Молекулярная биология клетки - Том 2 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1993
Внутриклеточная сортировка макромолекул и сохранение клеточных компартментов
Везикулярный транспорт и сохранение индивидуальности компартментов
Внутриклеточная сортировка, по-видимому, требует наличия как минимум 10 различных типов транспортных пузырьков, каждый из которых имеет уникальный набор «молекулярных адресных меток» на поверхности, позволяющих доставлять содержимое пузырьков только к определенной клеточной мембране. Следовательно, транспортные пузырьки, покидающие ЭР. должны сливаться только с цис-компартментом Гольд-жи, покидающие цис-компартмент должны сливаться только с промежуточным компартментом Гольджи и т. д. На каждом этапе происходит отпочковывание, стыковка и слияние пузырька с мембраной. Все эти процессы требуют высокоспецифичрюго узнавания. В настоящее время нам мало известно о молекулярных механизмах (см. рис. 8-73). В этом разделе мы проанализируем некоторые спорные взгляды на пути сохранения внутриклеточной компартментации и опишем некоторые новые экспериментальные подходы к решению этих спорных проблем.
8.10.1. Некоторые белки удерживаются в ЭР и аппарате Гольджи в качестве постоянных компонентов [67]
При перемещении «груза» из одного компартмента в другой транспортные пузырьки обязательно переносят как мембраны, так и содержимое органелл. Тем не менее и при таком выравнивающем процессе сохраняются различия в составе мембран разных компартментов: белок-рецептор SRP встречается только в мембране ЭР, а гликозилтрансферазы и ферменты процессинга олигосахаридов расположены только в мембранах определенных цистерн Гольджи и т. д. Следовательно, мембраны ЭР и каждою типа цистерн Гольджи должны иметь специальные механизмы для сохранения своей уникальности. Один из них - наличие специальных сигналов сортировки для каждого этапа продвижения продукта через ЭР и аппарат Гольджи. В результате, например, белки плазматической мембраны, попадающие в клетку путем специфического эндоцитоза. захватываются окаймленными ямками. Однако существует точка зрения, согласно которой при биосинтетическом транспорте через ЭР и аппарат Гольджи, используется противоположный механизм, т.е. транспорт происходит автоматически, а для удержания продукта в органелле требуются специфические сигналы. В соответствии с этой гипотезой каждый постоянный компонент ЭР или аппарата Гольджи должен иметь специальный сигнал, отвечающий за его сохранение в этом компартменте. Стратегия автоматического движения вперед и избирательного сохранения привлекательна еще и потому, что число белков, проходящих сквозь ЭР и аппарат Гольджи к месту конечного назначения, значительно больше числа белков, остающихся там. Более того, при такой стратегии те белки, которые утратили свои сигналы сортировки, или были направлены в неверном направлении, могут выводиться из клетки. Наконец, если бы сигналы требовались для транспорта, то они были бы необходимы для каждой его стадии - от ЭР к аппарату Гольджи и от каждой цистерны Гольджи к следующей. В этом случае структура многих белков оказалась бы перегруженной большим количеством сигналов сортировки, которые должны находиться на поверхности молекулы.
8-45
8.10.2. Существует по крайней мере два типа окаймленных пузырьков [68]
Клатрин находится на цитоплазматической поверхности плазматической мембраны и транс-сети Гольджи; по-видимому, этот белок участвует в транспорте, управляемом сигналами. Покрытые клатрином пузырьки отвечают за опосредованный рецепторами эндоцитозный путь от плазматической мембраны к эндосомам, а также за управляемый рецепторами путь от транс-сета Гольджи к эндолизосомам. В первом случае окаймленные пузырьки переносят избранный набор рецепторов клеточной поверхности. Вероятно, это верно и для различных типов покрытых клатрином пузырьков, отпочковывающихся от аппарата Гольджи.
Окаймленные клатрином пузырьки не отделяются ни от ЭР, ни от цис- или промежуточных цистерн аппарата Гольджи. Вместо них гам (а также и в транс-сети Гольджи) существует другой тип окаймленных пузырьков. Они не окрашиваются антителами к клатрину и отличаются от клатриновых окаймленных пузырьков при анализе в электронном микроскопе (рис. 8-83). Белки «каймы» этих пузырьков еще не идентифицированы.
Возможная модель белкового транспорта в неполяризованных клетках представлена на рис. 8-84. В соответствии с этой моделью, белки «по умолчанию» перемещаются из ЭР в аппарат Гольджи, от одной цистерны Гольджи к другой и от транс-сети Гольджи к поверхности клетки. Предполагают, что этот неизбирательный процесс происходит при помощи неклатриновых окаймленных пузырьков. От транс-сети Гольджи управляемый рецепторами транспорт в лизосомы происходит с участием клатриновых окаймленных пузырьков, а транспорт к секреторным гранулам - с участием частично покрытых клатрином пузырьков. Каким образом можно проверить эту модель?
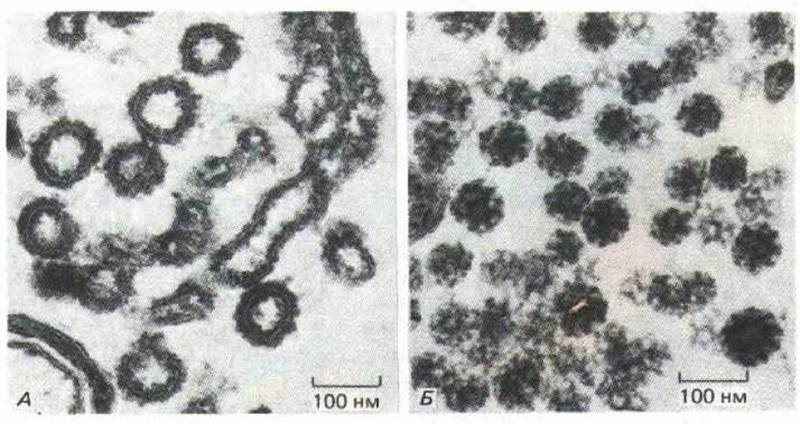
Рис. 8-83. Сравнение клатриновых и неклатриновых окаймленных пузырьков. А, Электронная микрофотография цистерн Гольджи из бесклеточной системы, в которой происходит отпочковывание неклатриновых окаймленных пузырьков в пробирке. Клатриновые окаймленные пузырьки, показанные на фото (Б), имеют гораздо более упорядоченное строение. (Электронные микрофотографии любезно предоставлены Lelio Orci, см. L. Оrci, В. Click, and J. Rolhman, Cell, 46: 171-184.)
