоме а-углерода каждой аминокислоты (см. также рис. 3-27).

Рис. 3-27. Пространственные модели а-спирали и ß-Слоя. Слева структуры показаны без боковых групп аминокислот, справа - с боковыми группами. А. а-Спираль (часть структуры миоглобина). Б. Участок ß-слоя (часть структуры домена иммуноглобулина). На фотографиях слева каждая поверхность цепи представлена только одним черным атомом (группы R на рис. 3-25 и рис. 3-26); вся поверхность цепи показана справа. (С любезного разрешения Richard J. Feldmann.)
3.3.3. Молекулы белков характеризуются чрезвычайным разнообразием [23]
Различия в природе боковых групп аминокислот обусловливает замечательное разнообразие возможных типов пространственной структуры белков. Рассмотрим в качестве примера крайних случаев два типа белков, секретируемых клетками соединительной ткани, - коллаген и эластин, которые относятся к белкам внеклеточного матрикса. В коллагене три отдельные полипептидные цепи, богатые пролином и содержащие в каждом третьем положении глицин, закручены одна вокруг другой и образуют тройную спираль (см. разд. 14.2.6). Эти молекулы коллагена в свою очередь упаковываются в волокна, в которых соседние молекулы скреплены ковалентными сшивками между соседними лизиновыми остатками. В результате формируются волокна, способные выдерживать исключительно большую нагрузку (рис. 3-28).
Другой предельный случай - эластин, в котором относительно рыхлые и неструктурированные полипептидные цепи образуют благодаря ковалентным сшивкам резиноподобную эластичную сеть, которая дает возможность таким тканям, как артерии и легкие, деформироваться и растягиваться, не причиняя себе вреда. Как показано на рис. 3-29, эластичность обусловлена способностью индивидуальных молекул обратимо разворачиваться под действием растягивающего усилия.
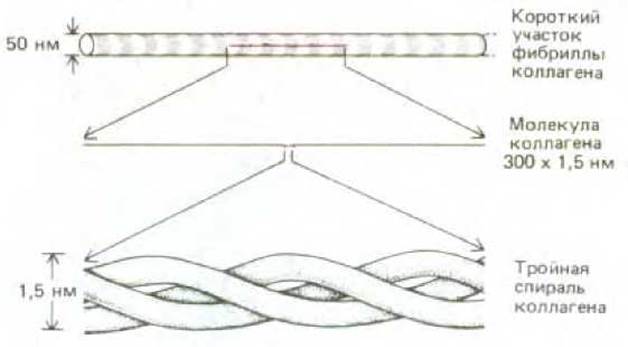
Рис. 3-28. Молекула коллагена - это тройная спираль, образованная тремя вытянутыми белковыми цепями. Множество сшитых вместе стержнеобразных молекул коллагена образует прочные нерастяжимые коллагеновые фибриллы (вверху), которые обладают прочностью на растяжение, сравнимой с прочностью стали.
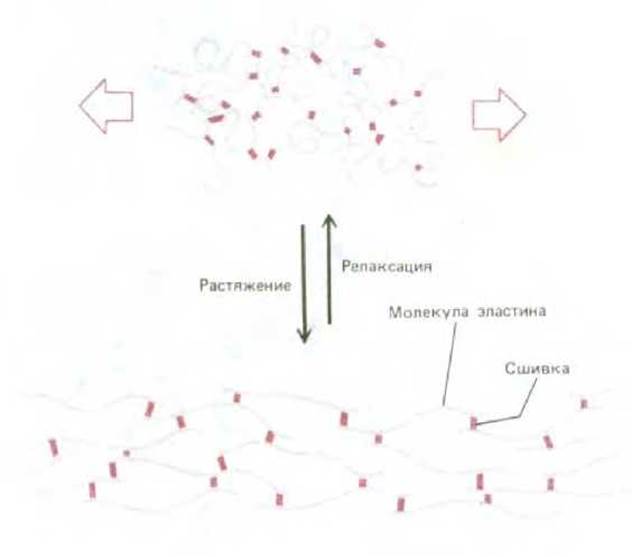
Рис. 3-29. Эластин состоит из полипептидных цепей, образующих благодаря поперечным сшивкам, растяжимые волокна. При растяжении каждая молекула эластина разворачивается, приобретая более протяженную конформацию. Разительный контраст между физическими свойствами эластина и коллагена обусловлен большими различиями в их аминокислотных последовательностях.
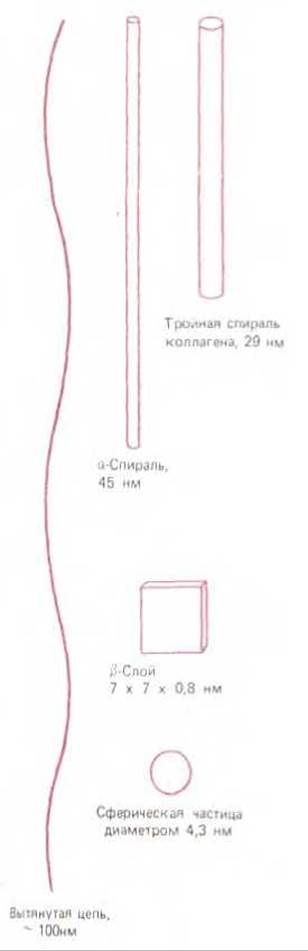
Рис. 3-30. Возможные размеры и форма молекулы белка из 300 аминокислот. Конкретная структура определяется последовательностью аминокислот (D. Е. Metzler, Biochemistry, New York; Academic Press, 1977; печатается с изменениями).
Примечательно, что одна и та же химическая структура - аминокислотная цепь - может приобретать самую различную конформацию. Назовем, например, резиноподобный эластин, похожий на стальной трос коллаген, разнообразные глобулярные белки - ферменты, очень различающиеся по форме своей каталитической поверхности. На рис. 3-30 показано, сколь различную форму может в принципе принимать полипептидная цепь длиной в 300 аминокислот. Реальная конформация, как мы уже отметили, полностью зависит от последовательности аминокислот.
3.3.4. Белки имеют различные уровни пространственной организации [24]
Рассматривая структуру белка, полезно различать разные уровни его пространственной организации. Аминокислотную последовательность называют первичной структурой белка. Регулярные водородные связи по всей длине непрерывной полипептидной цепи приводят к образованию а-спиралей и ß-слоев, которые представляют собой вторичную структуру белка. Некоторые комбинации а-спиралей и ß-слоев, упакованные вместе, формируют компактно уложенные глобулярные единицы, каждая из которых носит название белкового домена. Домены обычно состоят из отрезков полипептидной цепи, содержащих от 50 до 350 аминокислот; по-видимому, они являются теми модульными единицами, из которых строятся белки (см. ниже). Маленькие белки могут содержать только один домен, более крупные белки состоят из нескольких доменов, связанных сравнительно открытыми участками полипептидной цепи. Наконец, отдельные полипептиды могут служить субъединицами для формирования более крупных молекул, часто называемых белковыми агрегатами, или белковыми комплексами. В таких комплексах субъединицы связаны друг с другом большим числом слабых нековалентных взаимодействий (см. разд. 3.1.1), у внеклеточных белков эти взаимодействия часто стабилизированы дисульфидными связями.
