Молекулярная биология клетки - Том 1 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
Введение в биологию клетки
Эволюция клетки
От клеток - к многоклеточным организмам
Одноклеточные организмы, вроде бактерий и простейших, столь успешно адаптировались к разнообразным условиям среды, что составляют более половины всей биомассы Земли. В отличие от высших животных многие из этих одноклеточных способны синтезировать все необходимые им вещества из нескольких простых соединений, причем некоторые из них делятся чаще, чем раз в час. Чем же тогда объяснить появление и распространение на Земле многоклеточных организмов? По-видимому, их преимущество состоит в том, что такие организмы могут использовать ресурсы, недоступные единичной клетке. Например, наличие множества клеток позволяет дереву достичь больших размеров, иметь корни в земле (где определенные клетки поглощают воду и питательные вещества) и листья в воздухе (где другие клетки могут эффективно улавливать энергию солнечных лучей). Ствол дерева состоит из специализированных клеток, образующих каналы для транспорта воды и питательных веществ между листьями и корнями. Другая группа специализированных клеток образует слой коры, предотвращающий потерю воды и защищающий внутреннюю часть ствола. Дерево как целое не конкурирует с одноклеточными организмами за свою экологическую нишу, его способ выживания и размножения совершенно иной. По мере появления различных растений и животных изменялась и среда, в которой проходила эволюция, а это в свою очередь требовало новых способностей, ведь существование в джунглях, например, предполагает наличие у организмов совершенно иных свойств, чем жизнь в открытом море. Возникновение новых способов движения, сенсорного восприятия, коммуникаций, социальной организации - все это позволило эукариотическим организмам конкурировать, размножаться и заселять пространства Земли.
1.3.1. Одиночные клетки способны объединяться и образовывать колонии
Вполне вероятно, что ранней стадией эволюции многоклеточных явилось объединение одноклеточных организмов в колонии. Достигнуть этого проще всего, если дочерние клетки не расходятся после каждого клеточного деления. В примитивном виде такое общественное поведение встречается даже у прокариот. Например, миксобактерии, которые живут в почве и питаются нерастворимыми органическими молекулами, расщепляют их, секретируя специальные ферменты. Они образуют рыхлые колонии, в которых накапливаются ферменты от индивидуальных клеток и, таким образом, повышается эффективность питания (эффект «волчьей стаи»). Эти клетки представляют собой вершину структурной сложности, достигнутой эволюцией прокариот: когда истощаются запасы питательных веществ, они плотно агрегируют и образуют плодовое тело (рис. 1-31), в котором бактерии дифференцируются в споры, способные выжить даже в исключительно суровых условиях.
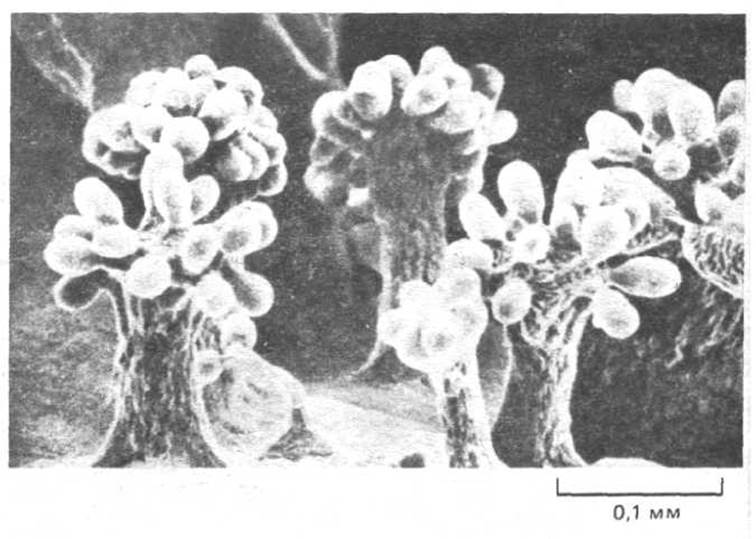
Рис. 1-31. Микрофотография (сканирующий электронный микроскоп) плодовых тел, образованных миксобактериями (Chondromyces crocatus). Каждое плодовое тело, заполненное спорами, образовалось в результате агрегации и дифференцировки примерно миллиона миксобактерии. (Из P. L. Grilione, J. Pangbom, J. Bacteriol., 124, 1558-1565, 1975.)
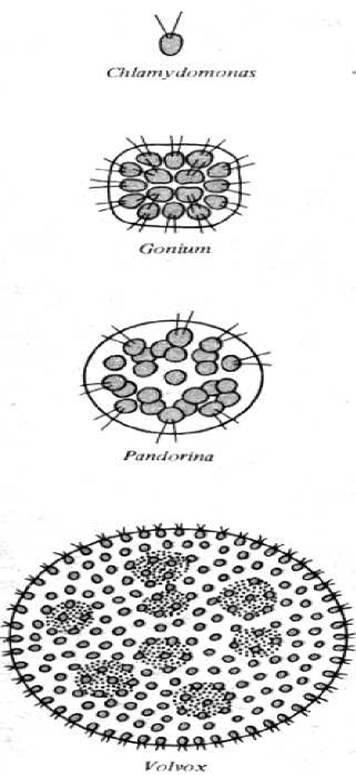
Рис. 1-32. Четыре близких рода зеленых водорослей, иллюстрирующих развитие от одноклеточной к колониальной и далее к многоклеточной организации.
Когда условия становятся более благоприятными, споры плодового тела прорастают и образуют новое скопление бактерий.
Зеленые водоросли (не путать с прокариотическими синезелеными водорослями, или цианобактериями) - это эукариоты, существующие в виде одноклеточных организмов, колоний и многоклеточных форм. Различные виды зеленых водорослей можно расположить в порядке возрастания сложности, иллюстрирующем последовательность эволюции, которую, видимо, прошли высшие растения и животные. Одноклеточные зеленые водоросли, такие как Chlamydomonas, напоминают жгутиковых простейших, но в отличие от них имеют хлоропласты и способны к фотосинтезу. У близких родов группы жгутиковых клеток образуют колонии, удерживаемые вместе матриксом из секретируемых самими же клетками молекул. Наиболее просто организованные виды (принадлежащие к роду Gonium) имеют форму вогнутого диска, состоящего из 4, 8, 16 или 32 клеток. Биения их жгутиков независимы друг от друга, но ориентированы в одном направлении, поэтому они способны приводить колонию в движение. Каждая клетка такой колонии равноценна любой другой, и деление каждой клетки может дать начало новой колонии. В других родах можно обнаружить большие по размерам колонии. Наиболее примечательный пример - род Volvox; некоторые его виды содержат до 50 000 и более клеток, образующих полый шарик. У представителей рода Volvox индивидуальные клетки колонии соединены тонкими цитоплазматическими мостиками, так что биение жгутиков координированы и вся колония движется подобно катящемуся шарику (рис. 1-32). В колонии Volvox наблюдается некоторое разделение функций - за воспроизведение отвечает небольшое количество клеток, служащих предшественниками новых колоний. Остальные клетки столь зависимы друг от друга, что неспособны к самостоятельному существованию, и разрушенная колония погибает.
1.3.2. Клетки высших организмов становятся специализированными и взаимозависимыми
В определенном смысле Volvox больше напоминает многоклеточный организм, чем просто колонию. При движении в воде биения жгутиков синхронизированы, колония структурно и функционально асимметрична. Она способна двигаться по направлению к отдаленному источнику света. Репродуктивные клетки обычно локализуются с одного края колонии, там они делятся и образуют новые миниатюрные колонии, укрытые вначале внутри родительского шарика. Таким образом, у Volvox в примитивной форме проявляются две существенные черты всех многоклеточных организмов: клетки специализируются и кооперируются. С помощью специализации и кооперации клетки образуют единый координированный организм, обладающий более широкими возможностями, чем любая из составляющих его частей.