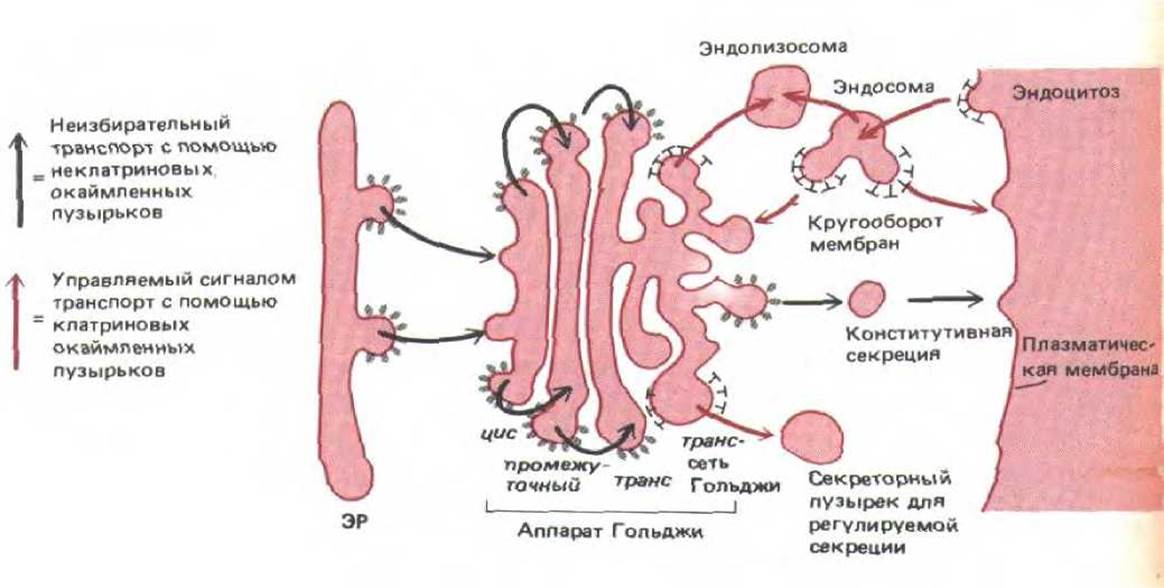
Рис. 8-84. Гипотетическая модель транспорта белков в неполяризованных клетках. Полагают, что неизбирательный (конститутивный) транспорт происходит при помощи неклатриновых окаймленных пузырьков, а различные варианты регулируемого сигналами транспорта при помощи клатриновых окаймленных пузырьков. Эти пузырьки отпочковываются от транс-сети Гольджи, эндосом и плазматической мембраны. В поляризованных клетках необходим еще дополнительный регулируемый сигналами транспортного от транс-сети Гольджи.
8.10.3. Раскрыть механизм транспорта помогают мутантные клетки с нарушенной секрецией [69]
Так как транспорт исключительно важен для эукариотической клетки, то мутантные клетки, неспособные осуществлять один из его этапов (например, дефектные по маркеру стыковки или акцепторному белку), должны были бы гибнуть. Однако, если мутантный белок не функционирует только при высоких температурах, в обычных условиях клетка с таким дефектом вполне жизнеспособна. У дрожжей было выявлено более 25 температурочувствительных (термочувствительных) мутаций в генах, отвечающих за секрецию. Клетки, несущие такие мутации, не способны при высоких температурах транспортировать белки из ЭР в аппарат Гольджи, от одной цистерны Гольджи к другой, из аппарата Гольджи к вакуоли или к плазматической мембране.
Используя температурочувствительные мутанты дрожжей, легко клонировать гены этого организма, ответственные за транспорт. Такой подход чрезвычайно плодотворен, т. к. позволяет напрямую выявить главные белки транспортного механизма, не зная даже, как происходит секреция. Один из дрожжевых генов, отвечающих за секрецию, который был выявлен таким образом - это ген sec 4. Полагают, что он кодирует GTP-связывающий белок из семейства ras (см. разд. 12.3.11 и 13.4.6). Биохимические эксперименты на клетках млекопитающих свидетельствуют о том, что сходный GTP-связывающий белок участвует в везикулярном транспорте и у высших эукариот. Возможно, он регулирует «раздевание» неклатриновых окаймленных пузырьков перед их связыванием с мембранами.
8.10.4. Бесклеточные системы дают другой плодотворный подход к изучению молекулярных механизмов везикулярного транспорта [70]
Чтобы понять молекулярные механизмы, лежащие в основе потока веществ между мембранными компартментами, надо выявить основные «рабочие части» транспортных пузырьков. Каким образом транспортные пузырьки отпочковываются от мембраны? Что направляет их к мембранам-мишеням? Как они сливаются с мембранами? Кроме только что описанных генетических экспериментов для ответа на эти вопросы были использованы опыты по реконструкции везикулярного транспорта в бесклеточной системе. Впервые этого удалось добиться для стопки Гольджи. Когда выделенные стопки Гольджи инкубировали[3H]-GlcNAc из UDP-[3H]-GlcNAc, добавленного в культуральную среду. Как показано на графике справа, транспорт идет только тогда, когда добавляется и цитозольный экстракт, и АТР. Такие же условия необходимы для отпочковывания неклатриновых окаймленных пузырьков. Такая схема опыта, впервые примененная для того, чтобы понять механизм переноса из цис-компартмента Гольджи в его промежуточный компартмент, позволила также воссоздать in vitro транспорт из ЭР в цис-аппарат Гольджи, из промежуточного в транс-компартмент Гольджи, от трансаппарата Гольджи к плазматической мембране, от эндосом к лизосомам, и от транс-аппарата Гольджи к эндолизосомам.