Пространственную структуру белка можно проиллюстрировать несколькими способами. Рассмотрим, например, необычно маленький белок - основной ингибитор трипсина поджелудочной железы, который содержит 58 аминокислотных остатков, упакованных в один домен. Этот белок можно представить в виде стереопары, показывающей все его неводородные атомы (рис. 3-31, А), или в виде тщательно выполненной трехмерной модели, где опущены многие детали (рис. 3-31, Б). Белок можно изобразить и более схематично, без боковых групп и атомов, чтобы сфокусировать внимание на последовательности основной полипептидной цепи (рис. 3-31, В, Г и Д). Такие схематические представления очень важны для выявления структуры белков, которые обычно крупнее, чем основной ингибитор трипсина, так как они дают возможность проследить за нерегулярным расположением полипептидной цепи внутри каждого домена (рис. 3-32).
На рис. 3-33 показано, как структура большого белка может быть сведена к разным уровням организации, каждый из которых иерархическим образом строится из предыдущих. Эти уровни возрастающей структурной организации, возможно, соответствуют стадиям свертывания новосинтезированного белка в конечную нативную структуру внутри клетки.
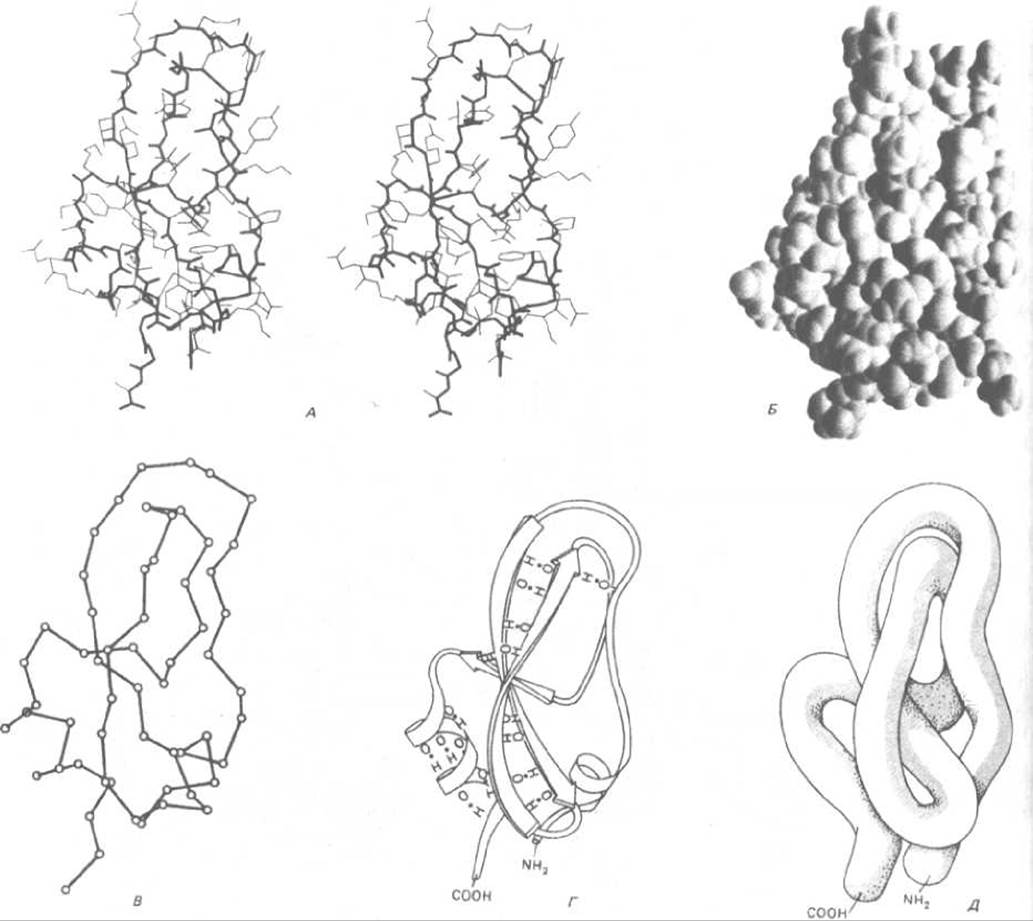
Рис. 3-31. Пространственная конформация малого белка основного ингибитора трипсина поджелудочной железы в пяти обычно использующихся вариантах изображения. А. Стереопара, показывающая положение всех неводородных атомов. Основная цепь выделена жирной линией, а боковые цепи - тонкими. Б. Пространственная модель, показывающая вандерваальсовы радиусы всех атомов (см. схему 3-1). В. Скелетная проволочная модель, составленная из отрезков, соединяющих все атомы а-углерода вдоль полипептидного скелета. Г. Ленточная модель, которая представляет все участки регулярных водородных связей либо в виде спиралей (а-спирали), либо в виде набора стрелок (ß-слои), указывающих на карбоксил- терминальный конец цепи; в этой модели также показаны водородные связи. Д. «Сосисочная» модель, которая демонстрирует расположение полипептидной цепи без всяких деталей. Следует иметь в виду, что сердцевина всех глобулярных белков плотно заполнена атомами, и впечатление пустого пространства вызвано только характером моделей В, Г и Д. (Б и В с любезного разрешения Richard J. Feldmann; А и Г - с любезного разрешения Jane Richardson.)