Организованные формы дифференцировки клеток встречаются даже у некоторых прокариот, например, многие цианобактерии не расходятся после деления, образуя нитевидные цепочки до метра длиной. Через регулярные интервалы в такой цепочке встречаются изменившиеся клетки, способные включать атмосферный азот в органические молекулы. Эти немногие специализированные клетки осуществляют фиксацию азота не только для себя, но и для соседних клеток, с которыми они обмениваются продуктами метаболизма. Однако эукариотические клетки добились значительно больших успехов в таком «разделении труда». И именно они, а не прокариоты являются теми живыми единицами, из которых построены все наиболее сложные многоклеточные организмы.
1.3.3. В основе многоклеточной организации лежит взаимодействие клеток
Чтобы образовать многоклеточный организм, клетки должны быть как-то связаны друг с другом. Эукариоты развили целый ряд приспособлений для осуществления этой функции. У Volvox, как уже отмечалось, клетки не полностью расходятся при делении, а остаются соединенными цитоплазматическими мостиками. У высших растений клетки не только связаны цитоплазматическими мостиками - плазмодесмами, но и заключены в жесткие «соты» со стенками из целлюлозы (клеточные стенки), которую сами клетки и секретируют.
Клетки большинства животных не имеют жестких стенок, а цитоплазматические мостики у них редки. Вместо этого клетки объединены сравнительно рыхлой сетью больших внеклеточных органических молекул (называемых внеклеточным матриксом), а также за счет слипания (адгезии) их плазматических мембран. Например, организм губок (их обычно считают наиболее примитивными из современных животных), как правило, состоит из пяти типов специализированных клеток, образующих оболочку тела с системой пор и каналов для прокачивания воды, из которой клетки отфильтровывают и поглощают частички пищи. Благодаря делению клеток, губки неограниченно растут: их размер и структура точно не предопределены. Они лишены нервной системы, которая могла бы координировать активность различных частей организма. Их можно описать как «свободную республику клеток» в отличие от более строго организованных клеточных сообществ, характерных для высших животных. Если продавить губку через тонкое сито, чтобы механически разделить отдельные клетки, эти клетки могут иногда самопроизвольно собраться в целую губку: сначала клетки агрегируют в большую неупорядоченную массу, а затем перегруппировываются в организованный многоклеточный слой. Такие слои клеток называют эпителием.
1.3.4. Эпителиальные слои клеток окружают защищенную от внешних воздействий внутреннюю среду организма
Из всех способов взаимосвязи клеток в тканях многоклеточных животных наиболее фундаментальное значение, видимо, имеет эпителиальная организация. В эволюции сложных многоклеточных организмов эпителиальный слой сыграл столь же большую роль, как и клеточная мембрана в эволюции сложных одиночных клеток.
Значение эпителиальных слоев легко проиллюстрировать на примере другой группы низших организмов - кишечнополостных, которые на эволюционной лестнице стоят на ступеньку выше губок, так как имеют что-то вроде нервной системы. Эта группа животных включает в себя медуз, актиний, коралловые полипы, а также гидру, маленький пресноводный организм. Тело кишечнополостных состоит из двух слоев эпителия: наружного - эктодермы и внутреннего - энтодермы. Энтодермальный слой окружает гастроваскулярную полость, в которой происходит переваривание пищи (рис. 1-33). Некоторые энтодермальные клетки секретируют пищеварительные ферменты в гастроваскулярную полость, другие осуществляют всасывание и дальнейшее переваривание пищевых молекул, образовавшихся под действием этих ферментов. Образуя плотный эпителиальный слой, клетки энтодермы препятствуют выходу всех этих молекул в окружающую среду. В результате в гастроваскулярной полости создаются условия, необходимые для нормального пищеварения. В то же время обращенные наружу клетки эктодермы сохранили специализацию, полезную при взаимоотношениях с внешним миром.
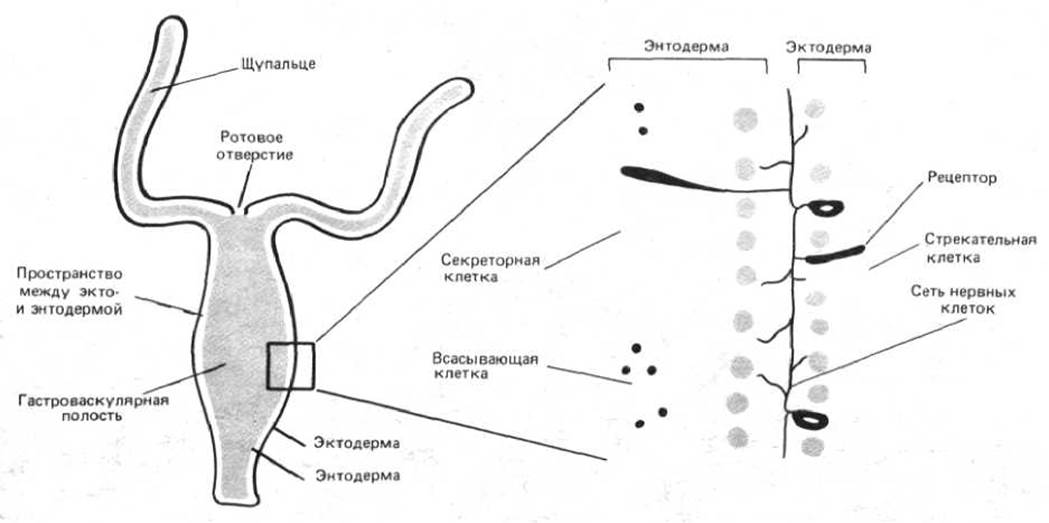
Рис. 1-33. Строение тела гидры. Наружный слой клеток (эктодерма) выполняет защитные, чувствительные функции и функции захвата добычи, а внутренний слой (энтодерма) в основном участвует в пищеварении. Оба клеточных пласта способны к сокращению (т. е. выступают в роли мышц), что дает возможность животному двигаться. Между этими слоями расположена сеть взаимосвязанных нервных клеток, которые координируют движение.
В эктодерме, например, есть специальные клетки (стрекательные клетки), содержащие капсулу, заполненную ядовитой жидкостью. Плотная стенка этой капсулы в дистальной части образует спирально закрученную стрекательную нить, способную выбрасываться и парализовать тех мелких животных, которыми гидра питается. Большинство других экто- и энтодермальных клеток обладают мышечноподобными свойствами, что дает возможность гидре двигаться, как и положено хищнику.
Между эктодермой и энтодермой находится еще один компартмент, отделенный как от гастроваскулярной полости, так и от внешней среды. Здесь в узком замкнутом пространстве между эпителиальными клетками расположены нервные клетки. Под внешней поверхностью формируются специализированные клеточные контакты, образующие непроницаемый барьер. Сокращая мышечноподобные клетки эктодермы и энтодермы, животное способно изменять форму и двигаться. Контроль и координацию этих сокращений осуществляют нервные клетки, которые способны проводить электрические сигналы (рис. 1-33, 1-34 и 1-35). Как мы увидим в дальнейшем, для нормального функционирования нервных клеток критическое значение имеет концентрация простых неорганических ионов в окружающей среде. Большинство нервных клеток, в том числе и наши собственные, приспособлены для работы в растворе, ионный состав которого аналогичен составу морской воды, что, видимо, отражает условия возникновения первой нервной клетки. Большинство кишечнополостных по-прежнему обитает в море, но не все. В частности, гидра живет в пресной воде. Очевидно, что заселение этой новой среды обитания оказалось возможным лишь в силу того, что нервные клетки гидры находятся в замкнутом и изолированном от внешней среды пространстве, образованном слоями эпителиальных клеток, которые поддерживают внутреннюю среду, необходимую для функционирования нервных клеток.