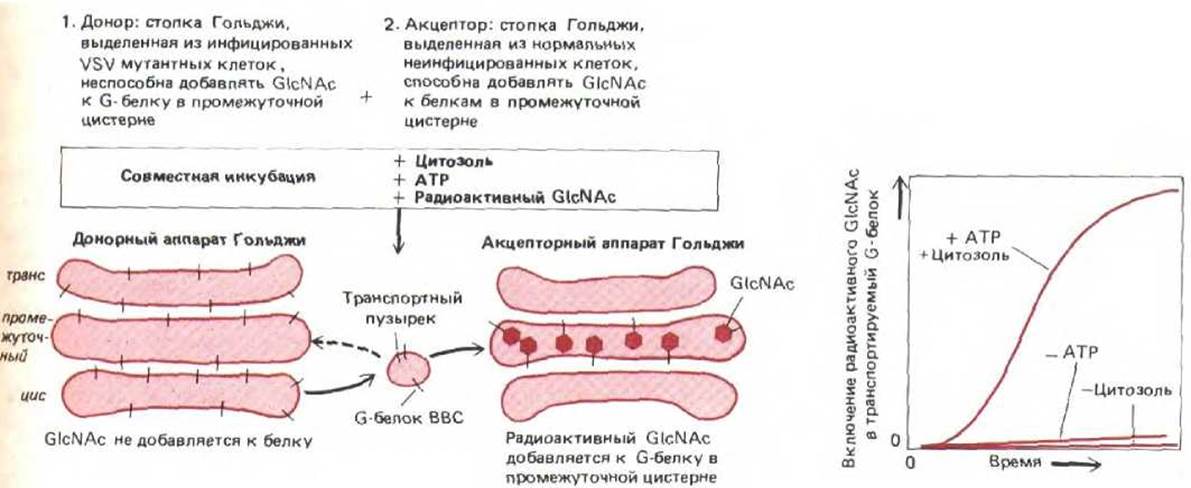
Рис. 8-85. Бесклеточная система, воссоздающая везикулярный транспорт вирусного белка между цистернами аппарата Гольджи. Когда препарат стопок Гольджи инкубируют с цитозольными экстрактами и АТР, от цистерн отпочковываются неклатриновые окаймленные пузырьки. Вирус везикулярного стоматита (VSV) кодирует белок оболочки, называемый G-белком. Чтобы понять, как происходит транспорт отого трансмембранного белка между цистернами аппарата Гольджи, две популяции выделенных стопок Гольджи инкубировали вместе. Популяция-«донор» была получена из инфицированных VSV мутантных клеток, не имеющих GlcNAc-трансферазы I. (поэтому в «донорной» популяции стопок Гольджи не происходит включения остатков GlcNAc в N-связанные олигосахариды G-белка). «Акцепторные» стопки Гольджи были получены из неинфицированных клеток дикого типа и поэтому содержали нормальную копию GlcNAc-трансферазы, но не содержали G-белка. Следовательно, для присоединения к G-белку GlcNAc нужно, чтобы этот белок из цис-компартмента донорной стопки Гольджи попал в промежуточный компартмент стопки-акцептора. Проверить, идет ли сопряженное с транспортом гликозилирование, можно, наблюдая за включением в G-белок
с цитозолем и АТР, от их краев отшнуровывались неклатриновые окаймленные пузырьки, которые, по-видимому, переносили белки между цистернами (рис. 8-83, А). Прослеживая последовательный процессинг олигосахаридов на гликопротеинах по мере их продвижения из одного компартмента Гольджи к другому, стало возможным реконструировать in vitro процесс везикулярного транспорта (рис. 8-85).
Было обнаружено, что отпочковывание неклатриновых окаймленных пузырьков в такой бесклеточной системе требует наличия смеси белков из цитозоля и АТР, из чего следует, что это активный процесс, а не простая самосборка. Необходимые для транспорта компоненты эволюционно крайне консервативны, так как в экспериментах по реконструкции цитозоль животной клетки можно с успехом заменить цитозолем дрожжей или растений. Таким образом, для идентификации молекул, участвующих в везикулярном транспорте, можно сочетать генетический подход, используемый для клеток дрожжей, и бесклеточные системы млекопитающих.
Заключение
ЭР и каждый компартмент аппарата Гольджи содержат уникальные наборы белков. По-видимому, эти белки задерживаются в органеллах благодаря имеющимся у них специальным сигналам. При этом большая часть веществ переносится от ЭР к аппарату Гольджи, сквозь стопку Гольджи и от транс-сети Гольджи к поверхности клетки автоматически. Высказано предположение, что транспорт при таком неизбирательном пути происходит с участием неклатриновых окаймленных пузырьков, которые не специализируются в зависимости от содержимого. В то мое время транспорт, управляемый сигналами (сортировка), опосредован покрытыми клатрином окаймленными пузырьками. Чтобы проверить эти гипотезы, необходимо расшифровать молекулярные механизмы, участвующие в отпочковывании транспортных пузырьков, направлении их к мишеням и слиянии с мембранами. В генетических экспериментах на дрожжах идентифицировано более 25 генов, продукты которых отвечают за отдельные стадии транспорта. Кроме того, для млекопитающих разработаны бесклеточные системы, в которых происходило избирательное отпочковывание и слияние пузырьков. Вероятно, именно сочетание генетического и биохимического подходов позволит в будущем выделить в чистом виде многие белки, участвующие в этих процессах.
Литература
Общая
Burgess Т. L., Kelly R. В. Constitutive and regulated secretion of proteins. Annu. Rev. Cell. Biol., 3, 243-293, 1987.
Dingwall C., Laskey R. A. Protein import into the cell nucleus. Annu. Rev. Cell Biol., 2, 367-390, 1986.
Kornfeld S. Trafficking of lysosomal enzymes. FASEB J., 1, 462-468, 1987.
Pfeffer S. R., Rothman J. E. Biosynthetic protein transport and sorting by the endoplasmic reticulum and Golgi. Annu. Rev. Biochem., 56, 829-852, 1987.
Verner K., Schatz G. Protein translocation across membranes. Science, 241, 1307-1313, 1988.
Цитированная
1. BolenderR. P. Stereological analysis of the guinea pig pancreas. J. Cell Biol., 61, 269 287. 1974.
Palade J. E., Farquhar M. G. Cell biology. In: Pathophysiology: The Biological Principles of Disease (L. H. Smith, S. D. Thier, eds.), pp. 1-56.