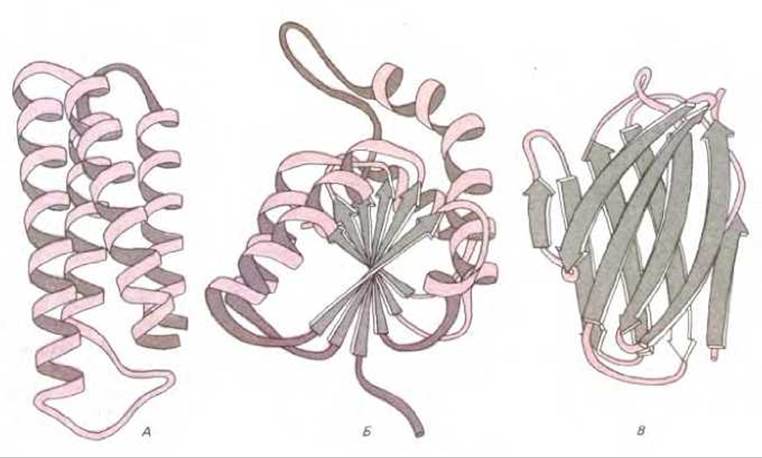
Рис. 3-32. Ленточные модели пространственной структуры некоторых белковых доменов с разной организацией. А. Цитохром b562, однодоменный белок, почти целиком состоящий из а-спиралей. Б. NAD-связанный домен лактат-дегидрогеназы, состоящий из смеси а-спиралей и ß-слоев. В. Изменчивый домен одной легкой цепи иммуноглобулина в виде сандвича из двух ß-слоев. На этих рисунках а-спирали и соединительные цепи окрашены, а цепи, составленные из ß-слоев, изображены серыми стрелками. Обратите внимание, что полипептидная цепь, как правило, пересекает домен два раза, делая резкие изгибы только на поверхности белковой молекулы. (Рисунок любезно предоставлен Jane Richardson.)
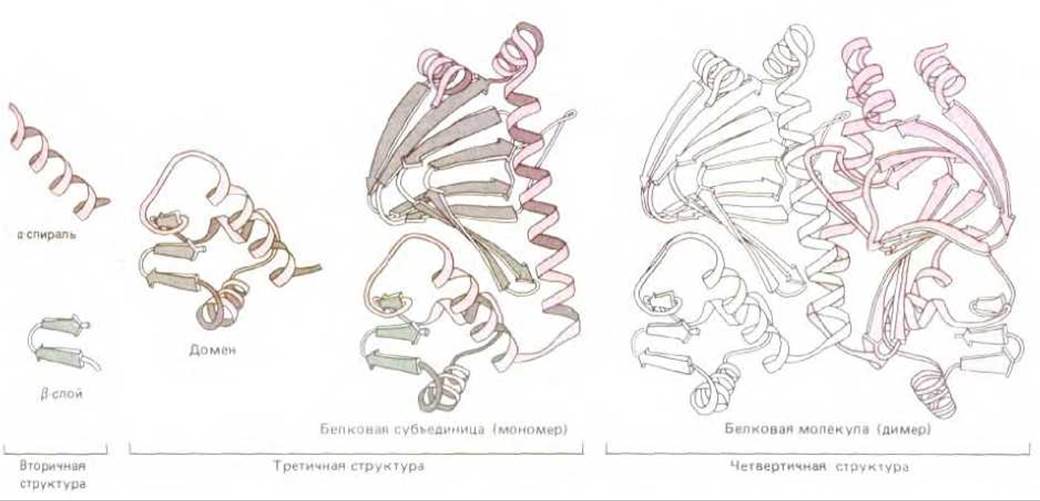
Рис. 3-33. Пространственная структура белка может быть описана в терминах различных уровней свертывания, каждый из которых составлен из структур предшествующего уровня в иерархическом порядке. Такие уровни иллюстрируются на этом рисунке на примере двухдоменного бактериального белка, активирующего катаболизм. Когда большой домен связывается с циклическим AMP, в белке происходит конформационное изменение, дающее возможность малому домену связываться со специфической последовательностью ДНК. Аминокислотная последовательность определяется как первичная структура белка, а первый уровень свертывания полипептидной цепи - как его вторичная структура. Как обозначено внизу рисунка под квадратными скобками, комбинацию второго и третьего уровней свертывания, представленную здесь, обычно называют третичной структурой, а четвертый уровень (комбинация субъединиц) - четвертичной структурой белка. (С изменениями с рисунков Jane Richardson.)

Рис. 3-34. Сопоставление аминокислотных последовательностей двух представителей семейства сериновых протеиназ. Показаны карбоксил- терминальные участки двух белков (от 149 до 245-й аминокислоты). Одинаковые аминокислоты соединены цветными штрихами, а сериновые остатки в активных центрах в положении 195 «высвечены». В участках полипептидных цепей, выделенных цветными прямоугольниками, каждая аминокислота этих двух ферментов в трехмерной структуре занимает одинаковое положение (см. рис. 3-35). Б. Стандартные однобуквенные и трехбуквенные обозначения аминокислот. (С изменениями из J. Greer, Proc. Natl, Acad. Sci. USA 77: 3393-3397, 1980.)
3.3.5. Сравнительно немногие потенциально возможные полипептидные цени могут оказаться полезными
Поскольку все 20 аминокислот химически различны и каждая может в принципе занимать в полипептидной цепи любое положение, то для пептида из четырех аминокислот возможны 20 ∙ 20 ∙ 20 ∙ 20 = 160000 различных цепей, а для полипептида из п аминокислот - 20n цепей. Таким образом, может существовать более 10390 различных белков со средней типичной длиной около 300 аминокислот.
Мы, однако, знаем, что лишь очень небольшая часть всех возможных белков примет стабильную пространственную конформацию. Все остальные должны иметь множество различных конформаций с разными химическими свойствами и приблизительно одинаковой энергией. Белки с такими изменчивыми свойствами не могут быть полезными и, следовательно, должны устраняться естественным отбором в ходе эволюции.
Удивительно точная пригнанность структуры современных белков к выполняемой ими функции обеспечивается их способностью свертываться уникальным образом. Последовательность аминокислот не только обеспечивает исключительную стабильность одной из конформаций, но и определяет необходимые для выполнения в клетке каталитической или структурной функции особенности этой конформаций и ее химические свойства. Белки строятся настолько точно, что замена даже нескольких атомов одной аминокислоты может нарушить структуру и привести к катастрофическим изменениям функции.
3.3.6. Новые белки часто возникают в результате незначительных изменений старых [25]
У клетки есть генетические механизмы, обеспечивающие дупликацию, модификацию и рекомбинацию генов в процессе эволюции (см, разд. 10.5.1). Следовательно, если уже какой-нибудь белок с полезными свойствами поверхности раз возникнет, то его основная структура может затем войти в состав многих других белков. В современных организмах различные белки с родственными функциями часто имеют схожую последовательность аминокислот. Считается, что такие семейства белков возникли путем дупликации одного предкового гена и последующего накопления в эволюции мутаций, постепенно обусловивших появление родственных белков с новыми функциями.