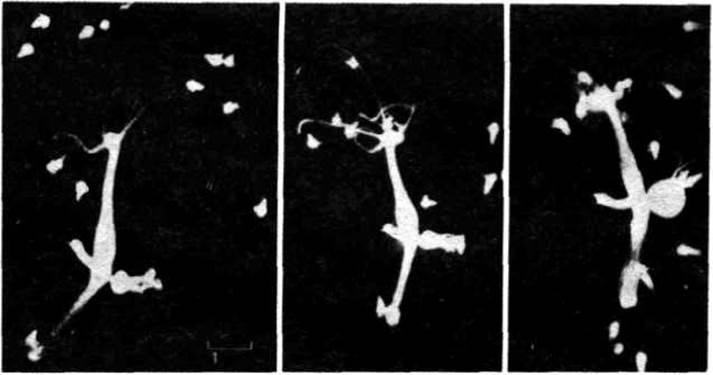
Рис. 1-34. Гидра может проявлять достаточно сложную активность. На фотографии видно, как она захватывает своими щупальцами дафнию и засовывает свою добычу в гастроваскулярную полость для переваривания. (С любезного разрешения Amata Horubruch.)
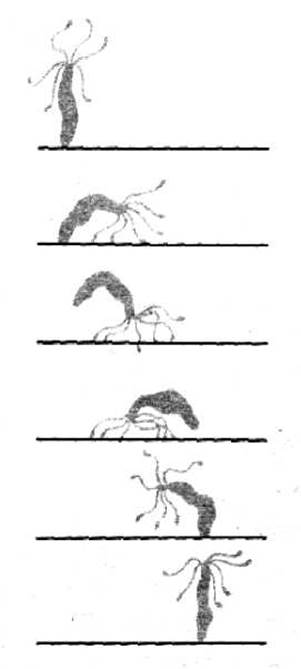
Рис. 1-35. Гидра может плавать, скользить на своей подошве или, как показано здесь, перемещаться кувыркаясь.
1.3.5. Межклеточные коммуникации определяют пространственное строение многоклеточных организмов
Клетки гидры связаны между собой не только механически, но и с помощью специальных контактов, изолирующих внутреннюю часть организма от внешней среды. Известно, что клетки этого организма способны передавать сигналы по всей длине тела. Если отрезать один конец гидры, оставшиеся клетки среагируют на отсутствие ампутированной части изменением своих свойств и перестроятся так, чтобы регенерировать целое животное. Очевидно, что от одной части организма к другой передаются сигналы, управляющие развитием формы тела, со щупальцами и ртом на одном конце и стебельком на другом. Более того, эти сигналы независимы от нервной системы. Если развивающуюся гидру обработать ядом, препятствующим образованию нервных клеток, то животное потеряет способность двигаться, ловить добычу и питаться. Однако его пищеварительные системы будут по-прежнему работать нормально, так что любой человек, достаточно терпеливый, чтобы вкладывать обычную добычу гидры ей в рот, сможет поддерживать ее существование. Форма тела у таких принудительно питаемых животных нормальная, и утерянные части регенерируют точно так же, как у животных с интактной нервной системой.
В ходе эволюции из просто устроенных организмов, похожих на кишечнополостных, развились высшие животные. Сложная организация последних обусловлена более изощренным использованием тех же принципов кооперирования клеток, которые лежат в основе строения гидры. Слои эпителиальных клеток выстилают все внутренние и наружные поверхности тела, создавая защищенные от внешних условий компартменты и контролируемую внутреннюю среду, в которой дифференцированные клетки выполняют специализированные функции. Специализированные клетки взаимодействуют и сообщаются друг с другом с помощью сигналов, управляющих свойствами каждой клетки в зависимости от ее места в общей структуре. Однако, чтобы понять, как устроен такой большой многоклеточный организм со сложной и точной организацией, каким является человек, необходимо более подробно рассмотреть последовательность событий при его развитии.
1.3.6. Клеточная память позволяет развиваться сложным формам
Клетки почти всех многоклеточных организмов возникают в результате последовательных делений одной-единственной клетки- предшественника, следовательно, они представляют собой клон. По мере деления и роста клеток происходит их дифференцировка, т.е. они начинают отличаться от других по своей структуре, метаболизму и функциям. Изменения эти происходят в ответ на сигналы, поступающие от соседних клеток. Примечательно, что эукариотические клетки и их потомство обычно сохраняют свое специализированное состояние даже после того, как влияние, вызвавшее дифференцировку, исчезает, другими словами, у этих клеток есть память. А это значит, что их конечные свойства определяются не только тем окружением, в котором они в конце концов оказались, а всей совокупностью влияний, которым они подвергались в ходе развития организма. Таким образом, по мере роста и развития постепенно проявляются все более и более тонкие детали плана строения взрослого организма. Так создается организм постепенно увеличивающейся сложности, окончательная форма которого является выражением длительной истории индивидуального развития.
1.3.7. Основные программы развития имеют тенденцию сохраняться в процессе эволюции
Строение животного является результатом его эволюционной истории, которая, как и история индивидуального развития, представляет собой летопись продвижения от простого к сложному. Каково же взаимоотношение этих двух аспектов развития - эволюционного и индивидуального? В процессе эволюции многие приспособления, использовавшиеся при индивидуальном развитии простейших многоклеточных организмов, сохранились в качестве основных принципов строения их более сложных потомков. Мы уже упоминали об эпителиальной организации клеток. Заслуживает внимания и тот факт, что некоторые специализированные типы клеток, например нервные клетки, можно найти практически у любого животного - от гидры до человека. Более того, ранние стадии индивидуального развития животных, взрослые формы которых совершенно различны, часто бывают удивительно похожи. Так, например, лишь специалист способен отличить куриный эмбрион от эмбриона человека на ранней стадии развития (рис. 1-36).
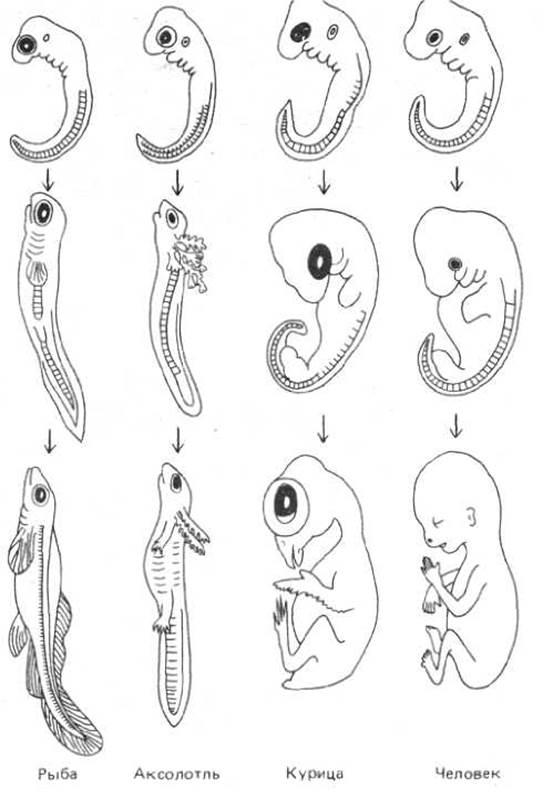
Рис. 1-36. Сравнение эмбрионального развития рыбы, амфибии, птицы и млекопитающего. Ранние стадии (вверху) очень похожи, более поздние стадии (внизу) различаются сильнее. (Из Е. Haeckel, Anthroppogenie, oder Entwickelungsgeschichte des Menschen. Leipzig, Engelmann, 1874.)