Philadelphia, Saunders, 1981.
Weibel E. R., Staubli W., GnagiH.R., Hess F. A. Correlated morphometric and biochemical studies on the liver cell. J. Cell Biol., 42, 68 91, 1969.
2. Blobel G. Intracellular protein topogenesis. Proc. Nail. Acad. Sci. USA, 77, 1496-1500, 1980.
GravM. W., Doolinle W. F. Has the endosymbiont hypothesis been proven? Microbiol. Rev., 46, 1-42, 1982.
Schwarz R. M., Davhoff M. 0. Origins of the prokaryotes, eukaryotes, mitochondria, and chloroplasts. Science, 199, 395-403, 1978.
3. Palade G. Intracellular aspects of the process of protein synthesis. Science, 189, 347-358, 1975.
4. Kelly R. B. Pathways of protein secretion in eukaryotes. Science, 230, 25-31, 1985.
Sabatini D. D., Kreibich G., Morimoto Т., Adesnik M. Mechanisms for the incorporation of proteins in the membranes and organelles. J. Cell Biol., 92, 1 -22, 1982.
5. Pfeffer S. R., Rothman J. E. Biosynthetic protein transport and sorting by the endoplasmic reticulum and Golgi. Annu. Rev. Biochem., 56, 829 852, 1987.
Wickner W. Т., Lodish H.E. Multiple mechanisms of protein insertion into and across membranes. Science, 230, 400-406, 1985.
6. Blobel G. Intracellular protein topogenesis. Proc. Natl. Acad. Sci. USA, 77, 1496-1500, 1980.
GaroffH. Using recombinant DNA techniques to study protein targeting in the eukaryotic cell. Annu. Rev. Cell Biol., 1, 403-445, 1985.
7. Warren G. Membrane traffic and organelle division. Trends Biochem. Sci., 10, 439-443, 1985.
8. Alien R.D. The microtubule as an intracellular engine. Sci. Am., 238(2), 42-49, 1987.
Fulton A.B. How crowded is the cytoplasm? Cell, 30, 345-347, 1982.
Lubv-Phelps K., Taylor D. L., Lanni F. Probing the structure of cytoplasm. J. Cell Biol., 102, 2015 2022, 1986.
ValeR. D. Intracellular transport using microtubule-based motors. Annu. Rev. Cell Biol., 3, 347-378, 1987.
9. ChockP. В., Rhee S. G., Stadtman E. R. Interconvertible enzyme cascades in cellular regulation. Annu. Rev. Biochem., 49, 813-843, 1980.
Holt G. D. et a/. Nuclear pore complex glycoproteins contain cytoplasmically disposed O-linked N-acetylglucosamine. J. Cell Biol., 104, 11571164, 1987.
WoldF. In vivo chemical modification of proteins (post-translational modification). Annu. Rev. Biochem., 50, 783-814, 1981.
10. Kamps M.P., Buss J.E., Sefton B.M. Mutation of NH2-terminal glycine of p60src prevents both myristoylation and morphological transformation. Proc. Ntl. Acad. Sci. USA, 82, 4625-4628, 1985.
Schutltz A. M., Henderson L. E., Orozlan S. Fatty acylation of proteins. Annu. Rev. Cell Biol., 4, 611-648, 1988.
Willumsen B. M., Harris K., Papayeorye A. G., Hubbert N. L., Lowy D. R. Harvey murine sarcoma virus p21ras protein: biological and biochemical significance of the cysteine nearest the carboxy terminus. EM BO J., 3, 2582-2585, 1984.
11. Dice J. F. Molecular determinants of protein half-lives in eukaryotic cells. FASEB J., 1, 349-357, 1987.
Goldbera A.L., GoffS.A. The selective degradation of abnormal proteins in bacteria. In: Maximizing Gene Expression (W. Reznikoff, L. Gold, eds.), pp. 287-314. Stoneham, MA, Butterworth, 1986.
12. Bachmair A., Finley D., Varshavsky A. In vivo half-life of a protein is a function of its amino-terminal residue. Science, 234, 179 186, 1986. Hershko A., Ciechanover A. The ubiquitin pathway for the degradation of intranuclear proteins. Prog. Nucleic Acid Res. Мої. Biol., 33, 19-56, 1986.
RechsteinerM. Ubiquitin-mediated pathways for intracellular proteolysis. Annu. Rev. Cell Biol., 3, 1 30, 1987.
13. Auyen J., Wold F. How much sequence information is needed for the regulation of amino-terminal acetylation of eukaryotic proteins? Trends Biochem. Sci, 11, 494-497, 1986.
Ferber S., Ciechanover A. Role of arginine-tRNA in protein degradation by the ubiquitin pathway. Nature, 326, 808-811, 1987.
Varshavsky A., Bachmair A., Finlev D., Gonda D., Wunniny I. The N-end rule of selective protein turnover: mechanistic aspects and functional implications. In: Ubiquitin (M. Rechsteiner, ed.), pp. 284 324. New York, Plenum, 1988.
14. Craiy E.A. The heat-shock response. CRC Crit. Rev. Biochem., 18, 239-280, 1985.
Lindquist S. The heat-shock response. Annu. Rev. Biochem., 55, 1151-1191, 1986.
Pelham H. R. B. Speculations on the functions of the major hat shock and glucose-regulated proteins. Cell, 46, 959-961, 1986.
15. Franke W.W., Scheer U.. Krohne G., Jarash E.D. The nuclear envelope and the architecture of the nuclear periphery. J. Cell Biol., 91, 39s 50s, 1981.
Newport J. W., Forbes D.J. The nucleus: structure, function, and dynamics. Annu. Rev. Biochem., 56, 535 565, 1987.
16. Banner W.M. Protein migration and accumulation in nuclei. In: The Cell Nucleus (H. Busch, ed.), Vol. 6, Part C, pp. 97-148. New York, Academic, 1978.
Lang L, Schol: M., Peters R. Molecular mobility and nucleocytoplasmic flux in hepatoma cells. J. Cell Biol., 102, 1183-1190, 1986.