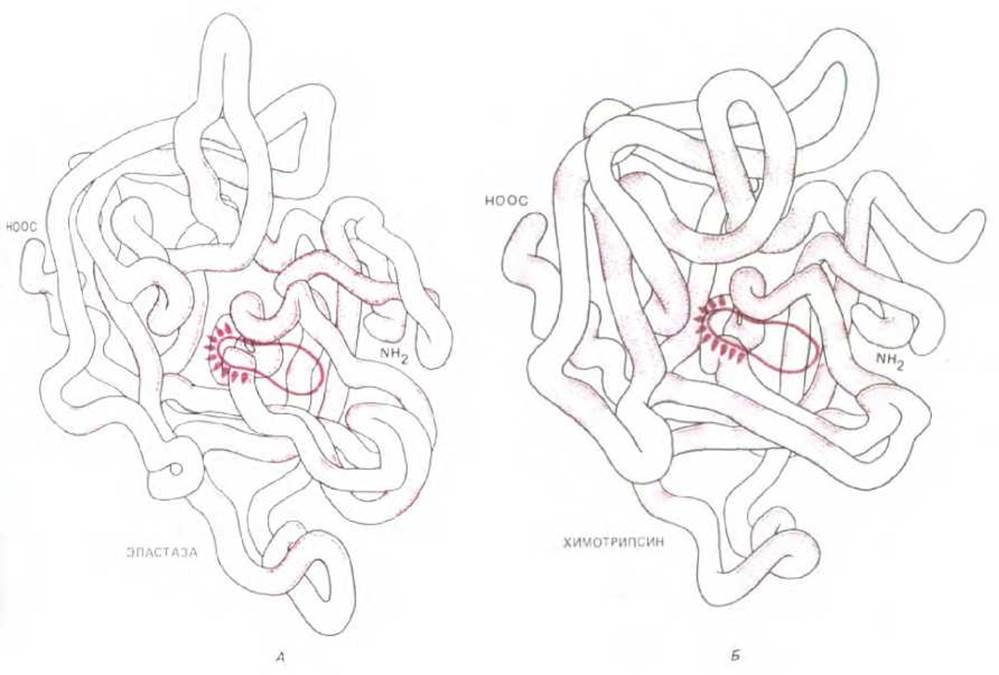
Рис. 3-35. Сравнение пространственной структуры эластазы (А) и химотрипсина (Б). У этих эволюционно родственных протеиназ одинаковы лишь те аминокислоты, которые расположены в выделенных цветом участках полипептидной цепи. Тем не менее конформации белков очень похожи. Обведены активные центры ферментов; оба активных центра содержат активированный остаток серина (см. рис. 3-47). Молекула химотрипсина имеет несколько (более двух) концов цепи, поскольку она образована протеолитическим расщеплением химотрипсиногена, неактивного предшественника.
Рассмотрим семейство протеолитических (расщепляющих) ферментов, сериновые протеиназы, включающие в себя пищеварительные ферменты химотрипсин, трипсин и эластазу, а также многие из факторов свертывания - протеиназ, контролирующих процесс свертывания крови. При сравнении любых двух ферментов этого семейства оказывается, что примерно 40% положений в полипептидной цепи занимают одни и те же аминокислоты (рис. 3-34). Еще более поразительное сходство выявляется при сравнении их конформаций, определенных методом рентгеноструктурного анализа: большинство поворотов и изгибов полипептидных цепей длиной в несколько сот аминокислот оказываются идентичными (рис. 3-35).
Тем не менее разные сериновые протеиназы имеют совершенно различные функции. Некоторые из аминокислотных замен, обусловивших различия ферментов этой группы, по-видимому, были отобраны в процессе эволюции, потому что привели к изменениям субстратной специфичности и регуляторных свойств белков, что в свою очередь породило все многообразие современных функциональных свойств. Другие аминокислотные замены могли быть «нейтральными», т. е. сохранились, потому что не повлияли ни на структуру, ни на функции белка. Поскольку мутирование - процесс случайный, должны были происходить и вредные замены, изменяющие пространственную структуру фермента достаточно сильно, чтобы его инактивировать. Эти измененные варианты были потеряны в процессе эволюции, так как производившие их индивидуальные организмы должны были оказаться в невыгодных условиях и исчезнуть в результате естественного отбора. Поэтому совершенно неудивительно, что клетки содержат целый набор структурно родственных полипептидных цепей, имеющих общих предков, но выполняющих разные функции.
3.3.7. Новые белки часто возникают в результате объединения разных полипептидных доменов [26]
При возникновении в клетке ряда стабильных белковых поверхностей новые поверхности с иной специфичностью связывания могут создаваться в результате объединения двух или более индивидуальных белков путем нековалентных взаимодействий. Для клеток характерно такое объединение глобулярных белков в более крупные функциональные белковые агрегаты: молекулярная масса многих белковых агрегатов достигает 1 млн. и более, хотя молекулярная масса типичной полипептидной цепи составляет всего лишь 40000-50000 (приблизительно 300-400 аминокислот); размер лишь немногих полипептидов втрое превышает эту среднюю величину.
Сходный, но другой способ образования новых белков из существующих полипептидных цепей - это слияние соответствующих последовательностей ДНК таким образом, что образуется ген, кодирующий одну большую полипептидную цепь (см. разд. 10.5.4). Считается, что белки, возникшие этим путем, в разных частях полипептидной цепи свертываются независимо в отдельные глобулярные домены. Такая «мультидоменная» структура характерна для многих белков, и, как и следовало ожидать, исходя из рассмотренных выше эволюционных предпосылок, функционально важные центры связывания часто оказываются расположенными на границе разных доменов (рис. 3-36). На рис. 3-37 показана структура конкретного мультидоменного белка.
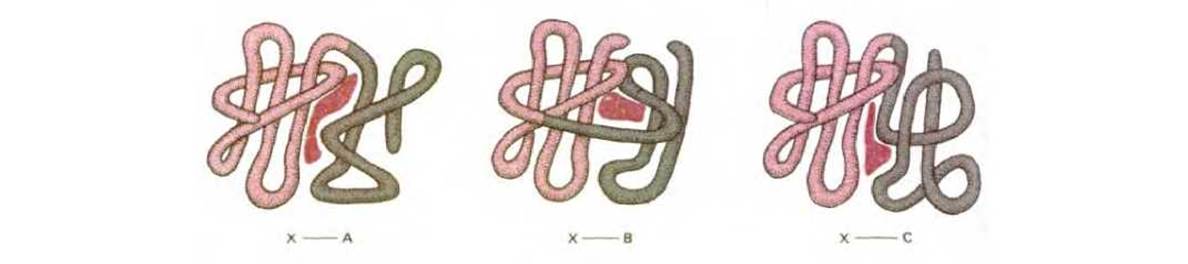
Рис. 3-36. Общий принцип, по которому наложение двух различных белковых поверхностей в процессе эволюции, приводит к появлению белков, которые содержат новые центры связывания для других молекул. Как показано на этом рисунке, лиганд - связывающие центры часто расположены в месте соприкосновения двух белковых доменов.