Подобные наблюдения нетрудно объяснить. Рассмотрим процесс возникновения в ходе эволюции новой анатомической особенности, скажем удлиненного клюва. Случайная мутация изменяет аминокислотную последовательность белка и, следовательно, его биологическую активность. Измененный белок может повлиять на клетки, ответственные за образование клюва таким образом, что в результате получится более длинный клюв. Но мутация должна быть совместима с развитием остальных частей организма - лишь в этом случае она будет подхвачена естественным отбором. Удлинение клюва вряд ли окажется выгодным, если оно сочетается с утерей языка или отсутствием ушей. Такие катастрофические последствия более вероятны в случае мутаций, затрагивающих ранние стадии индивидуального развития, чем в случае мутаций, влияющих на его поздние этапы. Ранние эмбриональные клетки подобны картам в основании карточного домика - от них зависит слишком многое, и даже незначительное изменение их свойств скорее всего приведет к печальным последствиям. Ранние стадии индивидуального развития оказались «замороженными» - так же точно, как в биохимической организации клеток «заморожены» генетический код и механизмы биосинтеза белка. В отличие от этого клетки, образующиеся на последних стадиях развития, имеют больше возможностей для изменений. Вероятно, именно по этой причине на ранних стадиях развития эмбрионы разных видов столь часто бывают похожи друг на друга и в процессе индивидуального развития, видимо, нередко повторяют пройденные ими этапы эволюции.
1.3.8. Эукариотические организмы имеют сложный аппарат воспроизведения
У многоклеточных организмов должны быть клетки, служащие предшественниками новых поколений. У высших растений и животных эти клетки весьма специализированы и называются первичными половыми или зародышевыми клетками. От них зависит размножение вида, и мощное давление отбора приводит структуру организма в целом в соответствие с требованием максимальной вероятности выживания зародышевых клеток. Прочие клетки могут погибнуть, но до тех пор, пока живы зародышевые клетки, будут появляться новые организмы, аналогичные родительскому. В этом смысле наиболее фундаментальное различие в многоклеточном организме существует между зародышевыми клетками и всеми остальными, т. е. между зародышевыми и соматическими клетками.
Не все многоклеточные организмы воспроизводятся с помощью специальных дифференцированных клеток. Многие простые животные (среди них губки и кишечнополостные) способны размножаться почкованием. Аналогичный способ используется многими растениями. Для полового размножения, однако, необходимы зародышевые клетки. Процесс полового размножения столь хорошо известен, что кажется простым, хотя он несомненно сложнее неполового воспроизведения и требует мобилизации значительных ресурсов. Две особи одного вида, но разного пола обычно производят совершенно различные зародышевые (половые) клетки: одна особь - яйцеклетки, другая - сперматозоиды. Яйцеклетка сливается со сперматозоидом с образованием зиготы - клетки, генотип которой представляет собой случайным образом перетасованный набор генов двух родителей. Практически все виды эукариот, как многоклеточные, так и одноклеточные (даже те, которые могут размножаться другими способами), способны к половому размножению. Развитию полового размножения вместо более простых стратегий, основанных на обыкновенном делении клеток, должны были способствовать мощные силы естественного отбора. Хотя трудно с уверенностью указать все преимущества, которые дает при отборе половое размножение, ясно по крайней мере, что этот процесс создает возможность перетасовывания и комбинирования генов. Половое размножение могло сыграть ведущую роль в эволюции новых генов и их комбинаций и таким образом внести свой вклад в бесконечное разнообразие форм и функций современных растений и животных.
1.3.9. Клетки позвоночных имеют более 200 различных типов специализации
Число различных специализаций, которые можно обнаружить у клеток высших животных, неизмеримо больше, чем у любого прокариотического организма. У позвоночных четко различают более 200 клеточных типов, причем многие из них, видимо, объединяют под общим названием большое количество более тонко различающихся вариантов. На схеме 1-2 представлено лишь несколько типов клеток. Несмотря на чрезвычайное разнообразие функций клеток, даже в пределах одного организма можно убедиться в их удивительной универсальности. Многие современные представления об общих свойствах клеток эукариот основаны на изучении специализированных типов, у которых исключительно сильно развито определенное полезное свойство, в той или иной степени присущее всем клеткам. Каждая особенность и каждая органелла (представленные на схеме 1-1) развиты в необычайно высокой степени или особенно четко выражены у того или другого типа клеток. В качестве произвольного примера рассмотрим нервно-мышечное соединение, в образовании которого участвуют клетки трех типов: мышечные, нервные и шванновские. Клетки каждого типа играют только им присущую роль (рис. 1-37).
1. Специализация мышечной клетки - сокращение. Ее цитоплазма заполнена упорядоченными рядами белковых нитей, в том числе актиновых. В промежутках между ними находится множество митохондрий, поставляющих АТР в качестве топлива для сократительного аппарата.
2. Нервная клетка стимулирует сокращение мышцы, подводя к ней возбуждающий сигнал от головного или спинного мозга. Поэтому нервная клетка необычайно вытянута: ее тело, содержащее ядро, может находиться на расстоянии метра или более от места соединения с мышцей. В процессе эволюции у нервных клеток появился хорошо развитый цитоскелет, необходимый для поддержания столь необычной формы и эффективного транспорта веществ из одного конца клетки в другой. Однако ключевая специализация нервной клетки связана с ее плазматической мембраной, содержащей белки, которые образуют ионные насосы и каналы и вызывают передвижение ионов, эквивалентное электрическому току. Хотя такие насосы и каналы имеются в плазматической мембране всех клеток, только нервная клетка использует их таким образом, чтобы электрический импульс мог распространиться от одного конца клетки до другого за какую-то долю секунды и передать сигнал к действию.
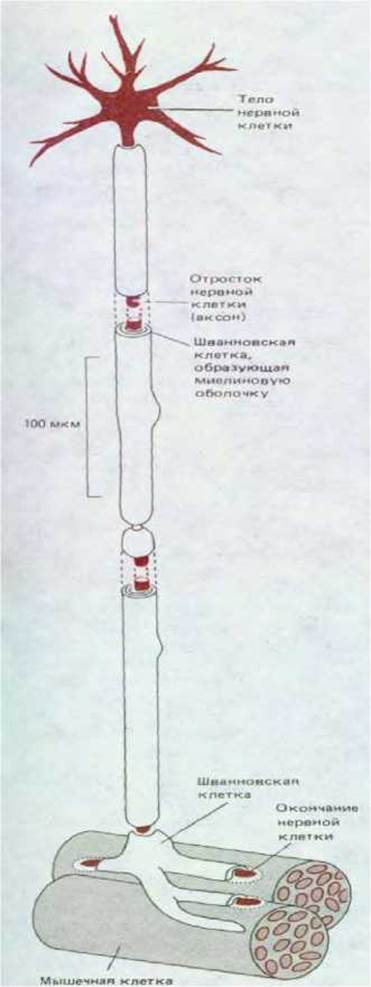
Рис. 1-37. Схематическое изображение нервномышечного соединения: отросток нервной клетки, окруженный миелиновой оболочкой, контактирует с мышечной клеткой.
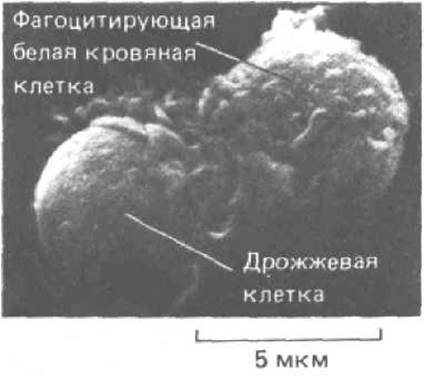
Рис. 1-38. Микрофотография (сканирующий электронный микроскоп) нейтрофила - одного из представителей лейкоцитов - поглощающего дрожжевую клетку. (Из J. Воуlеs, D.F. Bainton, Cell, 24, 905-914, 1981. © Cell Press.)
3. Наконец, шванновские клетки специализированы для массового «производства» плазматической мембраны, которую они слой за слоем, как ленту, навивают на вырост нервной клетки так, что получается служащая изолятором миелиновая оболочка.