17. Dingwall C., Laskey R.A. Protein import into the cell nucleus. Annu. Rev. Cell Biol., 2, 367-390, 1986.
Feldherr C. M., Kallenbach E., Schult: N. Movement of a karyophilic protein through the nuclear pores of oocytes. J. Cell Bol., 99, 2216-2222, 1984.
Newmeyer D.D., Forbes D.J. Nuclear import can be separated into distinct steps in vitro: nuclear pore binding and translocation. Cell, 52, 641653, 1988.
18. Goldfarb D.S., Gariepy J., Schoolnik G., Kornbery R.D. Synthetic peptides as nuclear localization signals. Nature, 322, 641-644, 1986.
Kalderon D., Roberts B. L., Richardson W. D., Smith A. E. A short amino acid sequence able to specify nuclear location. Cell, 39, 499-509, 1984.
Lanford R. E., Bute/ J. S. Construction and characherization of an SV40 mutant defective in nuclear transport of Т antigen. Cell, 37, 801-813, 1984.
19. Clawson G.A., Feldherr C.M., Smuckler E. A. Nucleocytoplasmic RNA transport. Мої. Cell. Biochem., 67, 87-100, 1985.
Dworetzky S. I., Feldherr С. М. Translocation of RNA-coated gold particles through the nuclear pores of oocytes. J. Cell Biol., 106, 575 584, 1988.
20. Attardi G., Schatz G. Biogenesis of mitochondria. Annu. Rev. Cell Biol., 4, 289-333, 1988.
Tzayoloff A. Mitochondria. New York, Plenum, 1982.
21. Hawlitschek G. et al. Mitochondrial protein import: identification of processing peptidase and of PEP, a procesing enchancing protein. Cell, 53, 795-806, 1988.
Hurt E.G., van Loon A.P.G.M. How proteins fnd mitochondria and intramitochondrial compartments. Trends Biochem. Sci. 11, 204-207, 1986. PfannerN.. Neupert W. Biogenesis of mitochondrial energy transducing complexes. Curr. Top. Bioenerg., 15, 177-219, 1987.
Raise D., Schatz G. Mitochondrial presequences. J. Biol. Chem., 263, 4509-4511, 1988.
22. Eilers M., Schatz G. Protein unfolding and the energetics of protein translocation across biological membranes. Cell, 52, 481-483, 1988.
Pfanner N.. Neupert W. Transport of proteins into mitochondria: a potassium diffusion potential is able to drive the import of ADP/ATP carrier. EM BO J., 4, 2819-2825, 1985.
Raise D., Horvath S.J., Tomich J.M., Richards J.H., Schatz G. A chemically synthesized pre-sequence of an imported mitochondrial protein can form an amphiphilic helix and perturb natural and artificial phospholipid bilayers. EM BO J., 5. 1327 1334. 1986.
23. Schleyer M., Neupert W. Transport of proteins into mitochondria: translocational intermediates spanning contact sites between outer and inner membranes. Cell, 43, 339-350, 1985.
Schwaiger M., Herzog V., Neupert W. Characterization of translocation contact sites involved in the import of mitochondrial proteins. J. Cell Biol., 105, 235-246, 1987.
24. Deshaies R. J., Koch B. D., Werner-Washburne M., Craig E. A., Schekman R. A subfamily of stress proteins facilitates translocation of secretory and mitochonrial precursor polypeptides. Nature, 332, 800-805, 1988.
Eilers M., Schatz G. Binding of a specific ligand inhibits import of a purified precursor protein into mitochondria, Nature, 322, 228-232, 1986. Pfanner N.. Tropschug M., Neupert W. Mitochondrial protein import: nucleoside triphosphates are involved in conferring import competence to precursors. Cell, 49, 815-823, 1987.
25. Hartl F. U., Ostermann J., Guiard В., Neupert W. Successive translocation into and out of the mitochondrial matrix: targeting of proteins to the intermembrane space by a bipartite signal peptids. Cell, 51, 1027-1037, 1987.
van Loon A. P. G. M., Brandli A. W., Schatz G. The presequences of two imported mitochondrial proteins contain information for intracellular and intramitochon-drial sorting. Cell, 44, 801-812, 1986.
26. PfallerR., Neupert W. High-affinity binding sites involved in the import of porin into mitochondria. EMBO J., 6, 2635 2642, 1987.
Pfanner N. et al. Role of ATP in mitochondrial protein import. J. Biol. Chem., 263, 4049-4051, 1988.
27. Boutry M., Nagy F., Polsen G., Aoyagi K., Chua N. H. Targeting of bacterial chloramphenicol acetyltranferase to mitochondria in transgenic plants. Nature, 328, 340 342, 1987.
Pain D., Kanwar Y.S., Blobel G. Identification of a receptor for protein import into chloroplasts and its localization to envelope contact zones. Nature, 331, 232-237, 1988.
Schmidt G. W., Mishkind M. L. The transport of proteins into chloroplasts. Annu. Rev. Biochem. 55, 879-912, 1986.
Smeekens S., Bauerie C., Hageman J., Keegstra K., Weisbeek P. The role of the transit peptide in the routing of precursors toward different chloroplast compartments. Cell 46, 365 375, 1986.
28. de Duve C. Microbodies in the living cell. Sci. Am., 248(5), 74 84, 1983. de Duve C.. Baudhuin P. Peroxisomes (microbodies and related particles). Physiol. Rev. 46, 323-357, 1966.
Fahimi H. D., Sies H., eds. Peroxisomes in Biology and Medicine. Heidelberg, Springer, 1987.
29. Tolbert N. E.. Essner E. Microbodies: peroxisomes and glyoxysomes. J. Cell Biol., 91, 271s-283s, 1981.
Veenbuis M., Van Dijken J. P., Harder W. The significance of peroxisomes in the metabolism of one-carbon compounds in yeasts. Adv. Microb. Physiol., 24, 1 -82, 1983.
30. Could S. J., KellerGA., Subramani S. Identification of a peroxisomal targeting signal at the carboxy terminus of four peroxisomal proteins. J. Cell Biol., 107, 897-905, 1988.
Imanaka Т., Small G. M., Lazarów P. B. Translocation of acyl-CoA oxidase into peroxisomes requires ATP hydrolysis but not a membrane potential. J. Cell Biol., 105, 2915-2922, 1987.