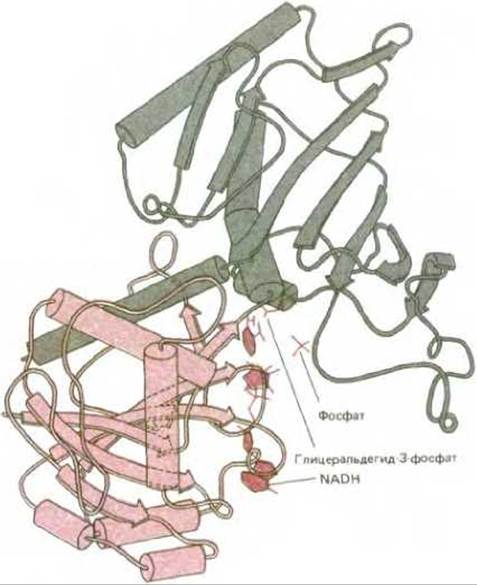
Рис. 3-37. Структура гликолитического фермента глицеральдегид-3-фосфат-дегидрогеназы. Белок состоит из двух доменов (выделены разным цветом). Участки а-спирали представлены в виде цилиндров, а ß-слоев - стрелками. Реакция, катализируемая этим ферментом, подробно приведена на рис. 2-21. Заметим, что три центра связывания субстратов расположены в зоне соприкосновения двух доменов. (С любезного разрешения Alan. J. Wonacott.)
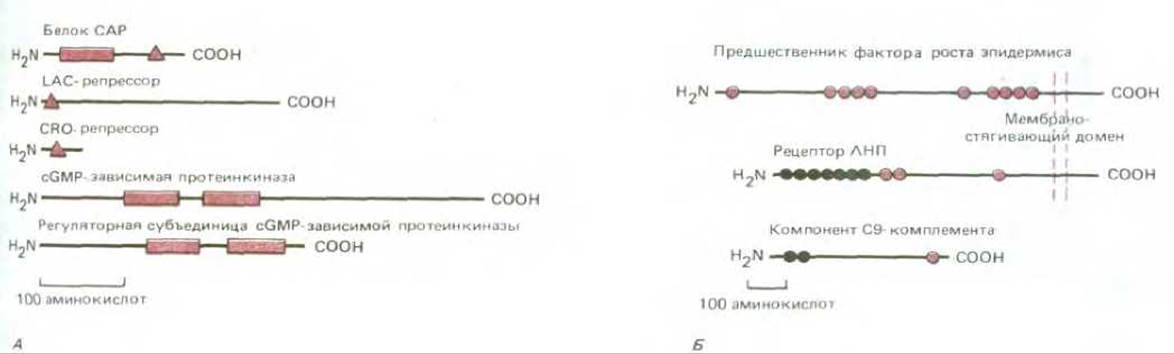
Рис. 3-38. Пример широко распространенной в эволюции белков «перетасовки» блоков белковых последовательностей. Участки белка, обозначенные окрашенными геометрическими фигурами, являются эволюционно родственными, но не идентичными. А. Бактериальный САР-белок состоит из двух доменов; один из них (закрашенный треугольник) связывается со специфической последовательностью ДНК, второй - связывает сАМР (см. рис. 3-33). ДНК-связывающий домен родствен ДНК-связывающим доменам многих других белков регуляторных генов, включая белки lac-репрессор и его-репрессор. Кроме того, две копии сАРМ-связывающего домена обнаружены в эукариотических киназах, регулируемых связыванием циклических нуклеотидов. Б. Представлены два домена, состоящие примерно из 40 аминокислот, каждый из которых встречается в трех больших белках позвоночных. Например, рецептор липопротеина низкой плотности (ЛНП) - это трансмембранный белок из 839 аминокислотных остатков, ответственный за выведение холестерола из клеток. Он содержит много доменов, имеющихся и в других белках, в частности, семь копий цистеин - богатого домена (светлые кружки), участвующих в связывании ЛНП, и три копии такого же размера (окрашенные кружки), функции которых неизвестны.
Другой путь повторного использования аминокислотной последовательности особенно распространен среди длинных фибриллярных белков, таких, как коллаген (см. рис. 3-28). В этом случае их структура формируется из многократных внутренних повторов предковой аминокислотной последовательности. Ясно, что сведение вместе аминокислотных последовательностей путем объединения ранее существовавших кодирующих последовательностей ДНК, является более эффективной стратегией для клетки, чем получать новые белковые последовательности в результате случайных мутаций ДНК.
3.3.8. Структурные гомологии могут помочь в определении функций вновь открытых белков [27]
Развитие методов быстрого секвенирования молекул ДНК сделало возможным определение аминокислотных последовательностей многих белков и нуклеотидных последовательностей соответствующих генов (см. разд. 4.6.6). Постоянно пополняемая «база данных белков» обрабатывается на компьютере для поиска возможных гомологии последовательностей между вновь секвенированным белком и изученными ранее. В настоящее время определена последовательность небольшого числа белков эукариотических организмов, при этом часто оказывается, что вновь секвенированный белок является гомологом уже известного белка в пределах какого-то участка его длины. Отсюда следует, что большинство белков, видимо, произошло от ограниченного числа предковых типов. Как и предполагалось, в последовательностях многих больших белков часто видны признаки того, что они возникли путем объединения ранее существовавших доменов в новых комбинациях, так называемого процесса «тасования доменов» (рис. 3-38).
Установление гомологии доменов также может быть полезным в другом аспекте. Определить пространственную структуру белка намного труднее, чем определить аминокислотную последовательность. Однако конфигурация домена вновь секвенированного белка может быть «отгадана», если он гомологичен домену белка, конформация которого ранее была определена методом рентгеноструктурного анализа. Часто можно с приемлемой точностью определить структуру нового белка, предполагая, что повороты и изгибы полипептидной цепи в двух белках будут одинаковыми, даже если есть отличия в аминокислотной последовательности.
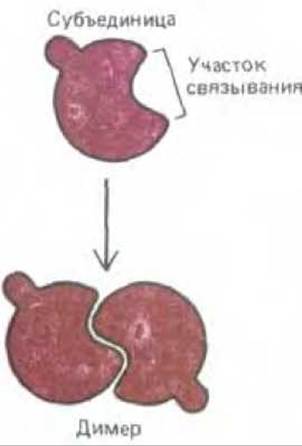
Рис. 3-39. Схема образования димера из идентичных белковых субъединиц. Если центр связывания узнает сам себя, димеры будут симметричными.
Эти пары часто в дальнейшем объединяются с другими субъединицами с образованием тетрамеров и более сложных ансамблей.
Такие сравнения белков важны еще и в том отношении, что сходные структуры часто предполагают и сходные функции. Можно избежать многолетних экспериментальных исследований, установив гомологию аминокислотной последовательности с белком, функция которого известна. Например, такие гомологии последовательностей впервые указали на то, что некоторые регуляторные гены клеточного цикла дрожжей и некоторые гены, вызывающие раковое перерождение клеток млекопитающих, кодируют протеинкиназы. Таким же способом было определено, что многие из белков, контролирующих морфогенез у плодовой мушки Drosophila, являются белками регуляторного гена, а один белок, участвующий в морфогенезе, был идентифицирован как сериновая протеиназа.