1.3.10. Клетки иммунной системы специализируются на химическом узнавании
Из всех имеющихся у высших животных клеточных систем две по сложности и тонкости организации достигли высшей степени развития. Это иммунная система и нервная система позвоночных. Каждая из них далеко превосходит любой искусственный прибор: иммунная система - по способности к химическому распознаванию, а нервная система - по способности к восприятию и управлению. Каждая система состоит из многих различных типов клеток и основана на взаимодействии между ними.
Защищенная от внешних воздействий и тщательно поддерживаемая внутренняя среда многоклеточного организма благоприятна не только для собственных клеток животного - она притягательна также и для посторонних организмов. Следовательно, животным необходимо защитить себя от вторжений, в особенности от вирусов и бактерий. Первоочередная задача иммунной системы состоит в уничтожении любых проникнувших в тело животного чужеродных микроорганизмов.
Как указывалось выше, многие эукариотические клетки способны к фагоцитозу: они могут поглощать извне частицы различных веществ и переваривать их. Среди дифференцированных клеток высших животных есть «специалисты» по фагоцитированию, например макрофаги, способные проглатывать и уничтожать бактерии и другие клетки (рис. 1-38). Но тут возникает одно осложнение: хорошо, когда фагоциты атакуют вторгнувшиеся извне клетки, но для организма было бы губительным, если бы они стали нападать также на близких родственников и коллег. Таким образом, иммунная система должна отличать собственные клетки от чужеродных, т.е. уметь распознавать «свое» и «чужое».
Для этой цели у позвоночных развился специальный класс узнающих клеток - лимфоциты. Сами по себе лимфоциты не являются фагоцитами, но кооперируются с последними, посылая им сигналы, показывающие, следует ли атаковать данную клетку или оставить ее в живых. В частности, некоторые лимфоциты (В-лимфоциты) вырабатывают специальные белковые молекулы - антитела, избирательно связывающиеся с определенными атомными группами на поверхности чужеродных организмов или производимых ими токсичных молекул. Чтобы пометить новый, вторгнувшийся извне организм как чужеродный, должен быть произведен новый класс антител, но, поскольку количество возможных чужеродных организмов очень велико и практически непредсказуемо, В-лимфоциты должны быть способны к синтезу бесконечного разнообразия антител. В то же время иммунная система не должна производить антитела, связывающиеся с собственными молекулами и клетками организма.
Огромное разнообразие антител создается уникальными генетическими механизмами, благодаря их существованию в организме синтезируются миллионы генетически различных лимфоцитов, каждый из них при пролиферации формирует клон, все члены которого вырабатывают одно и то же специфическое антитело. Те клоны из этого множества, которые вырабатывают антитела, реагирующие с собственными молекулами организма, уничтожаются или подавляются (с помощью все еще не ясных механизмов), а те, которые продуцируют антитела против чужеродных молекул, избирательно выживают и размножаются. Таким образом, развитие иммунной системы индивидуального животного, подобно эволюционному процессу, использует стратегию случайных изменений с последующим отбором.
1.3.11. Нервные клетки позволяют организму быстро адаптироваться в изменяющихся условиях [17, 18]
Наличие иммунной системы - признак, свидетельствующий о высокой организации позвоночных и отличающий их от низших животных, у которых лимфоциты отсутствуют. Нервная система, напротив, имеется практически у всех многоклеточных животных. Ее роль состоит в обеспечении еще более фундаментальной потребности - потребности в быстром адаптивном ответе на внешние события.
На протяжении многих поколений эволюция совершенствует структуру организма, доводя ее до оптимальной, и приводит ее в соответствие со средой обитания. Однако в подавляющем большинстве экологических ниш наблюдаются изменения, которые происходят слишком быстро для того, чтобы могла выработаться эволюционная адаптация. В этих условиях наиболее приспособленными окажутся организмы, способные к адаптации иного рода, не требующей генетических мутаций, но тем не менее модифицирующей поведение в соответствии с изменившимися обстоятельствами. Если последовательность изменений окружающей среды полностью предсказуема, наподобие смены дня и ночи или зимы и лета, автономные изменения организма по соответствующему расписанию могут быть генетически запрограммированы. Так, фотосинтезирующая активность принадлежащего к динофлагеллятам одноклеточного организма Gonyaulax (рис. 1-39) обнаруживает 24-часовую периодичность, поддерживающуюся, даже если организм в течение недели содержать в условиях постоянного освещения. Такие биологические часы есть и у многих других организмов, но механизм их действия все еще не разгадан.
Однако большинство изменений, происходящих в окружающей среде, не предсказуемы. Например, бактерии, обитающие в кишечнике, подвергаются нерегулярным колебаниям состава и количества доступного для них питания, и любая бактерия, способная подстраивать свой метаболизм к этим изменениям, будет иметь преимущество над бактерией, не способной к такой реакции. Поэтому бактерии приобрели способность регулировать скорость синтеза своих метаболических ферментов в зависимости от концентрации питательных веществ в окружающей среде. Для сопряжения внешнего стимула с адаптивным ответом служат специальные управляющие молекулы (такие, как циклический AMP, сАМР).
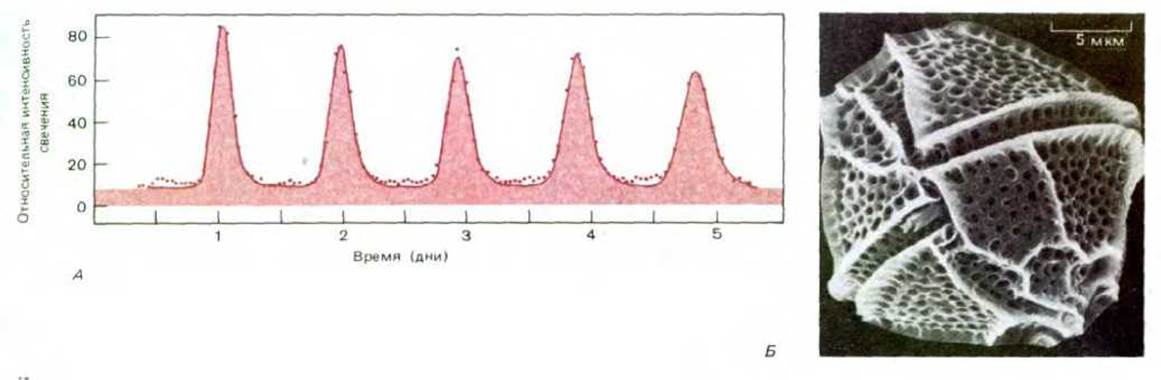
Рис. 1-39. А. Ритм свечения фотосинтезирующего простейшего Gonyaulax polyedra, соотнесенный с теоретической кривой. Средний период ритма свечения равен 22 ч 58 мин, а отклонение для индивидуальных клеток в популяции составляет в сутки ± 18 мин. Б. Микрофотография (сканирующий электронный микроскоп) Gonyaulax. (А - из D. Njus, V.D. Gooch, J. W. Hastings, Cell Biophys., 3, 223-231, 1981; Б - с любезного разрешения John Dolge.)
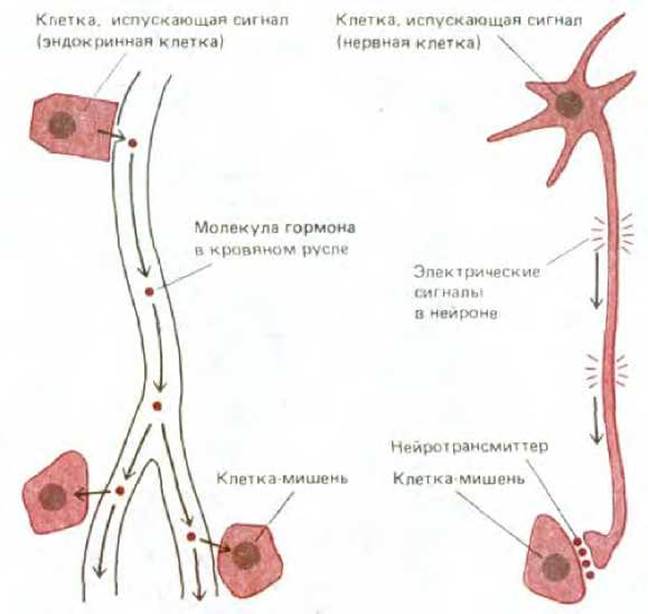
Рис. 1-40. Сравнение гормональной и нервной сигнализации. Эндокринная клетка при стимуляции высвобождает гормоны в кровяное русло, и циркулирующие гормоны вызывают ответ в любой клетке, чувствительной к ним, независимо от того, где она находится. Нервная клетка при стимуляции генерирует потенциал действия, распространяющийся вдоль аксона и быстро запускающий высвобождение нейротрансмиттера с окончания аксона; нейротрансмиттер действует только на клетки, находящиеся в непосредственной близости от окончания аксона.