Lazarów P. В., Fujiki Y. Biogenesis of peroxisomes. Annu. Rev. Cell Biol., 1, 489-530, 1985.
31. DePierre J. W., Dallner G. Structural aspects of the membrane of the endoplasmicreticulum. Biochim. Biophys. Acta, 415, 411-472, 1975.
FawcettD. The Cell, 2nd ed., pp. 303-352. Philadelphia, Saunders, 1981.
Lee C., BoChen, L. Dynamic behavior of endoplasmic reticulum in living cells. Cell, 54, 37-46, 1988.
32. Adelman M. R.. Sabatini D. D., Blobel G. Ribosome-membrane interaction: nondestructive disassembly of rat liver rough microsomes into
ribosomal and membranous components. J. Cell Biol., 56, 206-229, 1973.
Blobel G., Dohberstein B. Transfer of proteins across membranes. J. Cell Biol., 67, 852-862, 1975.
33. Jones A. L., Fawcett D. W. Hypertrophy of the agranular endoplasmic reticulum in hamster liver induced by phenobarbital. J. Histochem. Cytochem., 14, 215-232, 1966.
Mori H., Christensen A. K. Morphometric analysis of Leydig cells in the normal rat testis. J. Cell Biol., 84, 340-354, 1980.
34. Dallner G. Isolation of rough and smooth microsomes - general. Methods Enzymol., 31, 191-201, 1974.
de Duve C. Tissue fractionation past and present. J. Cell Biol., 50, 20d 55d, 1971.
35. Hortsch M., Avossa D., Meyer D.I. Characterization of secretory protein transloca-tion: ribosome-membrane interaction in endoplasmic reticulum. J. Cell Biol., 103, 241-253, 1986.
Kreibich G., Ulrick B. L., Sabatini D. D. Proteins of rough microsomal membranes related to ribosome binding. J. Cell Biol., 77, 464 487, 1978.
36. Blobel G., Dobberstein B. Transfer of proteins across membranes. J. Cell Biol., 67, 835-851, 1975.
Garoff H. Using recombinant DNA techniques to study protein targeting in the eucaryotic cell. Annu. Rev. Cell Biol., 1, 403-445, 1985. Milstein C., Brownlee G., Harrison Т., Mathews M. B. A possible precursor of immunoglobulin light chains. Nature New Biol., 239, 117-120, 1972. van Heijne G. Signal sequences: the limits of variation. J. Мої. Biol., 184, 99-105, 1985.
37. Meyer D. L, Krause E., Dobberstein B. Secretory protein translocation across membranes-the role of the "docking protein". Nature, 297, 647 650, 1982.
Tajima S., Lauffer L., Rath V. L.. Walter P. The signal recognition particle receptor is a complex that contains two distinct polypeptide chains. J. Cell Biol., 103, 1167-1178, 1986.
Walter P., Blobel G. Signal recognition particle contains a 7S RNA essential for protein translocation across the endoplasmic reticulum.
Nature, 299, 691-698, 1982.
Walter P., Lingappa V. R. Mechanism of protein translocation across the endoplasmic reticulum membrane. Annu. Rev. Cell Biol., 2, 499-516, 1986.
Wiedmann M., Kurzchalia T. V., Hartmann E., Rapoport T. A. A signal sequence receptor in the endoplasmic reticulum membrane. Nature, 328, 830- 833, 1987.
38. Chirico W.J., Waters M. G., Blobel G. 70K heat shock related proteins stimulate protein translocation into microsomes. Nature, 332, 805-810, 1988.
Perara E., Rothman R. E., Lingappa, V. R. Uncoupling translocation from translation: implications for transport of proteins across membranes. Science, 232, 348-352, 1986.
Zimmermann R., Meyer D.I. 1986 A year of new insights into how proteins cross membranes. Trends Biochem. Sci., 11, 512-515, 1986.
39. Rapoport T. A. Extensions of the signal hypothesis sequential insertion model versus amphipatic tunnel hypothesis. FEES Lett., 187, 1-10, 1985.
Wickner W.T., LodishH.F. Multiple mechanisms of protein insertion into and across membranes. Science, 230, 400-406, 1985.
40. Engelman D. M., Steitz T. A.. Goldman A. Identifying nonpolar transbilayer helices in amino acid sequences of membrane proteins. Annu. Rev.
Biophys. Biophys. Chem., 15, 321-353, 1986.
Kaiser C. A., Preuss D., Grisafl P., Botstein D. Many random sequences functionally replace the secretion signal sequence of yeast invertase. Science, 235, 312-317, 1987.
Kyte J., Doolittle R. F. A simple method for displaying the hydropathic character of a protein. J. Мої. Biol., 157, 105-132, 1982.
Zerial M., Huylebroeck D., Garoff H. Foreign transmembrane peptides replacing the internal signal sequence of transferrin receptor allow its translocation and membrane binding. Cell, 48, 147 155, 1987.
41. Bole D. G., Hendershof L.M., KearnyJ.F. Posttranslational association of immunoglobulin heavy chain binding protein with nascent heavy chains in nonsecrcting and secreting hybridomas. J. Cell Bol., 102, 1558 1566, 1986.
Lodish H. F. Transport of secretory and membrane glycoproteins from the rough endoplasmic reticulum to the Golgi. J. Biol. Chem., 263, 2107-2110, 1988.
Мито S., Pelham H. R. B. A C-terminal signal prevents secretion of luminal ER proteins. Cell, 48, 899 907, 1987.
42. Freedman R. Native disulphide bond formation in protein biosynthesis: evidence for the role of protein disulphide isomerase. Trends Biochem., Sci., 9, 438-441, 1984.
Holmgren A. Thioredoxin. Annu. Rev. Biochem., 54, 237-272, 1985.
43. Hirschberg С. В.. Snider M. D. Topography of glycosylation in the rough endoplasmic reticulum and Golgi apparatus. Annu. Rev. Biochem., 56, 63-87, 1987.
Kornfeld R., Kornfeld S. Assembly of asparagine-linked oligosaccharides. Annu. Rev. Biochem., 54, 631 664, 1985.