Каждый год эта база данных пополняется все новыми сведениями о белковых последовательностях, что увеличивает вероятность обнаружения полезных гомологии. Таким образом, сравнение аминокислотных последовательностей белков будет становиться все более важным инструментом клеточной биологии.
3.3.9. Белковые субъединицы способны к самосборке в большие клеточные структуры [28]
Принцип, позволяющий белковым доменам ассоциировать с образованием новых центров связывания, «работает» и при сборке значительно более крупных клеточных структур. Надмолекулярные структуры, такие, как ферментные комплексы, рибосомы, белковые волокна, вирусы и мембраны, не синтезируются в виде единых гигантских молекул, связанных ковалентными взаимодействиями, а собираются в результате нековалентной агрегации макромолекулярных субъединиц.
Использование субъединиц для построения больших структур имеет несколько преимуществ: 1) для построения большой структуры из многократно повторенных субъединиц меньшего размера требуется меньше генетической информации; 2) поскольку субъединицы связаны между собой многими сравнительно слабыми связями, их сборка и диссоциация легко поддаются контролю; 3) сборка структуры из субъединиц позволяет сводить к минимуму количество ошибок, так как функционирование специального механизма корректирования в процессе сборки может устранять испорченные субъединицы.
3.3.10. Одинаковые белковые субъединицы могут взаимодействовать с образованием геометрически регулярных структур [29]
При наличии в белке центра связывания, комплементарного какому-либо участку на его собственной поверхности, белок будет самопроизвольно агрегировать. В простейшем случае центр связывания узнает сам себя, и в результате образуется симметричный димер. Многие ферменты и другие белки образуют такие димеры, которые часто в свою очередь служат субъединицами для формирования более крупных агрегатов (рис. 3-39 и рис. 3-40).
Если центр связывания белка комплементарен другому участку на своей поверхности, то образуется цепь субъединиц. При некоторых взаимных ориентациях двух участков связывания цепь замкнется сама на себя и рост прекратится. В результате образуется кольцо из двух, трех, четырех или большего числа субъединиц (рис. 3-41). В более общем случае получится бесконечно длинный полимер из белковых субъединиц. При условии, что все субъединицы связаны друг с другом идентичным образом, субъединицы в такой цепи расположатся по спирали (см. рис. 3-3). Например, актиновая нить представляет собой спиральную структуру, собранную из одинаковых субъединиц глобулярного белка актина; актиновые нити являются основными компонентами цитозоля большинства эукариотических клеток. Когда особенно важна механическая прочность, надмолекулярные агрегаты обычно строятся не из глобулярных, а из фибриллярных субъединиц, поскольку фибриллярные субъединицы, обвиваясь вокруг друг друга в спираль, имеют обширные области для белокбелкового взаимодействия (рис. 3-42, А).
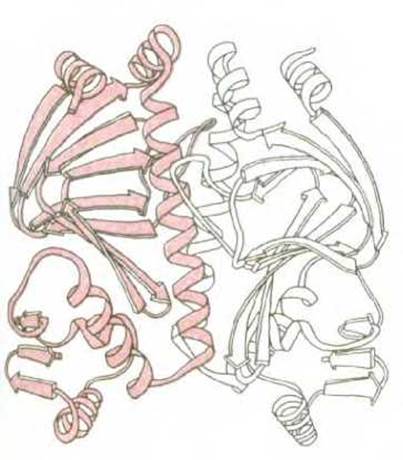
Рис. 3-40. Ленточная модель димера, образованного из двух идентичных белковых субъединиц (мономеров). Представленный белок является бактериальным белком САР, показанным ранее на рис. 3-33 и рис. 3-38, А. (С любезного разрешения Jane Richardson.)
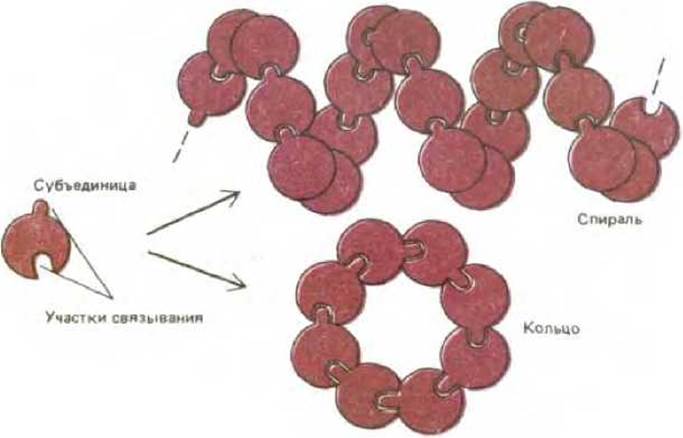
Рис. 3-41. Одинаковые субъединицы при взаимодействии друг с другом могут формировать кольца или спирали. Образование спирали было показано на рис. 3-3, образование кольца вместо спирали происходит, если субъединицы входят друг в друга, останавливая дальнейший рост цепи.
Гексагонально упакованные белковые субъединицы могут образовывать плоские слои. Иногда так агрегируют в липидных бислоях специализированные мембранные транспортные белки (см. разд. 6.2.8). При небольшом изменении геометрии субъединиц гексагональный слой превращается в полую трубку (рис. 3-42, Б). Такие цилиндрические трубки участвуют в образовании белковых оболочек некоторых удлиненных вирусов (рис. 3-43).
Образование замкнутых структур - колец, трубок или сферических частиц - дополнительно стабилизирует весь арегат; общее число связей между белковыми субъединицами в этом случае увеличивается. Более того, поскольку такая структура формируется благодаря взаимозависимым кооперативным взаимодействиям, то сборка и разборка могут производиться относительно малыми изменениями, затрагивающими сами субъединицы. Особенно ярко это можно проиллюстрировать на примере белковых оболочек многих простых вирусов, имеющих форму полого шара. Такие оболочки часто собраны из сотен идентичных белковых субъединиц, окружающих и защищающих вирусную нуклеиновую кислоту (рис. 3-43). Структура белков оболочки должна быть особенно гибкой, так как она должна допускать различные типы межсубъединичных контактов, а также обеспечивать изменение упаковки субъединиц при выходе нуклеиновой кислоты в начале цикла размножения вируса.
