Молекулярная биология клетки - Том 2 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1993
Межклеточная сигнализация
Три стратегии химической сигнализации: использование гормонов, локальных химических медиаторов и нейромедиаторов
Химические сигнальные механизмы различаются по расстояниям, на которых они действуют: 1) в случае эндокринной сигнализации специализированные эндокринные клетки выделяют гормоны, которые разносятся кровью и воздействуют на клетки-мишени, находящиеся иногда в самых разных частях организма; 2) в случае паракринной сигнализации клетки выделяют локальные химические медиаторы, которые поглощаются, разрушаются или иммобилизуются так быстро, что успевают подействовать только на клетки ближайшего окружения, быть может, в радиусе около миллиметра; 3) при синаптической передаче, используемой только в нервной системе, клетки секретируют нейромедиаторы в специализированных межклеточных контактах, называемых химическими синапсами, Нейромедиаторы диффундируют через синаптическую щель, обычно на расстояние около 50 нм, и воздействуют только на одну постсинаптическую клетку-мишень (рис. 12-2). В каждом случае мишень реагирует на определенный внеклеточный сигнал с помощью специальных белков, называемых рецепторами, которые связывают сигнальную молекулу и инициируют ответ. Многие сигнальные молекулы и рецепторы используются в передаче сигнала и по эндокринному, и по паракринному, и по синаптическому типу. Главные различия касаются быстроты и избирательности воздействия сигнала на определенные мишени.
12-3
12-4
12.1.1. Эндокринные клетки и нервные клетки специализированы для разных типов химической сигнализации [1]
Эндокринные и нервные клетки совместно координируют разнообразные функции миллиардов клеток, из которых состоит тело у высших животных. Эндокринные клетки обычно собраны в специальные железы и выделяют свои гормоны во внеклеточную (интерстициальную) жидкость, окружающую все клетки в тканях. Отсюда молекулы диффундируют в капилляры и разносятся с кровью по всему телу. В каждой ткани они проникают из капилляров в интерстициальную жидкость и могут связываться клетками-мишенями. Поскольку распространение эндокринного сигнала определяется диффузией и кровотоком, оно происходит сравнительно медленно: обычно требуются минуты, чтобы гормон достиг своей мишени. Кроме того, специфичность сигналов в эндокринной системе всецело зависит от химической природы сигнального вещества и рецепторов на поверхности клетки-мишени: каждый тип эндокринной клетки секретирует в кровь свой гормон, и любая клетка, имеющая комплементарный рецептор для этого гормона, ответит реакцией, характерной для данной клетки (рис. 12-3, А).
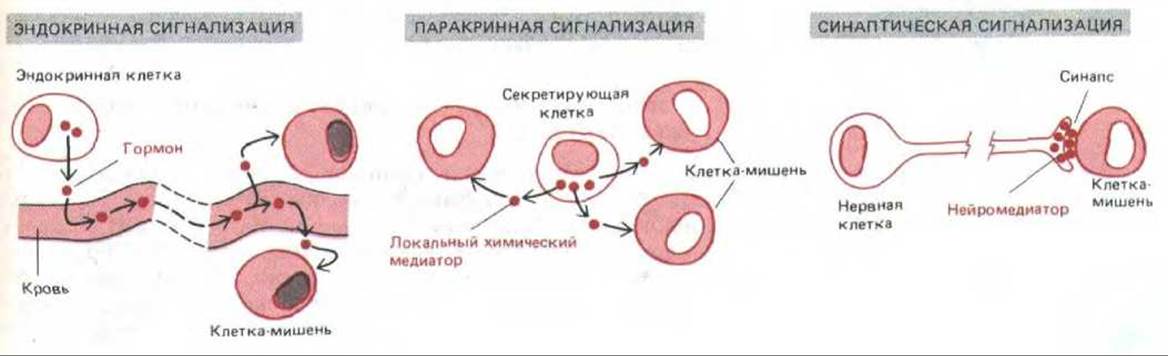
Рис. 12-2. Три формы сигнализации с помощью секретируемых молекул. Не все нейромедиаторы действуют в синапсах, как показано на рисунке; некоторые из них работают как локальные химические медиаторы (по паракринному типу), влияя сразу на целую группу соседних клеток-мишеней.
Работа нервных клеток отличается гораздо большей быстротой и точностью. Они могут передавать информацию на большие расстояния по нервному волокну с помощью электрических импульсов со скоростью более 100 м/с. Только в нервных окончаниях, где высвобождается нейромедиатор, эти импульсы преобразуются в химические сигналы. Химический сигнал нервной клетки может действовать как паракринный или как синаптический. В первом случае нейромедиатор, подобно локальному химическому медиатору, диффундирует наружу и влияет на все соседние клетки-мишени, у которых имеется надлежащий рецептор. При синаптической передаче сигнал гораздо более точен и действие нейромедиатора ограничено единственной клеткой-мишенью, даже если соседние клетки имеют рецепторы для того же нейромедиатора (рис. 12-3, Б). Поскольку расстояние, на которое нейромедиатор должен в таких случаях диффундировать, меньше 100 нм, процесс длится менее миллисекунды (рис. 12-2).