Torres C., Hart G. Topography and polypeptide distribution of terminal N-acetyl-glucosamine residues on the suface on intact lymphocytes. J. Biol. Chem., 259, 3308-3317, 1984.
44. Cross G. A. M. Eukaryotic protein modification and membrane attachment via phosphatidylinositol. Cell, 48, 179-181, 1987.
Ferguson M. A.J., Williams A. F. Cell-surface anchoring of proteins via glycosil-phosphatidylinositol structures. Annu. Rev. Biochem., 57, 285-320, 1988.
Loif M. G., Sa/tiel A. R. Structural and functional roles of glycosil-phosphatidylino-sitol in membranes. Science, 239, 268-275, 1988.
45. Bishop W. R., Bell R. M. Assembly of phospholipids into cellular membranes: biosynthesis, transmembrane movement, and intracellular translocation. Annu. Rev. Cell Biol., 4, 579-610, 1988.
Bishop W. R., Bell R. M. Assembly of the endoplasmic reticulum phospholipid bilayer: the phosphatidylcholine transporter. Cell, 42, 51-60, 1985.
Dawidowic: E. A. Dynamics of membrane lipid metabolism and turnover. Annu. Rev. Biochem., 56, 43-61, 1987.
Pagano R. E., Sleight R. G. Defining lipid transport pathways in animal cells. Science, 229, 1051 1057, 1985.
Rothman J.E., Lenard J. Membrane asymmetry. Science, 195, 743-753, 1977.
46. Dawidowic: E. A. Lipid exchange: transmembrane movement, spontaneous movement, and protein-mediated transfer of lipids and cholesterol.
Curr. Top. Memb. Transp., 29, 175-202, 1987.
Yaffe M. P.. Kennedy E. P. Intracellular phospholipid movement and the role of phospholipid transfer proteins in animal cells. Biochemistry, 22, 1497-1507, 1983.
47. FarqulmrM.G., PaladeG.E. The Golgi apparatus (complex)-(1954 1981)-from artifact to center stage. J. Cell. Biol., 91, 77s-103s, 1981.
PavelkaM. Functional morphology of the Golgi apparatus. Adv. Anat. EmbryolCell Biol, 106, 1-94, 1987.
Rothman J. E. The compartmental organization of the Golgi apparatus Sci Am 253(3), 74-89, 1985.
48. HubbardS. C, IvattR. J. Synthesis and processing of asparagine-linked oligosaccharides. Annu. Rev. Biochem., 50, 555-583, 1981.
KornfeldR., KornfeldS. Assembly of asparagine-linked oligosaccharides. Annu Rev. Biochem., 54, 631 664, 1985.
Schachter H., Roseman S. Mammalian glycosyltransferases: their role in the synthesis and function of complex carbohydrates and glicolipids. In: The Biochemistry of Glycoproteins and Proteoglycanes (W.J. Lennarz, ed.), Chapter 3 New York Plenum, 1980. '
49. Elbein A. D. Inhibitors of the biosynthesis and processing of N-linked oligosacchande chains. Annu. Rev. Biochem., 56, 497-534, 1987.
StanleyP. Glycosylation mutants and the functions of mammalian carbohydrates Trends Genet., 3, 77-81, 1987.
West C. M. Current ideas on the significance of protein glycosylation. Мої Cell Biochem., 72, 3-20, 1986.
50. Hassel J. R.. Kimura J. H., Hascal V. C. Proteoglycan core protein families Annu Rev. Biochem., 55, 539-567, 1986.
Huttner W. B. Tyrosine sulfation and the secretory pathway. Annu. Rev. Physiol., 50, 363 - 376, 1998.
RuoslahtiF. Structure and biology of proteoglycans. Annu. Rev. Cell Biol., 4, 229-255, 1988.
WaghP. V., Bahl O. P. Sugar residues on proteins. CRC Crit. Rev. Biochem., 10, 307-377, 1981,
51. Douglass J., Civelli O., Herbert E. Polyprotein gene expression: generation of diversity of neuroendocrine peptides. Annu. Rev. Biochem., 53,
665-715, Orel L. et al. Conversion of proinsulin to insulin occurs coordinately with acidication of maturing secretory vesicles. J. Cell Biol., 103, 22732281, 1986.
52. Dunphy W. G., Rothman J. E. Compartmental organization of the Golgi stack. Cell, 42, 13-21, 1985.
53. Bainton D. The discovery of lysosomes. J. Cell Biol., 91, 66s- 76s, 1981. de Duve C. Exploring cells with a centrifuge. Science, 189, 186 194, 1975.
54. Holzman E. Lysosomes: A Survey. New York, Springer-Verlag, 1976.
55. Griffiths G., Holfack В., Simons K., Mellman /., Kornfeld S. The mannose 6-phosphate receptor and the biogenesis of lysosomes. Cell, 52, 329 341, 1988.
Helenius A., Mellman /., Wall D., Hubbard A. Endosomes. Trends Biochem. Sci., 8, 245-250, 1983.
Mayer R.J., Dohertv F. Intracellular protein catabolism: state of the art. FEBS Lett., 198, 181-193, 1986.
Mellman I., Fuchs R., Helenius A. Acidification of the endocytic and exocytic pathways. Annu. Rev. Biochem., 55, 663-700, 1986.
Silverstein S.C., Steinman R.M., Cohn Z.A. Endocytosis. AnhurRev. Biochem., 46, 669-722, 1977.
56. Dahms N. M., Label P., Breitmeyer J., Chirgwin J. M., Kornfeld S. 46 kd mannose 6-phosphate receptor: cloning, expression, and homology to the 215 kd mannose 6-phosphate receptor. Cell, 50, 181 192, 1987.
Kornfeld S. Trafficking of lysosomal enzymes. FASEB J., 1, 462-468, 1987.
Pfeffer S. R. Mannose 6-phosphate receptors and their role in targeting of proteins to lysosomes. J. Membr. Biol., 103, 7-16, 1988. vanFiquraK., HasilikA. Lysosomal enzymes and their receptors. Annu. Rev. Biochem., 55, 167-193, 1986.
57. Brown W.J., Goodhouse J.; Farquhar. M.G. Mannose 6-phosphate receptors for lysosomal enzymes cycle between the Golgi coplex and endosomes. J. Cell Biol., 103, 1235-1247, 1986.