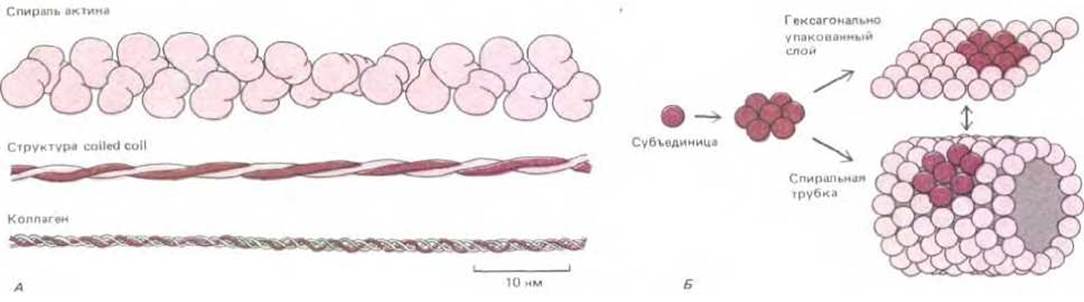
Рис. 3-42. Некоторые структуры, образующиеся при самосборке белковых субъединиц. А. Три общих типа спиральных ансамблей белка. В актиновой нити содержится примерно две глобулярные белковые субъединицы на один оборот, а многие другие цитоскелетные белки содержат стержневидные участки, в которых две а-спирали объединяются в структуру "coiled coil". В спирали коллагена три вытянутые белковые цепи объединяются друг с другом на большом расстоянии с образованием очень прочной стержнеобразной структуры. Б. Гексагонально упакованные глобулярные белковые субъединицы могут формировать либо плоские структуры, либо трубки.
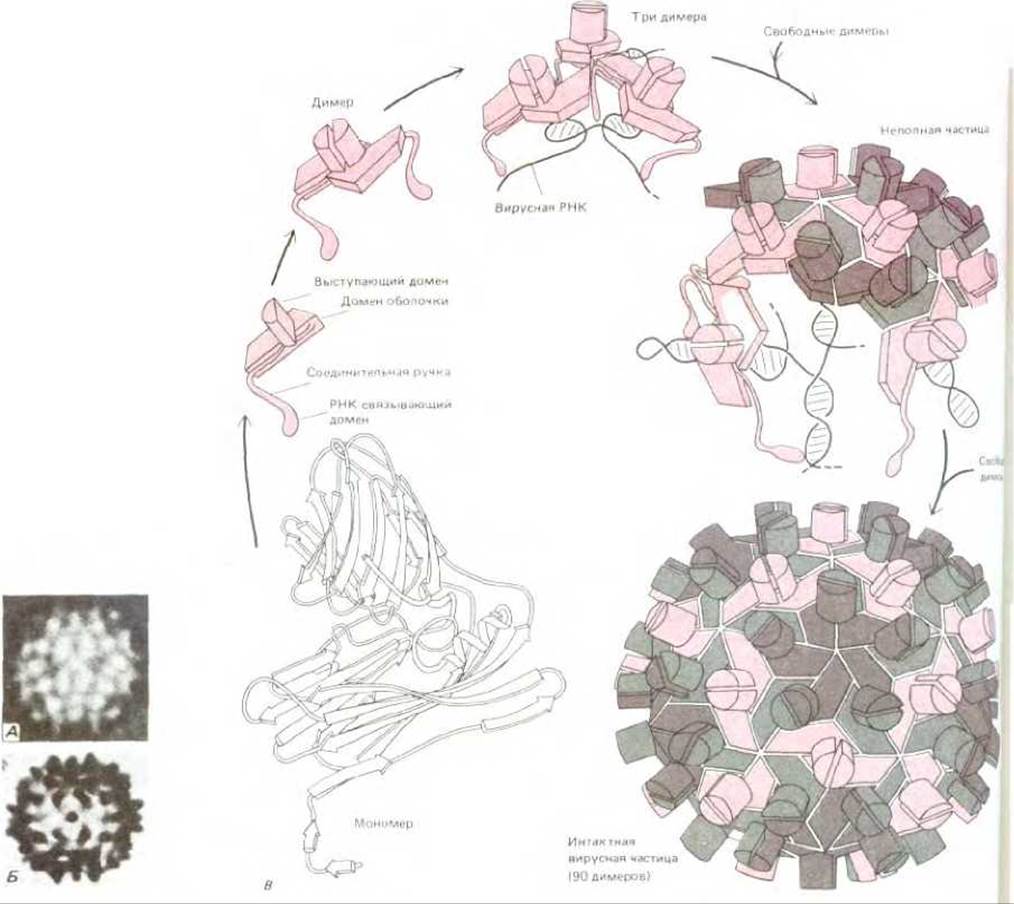
Рис. 3-43. Структура сферического вируса. Во многих вирусах идентичные белковые субъединицы упаковываются с образованием сферической оболочки, которая заключает вирусный геном, состоящий из РНК или ДНК. По геометрическим соображениям симметричным образом могут упаковаться не более 60 субъединиц. Однако если допустимы небольшие отклонения от регулярности, то можно использовать больше субъединиц для образования более крупного капсида. Например, вирус кустистой карликовости томата (TBSV) имеет форму сферы около 33 нм в диаметре. На электронной микрофотографии (А) и на схеме (Б) можно видеть, что он состоит из более, чем 60 субъединиц. Предполагаемый способ сборки и трехмерная структура по данным рентгеноструктурного анализа этого вируса представлены на В. Вирусная частица состоит из 180 идентичных копий капсидного белка (насчитывающих по 386 аминокислот) и генома РНК, включающего 4500 нуклеотидов. Чтобы сформировать такой крупный капсид, белок должен быть способен упаковываться тремя несколько различными способами (обозначены разным цветом). (Рисунки выполнены Steve Harisson; электронные микрофотографии - с любезного разрешения John Finch.)

Рис. 3-44. Электронная микрофотография вируса табачной мозаики (ВТМ). Вирус состоит из одной длинной молекулы РНК, окруженной плотно уложенной спиралью из идентичных белковых субъединиц, образующих цилиндрическую оболочку. Очищенная РНК и белок оболочки при смешивании в пробирке самопроизвольно образуют полностью инфекционные вирусные частицы. (С любезного разрешения Robley Williams.)
3.3.11. Самособирающиеся структуры могут состоять из различных белковых субъединиц и нуклеиновых кислот [30]
Многие белковые клеточные структуры, такие, как вирусы и рибосомы, построены из белковых субъединиц и молекул РНК или ДНК. Информация о сборке таких сложных агрегатов заключена в строении самих макромолекулярных субъединиц и в соответствующих условиях изолированные субъединицы могут самопроизвольно собираться в пробирке в конечную структуру. Впервые возможность самосборки большого макромолекулярного агрегата из отдельных компонентов была обнаружена у вируса табачной мозаики (ВТМ). Этот вирус представляет собой длинный стержень, в котором белковый цилиндр окружает спиральную сердцевину из РНК (рис. 3-44 и рис. 3-45). Если очищенную вирусную РНК и белковые субъединицы смешать в растворе, они агрегируют с образованием полностью активных вирусных частиц. Процесс самосборки оказался неожиданно сложным: он сопряжен с образованием особых промежуточных структур - двойных белковых колец, присоединяющихся к растущей ободочке вируса.