У многоклеточного организма сигнал, связывающий восприятие с действием, должен, как правило, передаваться от одних клеток к другим. Так, подстройка метаболизма часто осуществляется гормонами, которые, будучи секретированы одной группой клеток, путешествуют в тканях и вызывают ответ в других группах клеток. Но гормону требуется много времени, чтобы пройти большое расстояние, кроме того, он при этом диффундирует во все стороны. Чтобы химический сигнал передавался быстро, он должен испускаться вблизи своей мишени, тем же путем достигается точная локализация действия сигнала. Но если химический сигнал должен испускаться вблизи мишени, то как же можно использовать такой способ связи для сопряжения восприятия с реакцией на него в удаленной части организма? Проблему решают нервные клетки. Один их конец чувствителен к химическим или физическим стимулам, а другой - продуцирует химический сигнал - нейромедиатор, действующий на другие клетки (рис. 1-40). Стимуляция одного конца клетки вызывает электрическое возбуждение, которое быстро распространяется до другого конца, где вызывает высвобождение нейромедиатора. Такое сигнальное устройство позволяет многоклеточным животным быстро реагировать на изменчивый окружающий мир, а также точно координировать активность далеко отстоящих друг от друга частей тела.
1.3.12. Связи между нервными клетками определяют тип поведения
Одиночный нейрон человека не очень отличается от одиночного нейрона червя. Преимущество нервной системы человека основано на огромном количестве входящих в нее клеток и, самое главное, на способе их соединения друг с другом, который и определяет возможности нейронов в передаче, комбинировании и интерпретации сенсорных сигналов, а также в координации сложных последовательностей действий. Чтобы понять клеточные основы эволюции нервной системы, необходимо рассмотреть механизмы, с помощью которых эмбриональные нервные клетки приобретают фантастически сложные формы и достигают строго упорядоченного способа их соединения (рис. 1-41). Некоторое представление об изумительной сложности и организованности сетей, которые сплетают развивающиеся нервные клетки, дает зрительная система мухи (рис. 1-42). Строение всей этой структуры задано генетически, и развитие ее происходит нормально даже в отсутствие света. При анализе развития нервной системы обнаруживается, что у всех видов оно основано на одних и тех же фундаментальных принципах: одинаковые механизмы используются для клеточного движения, межклеточной адгезии, химической сигнализации и т.д. Эволюция сложной нервной системы основывается прежде всего на эволюции сложных контрольных механизмов сочетания и координирования этих главных типов клеточного поведения.
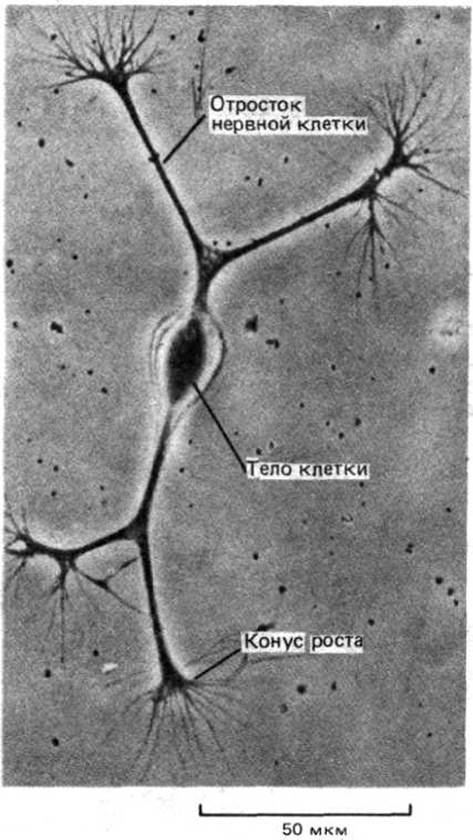
Рис. 1-41. Микрофотография изолированной нервной клетки куриного эмбриона, помещенной в культуральный сосуд с питательным раствором. У клетки появляются длинные выросты, каждый из которых продвигается с помощью структуры, называемой конусом роста. (С любезного разрешения Zoltan Gabor.)
Характер связей между нервными клетками определяет поведение животного. Без всякого обучения и опыта самец мухи спаривается с самкой, паук плетет свою паутину, птицы летят на юг. Все эти действия предначертаны в ДНК этих видов, которая контролирует поведение отдельных клеток при построении нервной системы у эмбриона и работу нервной системы у взрослого организма.
Но не всякое поведение генетически детерминировано, важное значение имеет и собственный опыт животного. Лишение развивающегося млекопитающего сенсорных стимулов может изменить микроструктуру мозга. Взрослые животные почти всех видов - от кишечнополостных до человека - в той или иной степени способны к обучению. По определению обучение - это результат опыта и, следовательно, электрической активности нервных клеток, которая должна приводить к длительным изменениям нейронных связей. Детальная расшифровка этих механизмов является одной из центральных задач современных нейробиологических исследований.