Duncan J. R., Kornfeld S. Intracellular movement of two mannose 6-phosphate receptors: return to the Golgi apparatus. J. Cell Biol., 106, 617628, 1988.
Geuze H.J., Slot J. W., Strous G.J.A.M., Hasilik A., van Figura K. Possible pathways for lysosomal enzyme delivery. J. Cell Biol., 101, 22532262, 1985.
Rothman J. E., Schmid S. L. Enzymatic recycling of clathrin from coated vesicles. Cell, 46, 5-9, 1986.
58. Lane/L., Reitman M., Tang J., Roberts R. M., Kornfeld S. Lysosomal enzyme phosphorylation. J. Biol. Chem., 259, 14663-14671, 1984.
Reitman M. L., Kornfeld S. Lysosomal enzyme targeting. N-acetylglucosaminyl-phosphotransferase selectively phosphorylates native lysosomal enzymes. J. Biol. Chem., 256, 11977 11980, 1981.
59. Kornfeld S. Trafficking of lysosomal enzymes in normal and disease states. J. Clin. Invest., 77, 1-6, 1986.
Neufeld E. F., Lim T. W., ShapiroL. J. Inherited disorders of lysosomal metabolism. Annu. Rev. Biochem. 44, 357 376, 1975.
60. Burgess T. L., Kelly R. B. Constitutive and regulated secretion of proteins. Annu. Rev. Cell Biol., 3, 243-293, 1987.
61. Griffiths G., Simons K. The trans-Golgi network: sorting at the exit site of the Golgi complex. Science, 234, 438-443, 1986.
Orci L. et a/. The trans-most cisternae of the Golgi complex: a compartment for sorting of secretory and plasma membrane proteins. Cell, 51, 1039-1051, 1987.
62. Herzoy V., FarquharM. G. Luminal membrane retrieved after exocytosis reaches most Golgi cisternae in secretory cells. Proc. Natl. Acad. Sci.
USA, 74, 5073-5077, 1977.
Snider M. D.. Rogers О. С. Membrane traffic in animal cells: cellular glycoproteins return to the site of Golgi mannosidase I. J. Cell Biol., 103, 265-275, 1986.
63. LodishH. F. Transport of secretory and membrane glycoproteins from the rough endoplasmic reticulum to the Golgi. J. Biol. Chem., 263, 2107-2110, 1988.
Rothman J. E. Protein sorting by selective retention in the endoplasmic reticulum and Golgi stack. Cell, 50, 521-522, 1987.
Wieland F. Т., Gleason M. L., Serafini T. A., Rothman J. E. The rate of bulk flow from the endoplasmic reticulum to the cell surface. Cell, 50, 289-300, 1987.
64. Bartles J. R.. Hubbard A. L. Plasma membrane protein sorting in epithelial cells: do secretory pathways hold the key? Trends Biochem. Sci., 13, 181 184, 1988.
Matlin K. S. The sorting of proteins to the plasma membrane in epithelial cells. J. Cell Biol., 103, 2565 2568, 1986.
Mostov K. E., Breitfeld P., Harris J. M. An anchor-minus form of the polymeric immunoglobulin receptor is secreted predominantly apically in Madin-Darby canine kidney cells. J. Cell Biol., 105, 2031-2036, 1987.
Simons K., Fuller S. D. Cell surface polarity in epithelia. Annu. Rev. Cell Biol., 1, 243-288, 1985.
65. Simons К., GaroffH., Helenius A. How an animal virus gets into and out of its.host cell. Sci. Am., 246(2), 58-66, 1982.
Simons K., Warren G. Semliki forest virus: a probe for membrane traffic in the animal cell. Adv. Protein Chem., 36, 79-132, 1984.
66. Rodriguez-Boulan F. J. Membrane biogenesis, enveloped RNA viruses, and epithelial polarity. In: Modern Cell Biology. Vol. 1 (J. R. Mcintosh, В. Н. Satir, eds.), pp. 119 170, 1983.
Rodriguez-Boulan E. J., Sabatini D. D. Asymmetric budding of viruses in epithelial monolayers: a model system for the study of epithelial polarity. Proc. Natl. Acad. Sci. USA, 75, 5071-5075, 1978.
Roth M. G., Sirnivas R. V., Compans R. W. Basolateral maturation of retroviruses in polarized epithelial cells. J. Virol., 45, 1065-1073, 1983. Strauss E. G., Strauss J. H. Assembly of enveloped animal viruses. In: Virus Structure and Assembly (S. Casjens, ed.), Chapter 6. Boston, Jones and Bartlett, 1985.
67. Rothman J. E. Protein sorting by selective retention in the endoplasmic reticulum and Golgi stack. Cell, 50, 521-522, 1987.
68. Griffiths G., Pfeiffer S., Simons K., Mat/in K. Exit of newly synthesized membrane proteins from the trans cisterna of the Golgi complex to the plasma membrane. J. Cell Biol., 101, 949-964, 1985.
Orci L., Click B. S., Rothman J. E. A new type of coated vesicular carrier that appears not to contain clathrin: its possible role in protein transport within the Golgi stack. Cell, 46, 171-184, 1986.
69. Bourne H. Do GTPases direct membrane traffic in secretion? Cell, 53, 669-671, 1988.
Novick P., Field C., Schekman R. Identification of 23 complementation groups required for post-translational events in the yeast secretory pathway. Cell, 21, 205-215, 1980.
Scheckman R. Protein localization and membrane traffic in yeast. Annu. Rev. Cell Biol., 1, 115-143, 1985.
70. Batch W. E., Dunphy W. G., Braell W. A., Rothman J. E. Reconstitution of the transport of protein between successive compartments of the
Golgi measured by the coupled incorporation of N-acetylglucosamine. Cell, 39, 405-416, 1984.
Dunphy W. G. et al. Yeast and mamals utilize similar cytosolic components to drive protein transport through the Golgi complex. Proc. Natl. Acad. Sci. USA, 83, 1622-1626, 1986.
Fries E., Rothman J. E. Transport of vesicular stomatitis virus glycoprotein in a cell-free extract. Proc. Natl. Acad. Sci. USA, 77, 3870-3874, 1980.