Другой пример макромолекулярного арегата, структура которого после диссоциации на отдельные компоненты восстанавливается, - это рибосома бактерий. Бактериальные рибосомы состоят приблизительно из 55 различных белковых молекул и трех различных молекул РНК (см. разд. 5.1.8). Если инкубировать в пробирке в соответствующих условиях все индивидуальные компоненты, то они самопроизвольно соберутся в рибосому. Важнее всего то, что такие реконструированные рибосомы способны осуществлять биосинтез белков. Реконструкция рибосом, как и предполагалось, происходит упорядоченно: сначала к РНК присоединяются определенные белки, затем другие белки узнают образовавшийся комплекс и т.д., пока не завершится формирование полной структуры.
До сих пор неясно, каким образом осуществляется регуляция некоторых более сложных процессов самосборки. Оказалось, например, что многие клеточные структуры имеют точно определенную длину, во много раз превышающую длину всех составляющих их макромолекул. Как достигается столь точное ограничение длины, остается загадкой. На рис. 3-46 представлены три возможных механизма такого ограничения. В простейшем случае длинный каркас белка или другой макромолекулы является ограничителем, который определяет размер конечной структуры. Именно такой механизм определяет длину частицы ВТМ, где молекула РНК служит таким стержнем. Аналогично было показано, что белковый каркас определяет длину хвостов некоторых бактериальных вирусов (рис. 3-47).

Рис. 3-45. Модель элемента структуры вируса табачной мозаики. Одноцепочечная молекула РНК из 6000 нуклеотидов упакована в белковую оболочку, состоящую из 2130 копий специального белка (каждая его молекула состоит из 158 аминокислотных остатков).

Рис. 3-46. Три возможных способа, с помощью которых большие белковые ансамбли могут поддерживать фиксированную длину: А. Объединение вдоль вытянутого каркаса из белка или другой макромолекулы, который служит в качестве «измерительного устройства»; Б. Добавление к полимерной структуре дополнительных субъединиц сверх определенной длины требует слишком много энергии и объединение субъединиц прекращается. В. Сборка по типу нониуса. Два набора стержневидных молекул отличаются по длине от собранного комплекса и его рост прекратится, когда концы таких молекул в точности совпадут.

Рис. 3-47. Электронная микрофотография бактериофага X. Конец хвостового отростка фаговой частицы прикрепляется к специфическому белку на поверхности бактериальной клетки, после чего ДНК, плотно упакованная в головке вируса, инъецируется через хвост в клетку. Хвост имеет точную длину, которая определяется при помощи механизма, показанного на рис. 3-46, А.
3.3.12. Не все клеточные структуры образуются путем самосборки [31]
Некоторые клеточные структуры, удерживаемые вместе нековалентными связями, не способны к самосборке. Например, митохондрии, реснички или миофибриллы не могут самопроизвольно собираться в растворе из макромолекулярных компонентов, поскольку часть информации для их сборки заложена в специальных ферментах и других клеточных белках, выполняющих функции шаблонов и матриц, но не входящих в состав окончательной структуры. Порой даже маленькие структуры лишены некоторых необходимых для сборки компонентов. Например, при формировании некоторых бактериальных вирусов головка, построенная из одинаковых белковых субъединиц, собирается на временном каркасе, построенном из другого белка. Этого второго белка нет в окончательной вирусной частице, и, следовательно, головка не может самопроизвольно собраться в его отсутствие. Известны другие примеры, когда существенной и необратимой стадией процесса сборки является протеолитическое расщепление. Именно так формируются оболочки некоторых бактериальных вирусов и даже некоторые простые белки, в том числе структурный белок коллаген и гормон инсулин (рис. 348). На основании этих сравнительно простых примеров можно прийти к выводу, что сборка таких сложных структур, как митохондрия или ресничка, управляется и во времени, и в пространстве другими клеточными компонентами и, кроме того, включает в себя стадии необратимого созревания, катализируемые расщепляющими ферментами.
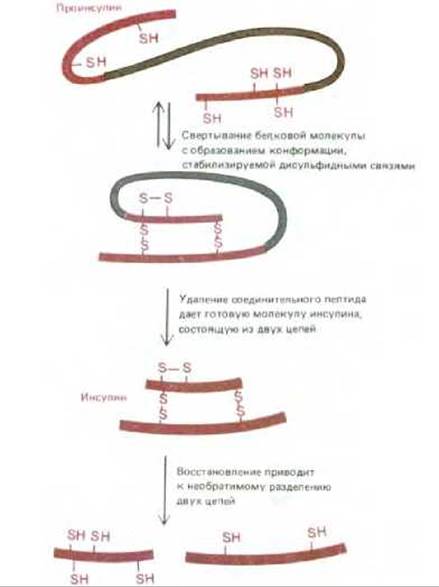
Рис. 3-48. Полипептидный гормон инсулин синтезируется в виде белка-предшественника проинсулина, который свертывается нужным образом, а затем расщепляется протеолитическим ферментом. Поэтому после восстановления дисульфидных связей инсулин не может самопроизвольно принять исходную конформацию. Вырезание части полипептидной цепи проинсулина приводит, таким образом, к потере информации, необходимой для самосборки молекулы.
Заключение
Аминокислотная последовательность белковой молекулы определяет ее пространственную структуру. Конкретная структура полипептидной цепи стабилизируется нековалентными взаимодействиями между ее частями. Аминокислоты с гидрофобными группами стремятся сгруппироваться внутри молекулы, а возникновение локальных водородных связей между соседними пептидными группами приводит к образованию а-спиралей и ß-слоев. Многие белки собраны, как из модулей, из небольших глобулярных образований, называемых доменами; малые белки обычно состоят из одного домена, тогда как большие содержат несколько доменов, скрепленных вместе короткими участками полипептидной цепи. При построении новых белков домены изменяются и комбинируются с другими доменами.
Те же силы, которые определяют пространственную структуру белков, ответственны и за образование белковых агрегатов. Белки, имеющие центр связывания, комплементарный их собственной поверхности, могут образовывать димеры, замкнутые кольца, сферические частицы или спиральные полимеры. Смесь множества различных белков, содержащая иногда структурные нуклеиновые кислоты, может самопроизвольно собираться в пробирке в большие сложные структуры. Однако не все клеточные структуры способны к самопроизвольной реконструкции после диссоциации на отдельные компоненты, так как процесс сборки во многих случаях включает необратимые этапы.