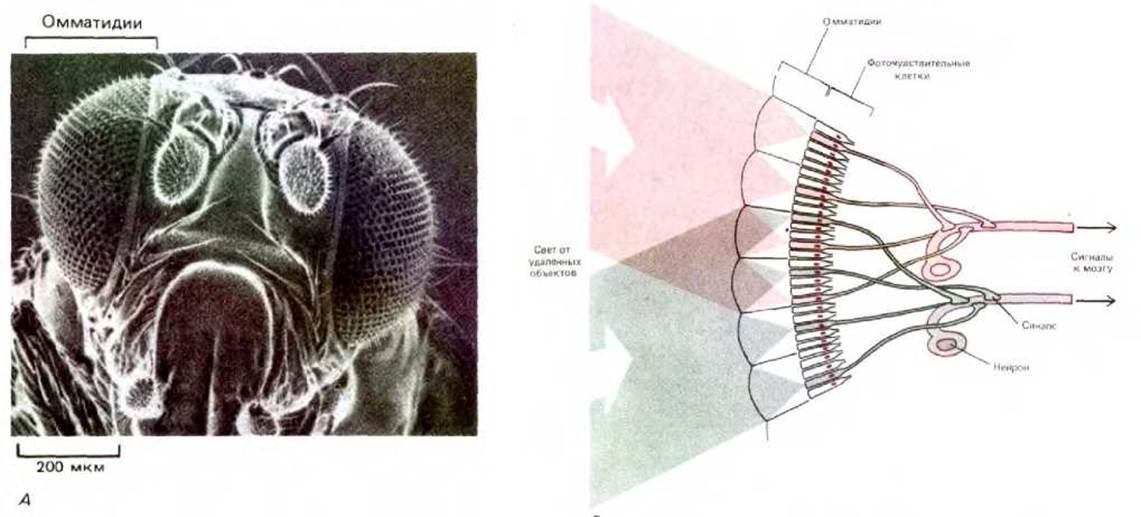
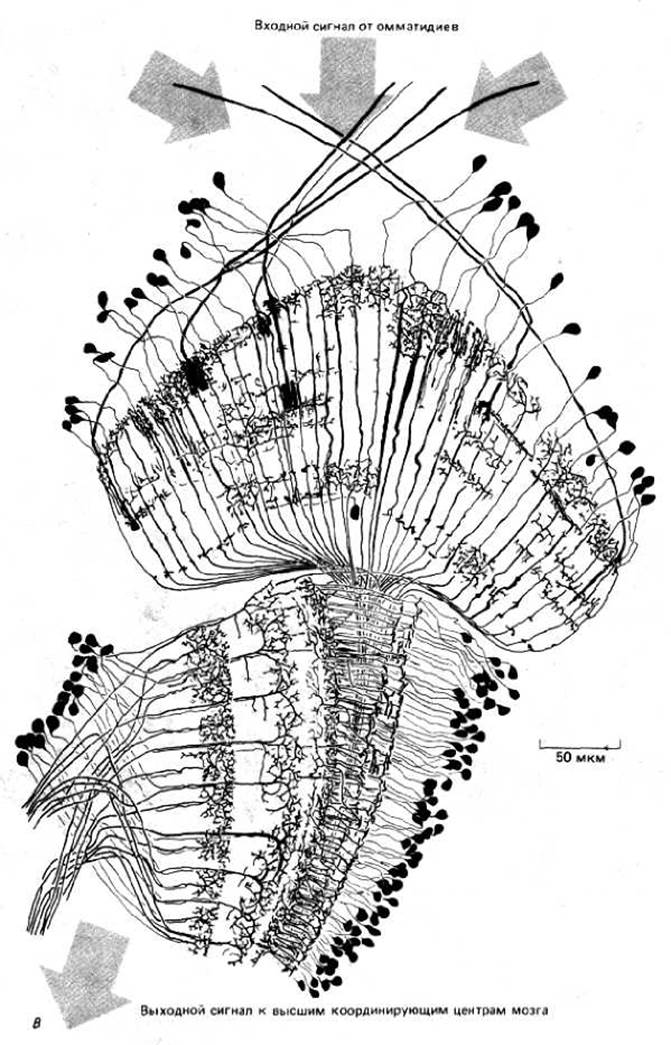
Рис. 1-42. А. Вид в сканирующем электронном микроскопе головы плодовой мушки (Drosophila). С двух сторон головы расположены два больших сложных глаза, состоящие из множества элементов, называемых омматидиями. Каждый омматидий имеет свою линзочку, фокусирующую свет на группу находящихся в его основании клеток-фоторецепторов. Б. Схематическое изображение нейронных связей в наружном слое глаза мухи, видимых на вертикальном срезе. Свет проникает в каждый омматидий сложного глаза и фокусируется на одну из восьми находящихся в основании омматидия фоточувствительных клеток-рецепторов (здесь показаны лишь пять из них). Из-за кривизны сложного глаза свет от удаленного точечного источника фокусируется в разных омматидиях на разные фоточувствительные рецепторы. Однако короткие аксоны фоторецепторов, «смотрящих» на одну и ту же точку, переплетены таким образом, что оказываются подсоединенными к одному и тому же идущему в мозг насекомого пучку аксонов. В каждом глазу мухи присутствует более тысячи таких аксонных пучков, причем каждый из них в ходе индивидуального развития точно присоединяется к правильному набору фоторецепторов. В. Сеть нервных клеток участка мозга мухи, получающего и обрабатывающего входной сигнал от омматидиев. (А - с любезного разрешения Rudi Turner and Antony Mahowald; В - из N. Stausfeld, Atlas of an Insect Brain. New York, Springer, 1976.)
Многие нейронные связи мозга, позволяющие нам читать, писать и говорить, появляются в результате обучения и отражают негенетический тип наследования. Обучение и обмен информацией дали возможность человеку как виду адаптироваться таким способом, который для менее высокоорганизованных существ возможен только лишь путем генетической эволюции. Тем не менее ясно, что даже эти очень сложные способности, лежащие в основе нашей культуры и общества, имеют в своей основе тонкие особенности поведения клеток - правила, по которым нейроны на долгое время модифицируют свои связи в результате электрической активности.
Конечно, изучая лишь единичные клетки, мы не поймем, как устроен многоклеточный организм, точно так же, как мы не поймем устройства клетки, анализируя отдельные биологические молекулы. Но все же, не зная строения клетки, мы не можем рассчитывать на полное понимание работы организма. И мы не можем как следует разобраться в механизмах функционирования клетки, не зная составляющих ее молекул. Таким образом, обсуждение живой клетки, к которому мы переходим в следующей главе, должно начинаться с описания входящих в ее состав молекул.
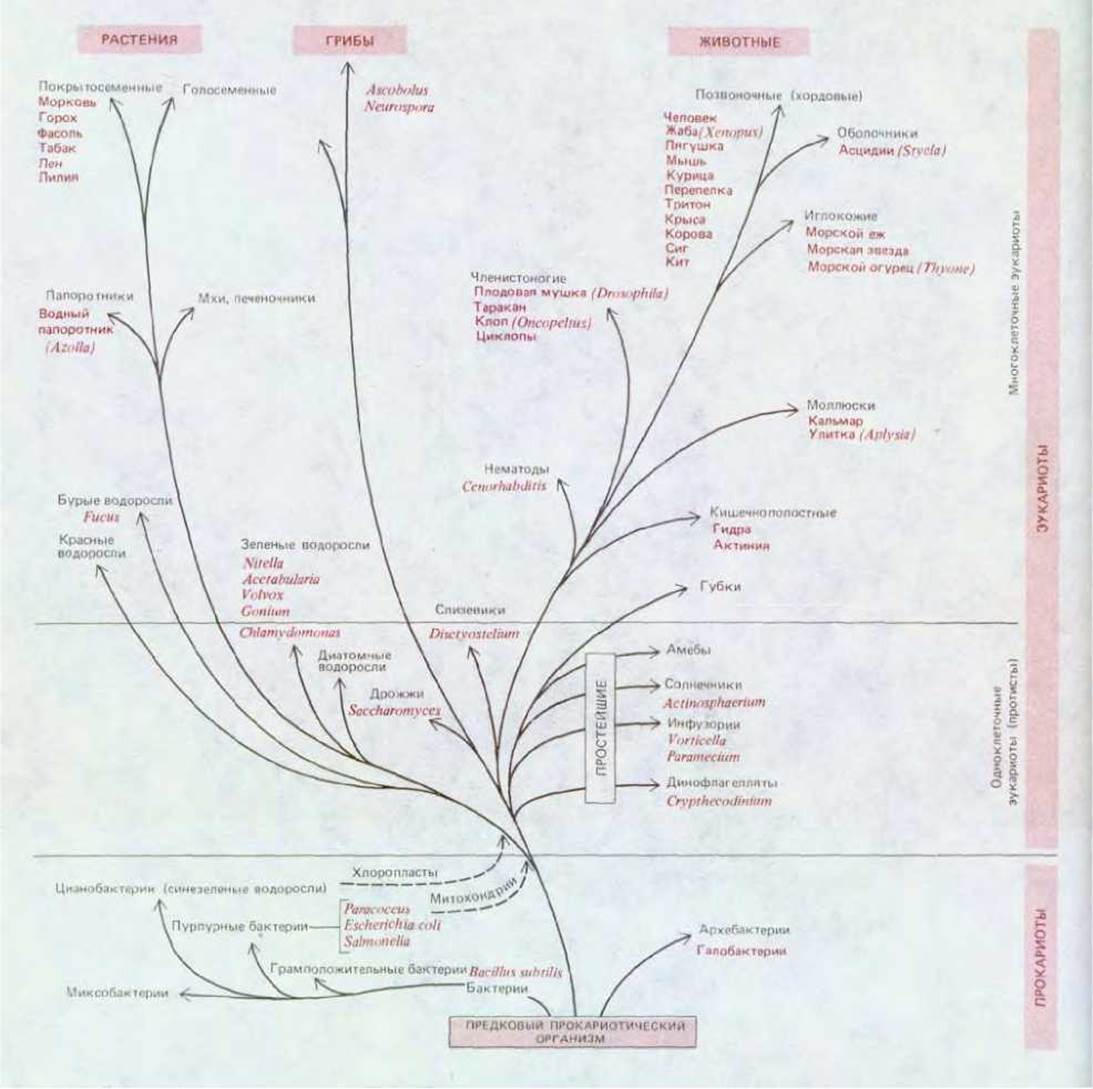
Рис. 1-43. Эволюционное родство между некоторыми упомянутыми в этой книге организмами. Ветви древа показывают пути общего происхождения, но их длина не отражает реального временного масштаба. (Отметим также, что вертикальная ось диаграммы демонстрирует не время, а основные категории организмов.)
Заключение
Эволюция крупных многоклеточных организмов связана со способностью эукариотических клеток по-разному экспрессировать наследственную информацию, а также с умением этих клеток функционировать сообща. Одним из наиболее ранних этапов на пути к многоклеточности было появление эпителия, в котором клетки соединены в слои, отделяющие внутреннюю среду организма от внешнего окружения. Первыми примитивными типами дифференцированных клеток должны были быть наряду с эпителиальными клетками нервные клетки, мышечные клетки и клетки соединительной ткани. Все эти типы клеток можно найти даже у очень примитивных современных животных.
Эволюция высших животных, используя ту же основную стратегию развития, шла в направлении создания все возрастающего числа специализированных клеточных типов и все более утонченных методов координации их активности (рис. 1-43). Две системы клеток высших животных представляют каждая в своем роде вершину сложности многоклеточной организации. Одна - это иммунная система позвоночных, клетки которой способны производить миллионы различных антител. Другая - это нервная система. У низших животных большая часть нейронных связей жестко генетически детерминирована, и программа поведения эволюционирует лишь благодаря мутациям генетического материала. У высших животных работа и структура нервной системы становились все более подверженными модификациям (обучению) благодаря способности нервных клеток изменять свои связи в ответ на вызванную внешними стимулами электрическую активность.
Литература
Общая
Bendall D. S. ed. Evolution from Molecules to Men. Cambridge, UK. Cambridge University Press, 1983.
Curtis H. Biology, 4th ed. New York. Worth, 1983.
Darnell J. E., Lodish H. F., Baltimore D. Molecular Cell Biology, Chapter 25.
New York. W.H. Freeman, 1986. Darwin C. On the Origin of Species. London. Murray, 1859. Reprinted, New York, Penguin, 1984. Evolution. Sci. Am. 239(3), 1978. (An entire issue devoted to the topic.)
Keeton W. Т., Gould J. W., Gould C. G. Biological Science, 4th ed. New York, Norton, 1986.
Maynard Smith J. The Theory of Evolution, 3rd ed. New York, Penguin, 1975.
Watson J. D., Hopkins N. H., Roberts J. W., Steitz J. A., Weiner A. M. Molecular Biology of the Gene, 4th ed. Chapter 28. Menlo Park CA.
Benjamin-Cummings, 1987.
Wilson E. B. The Cell in Development and Heredity, 3rd ed. New York: Macmillan, 1925. (Reprinted, New York: Garland, 1987.)
Цитируемая
1. Ferris J.P., Usher D.A. Origins of life. In: Biochemistry, 2nd ed. (G. Zubay ed.), pp. 1120-1151. New York: Macmillan, 1988.
Miller S. M., Orgel L. E. The Origins of Life on the Earth. Englewood Clifis, NJ. Prentice-Hall. 1974.
Schopf J. W., Hayes J. M., Walter M. R. Evolution of earth's earliest ecosystems: recent progress and unsolved problems. In: Earth's Earliest
Biosphere: Its Origin and Evolution (J. W. Schopf ed.), pp. 361-384. Princeton, NJ. Princeton University Press, 1983.
2. EigenM., Gardiner W., SchusterP., Winkler-OswatitschR. The origin of genetic information. Sci. Am., 244(4), 88-118, 1981.
3. Cech T.R. RNA as an enzyme. Sci. Am. 255(5), 64-75, 1986.
4. Alberts В. М. The function of the hereditary materials: biological catalyses reflect the cell's evolutionary history. Am. Zool. 26, 781 - 796, 1986.
Darnell J. E., Doolittle W. F. Speculations on the early course of evolution. Proc. Natl. Acad. Sci. USA, 83, 1271 - 1275, 1986.
Orgel L.E. RNA catalysis and the origin of life. J. Theor. Biol. 123, 127-149, 1986.
5. Rogers M. et al. Construction of the mycoplasma evolutionary tree from 5S rRNA sequence data. Proc. Natl. Acad. Sci. USA, 82, 1160-1164, 1985.
6. Vidal G. The oldest eukaryotic cells. Sci. Am., 250(2), 48-57, 1984.
7. Clarke P. H. Enzymes in bacterial populations. In: Biochemical Evolution (H. Gutfreund ed.), pp. 116-149. Cambridge UK. Cambridge University Press, 1981.
Doolittle W.F. Archaebacteria coming of age. Trends Genet., 1, 268-269, 1985.
Woese C. R. Bacterial evolution. Microbiol. Rev., 5, 221-271, 1987.
8. Wilson A. The molecular basis of evolution. Sci. Am., 253(4), 164-173, 1985.
9. DickersonR. E. Cytochrome с and the evolution of energy metabolism. Sci. Am., 242(3), 136-153, 1980.
10. Cavalier-Smith T. The origin of eukaryotic and archaebacterial cells. Ann.N.Y. Acad. Sci. 503, 17-54, 1987.
Margulis L. Symbiosis in Cell Evolution. New York, W. H. Freeman, 1981.
11. Vossbrinck C.R., Maddox J.V., Friedman S., Debrunner-Vossbrinck B. A., Woese C. R., Ribosomal RNA sequence suggest microsporidia are extremely ancient eukarvotes. Nature, 326, 411-414, 1987.
Yang D., Oyaizu Y., Oyaizu H., Olsen G. J., Woese C. R. Mitochondrial origins. Proc. Natl. Acad. Sci. USA, 82, 4443-4447. 1985.
12. SleighM. A. The Biology of Protozoa. London, Edward Arnold, 1973.
Sogin M. I., Elwood H. J., Gunderson J. H. Evolutionary diversity of eukaryotic small-subunit rRNA genes. Proc. Natl. Acad. Sci. USA, 83, 1383-1387, 1986.
13. Buchsbaum R. Animals Without Backbones, 2nd ed. Chicago. University of Chicago Press, 1976.
Field K. G. et al. Molecular phytogeny of the animal kingdom. Science, 239, 748-753, 1988.
Margulis L., Schwartz K. V. Five Kingdoms: An illustrated Guide to the Phyla of Life on Earth, 2nd ed. New York. W.H. Freeman, 1987. Shapiro J. A. Bacteria as multicellular organisms. Sci. Am., 258(6), 82-89, 1988.
Valentine J. W. The evolution of multicellular plants and animals. Sci. Am., 239(3), 140-158, 1978.
14. Bode P. M., Bode H. R. Patterning in Hydra. In: Pattern Formation (G. M. Malacinski, S.V. Bryant eds.), pp. 213-244. New York, Macmillan, 1984.
15. Raff R. A., Kaufman T. C. Embryos, Genes, and Evolution. New York. Macmillan, 1983.
16. Maynard Smith J. The Evolution of Sex. Cambridge, UK. Cambridge University Press, 1978.
Michod R. E., Levin B. R. eds. The Evolution of Sex: An Examination of Current Ideas. Sunderland, MA. Sinauer, 1988.
17. Winfree A. T. The Timing of Biological Clocks. New York. W. H. Freeman, 1987.
18. Bullock Т. Н., Orkand R., Grinnell A. Introduction to Nervous Systems. San Francisco. Freeman, 1977.