Гормоны в крови и интерстициальной жидкости очень сильно разбавляются, и поэтому они должны быть способны действовать в чрезвычайно низких концентрациях (обычно менее 10-8 М); разбавление же нейромедиаторов на их коротком пути незначительно, и их концентрация около мембраны постсинаптической клетки может быть сравнительно высокой. Например, концентрация ацетилхолина в синаптической щели нервно-мышечного соединения составляет около 5-10-4 М. В соответствии с этим рецепторы нейромедиатора в синапсе обладают относительно низким сродством к своему лиганду и не могут заметным образом реагировать на низкие концентрации нейромедиатора, приходящего путем диффузии от соседних синапсов. Нейромедиатор быстро удаляется из синаптической щели специальными гидролитическими ферментами или мембранными транспортными белками, которые перекачивают его обратно в нервное окончание. Этим достигается точность воздействия сигнала не только в пространстве, но и во времени: короткое, «импульсное» освобождение нейромедиатора вызывает быстрый и краткий ответ, что позволяет сохранять временные характеристики сигнала при передаче его от клетки к клетке.
12.1.2. Главным регулятором эндокринной системы служит гипоталамус [1, 2]
В определенном участке мозга-гипоталамусе-эндокринная и нервная системы физически и функционально связаны друг с другом. Гипоталамус расположен непосредственно над гипофизом, с которым соединен ножкой гипофиза. Гипоталамус выполняет свою связующую роль с помощью клеток, сочетающих особенности нейрона и эндокринной клетки; соответственно их называют нейроэндокринными (нейросекреторными) клетками. Большая часть таких клеток гипоталамуса отвечает на стимуляцию их нейронами других отделов мозга, выделяя в кровеносные сосуды гипофизарной ножки определенный пептидный гормон, который затем специфически стимулирует или подавляет секрецию гипофизом другого гормона. (Другие нейроэндокринные клетки гипоталамуса посылают аксоны в гипофиз и через них выделяют свои секреты в общий кровоток.) Многие из гормонов гипофиза, выделяемых под контролем гипоталамуса, стимулируют какую-либо из других эндокринных желез и вызывают секрецию в кровь третьего гормона. Таким образом, гипоталамус служит у позвоночных главным регулятором эндокринной системы. В качестве примера на рис. 12-4 показано, каким образом он регулирует секреторную функцию щитовидной железы.
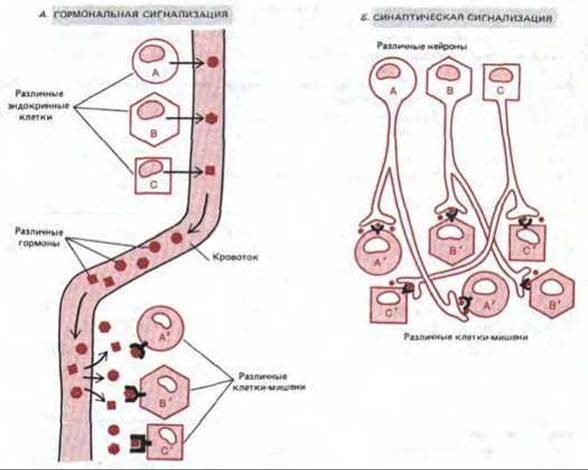
Рис. 12-3. Различие между гормональной (А) и синаптической (Б) передачей сигналов. Эндокринные клетки выделяют в кровь множество гормонов, и клетки-мишени, чувствительные к данному гормону, т.е. имеющие рецепторы для его связывания, «вылавливают» соответствующий гормон из внеклеточной жидкости. При синаптической же передаче специфичность определяется тесным контактом между окончанием нервного волокна и той клеткой-мишенью, которой это волокно передает сигнал: медиатор, выделяемый нервным окончанием, достигает только этой клетки. Для специфической коммуникации с различными клетками-мишенями разные эндокринные клетки должны использовать разные гормоны, в то время как многие нервные клетки могут использовать один и тот же нейромедиатор, сохраняя при этом специфичность воздействия.
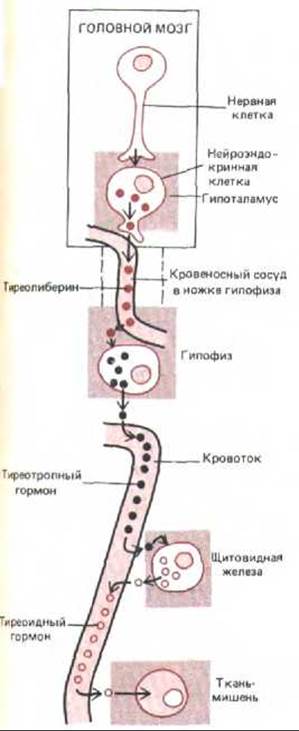
Рис. 12-4. Секреция тиреоидного гормона опосредованно регулируется нервной системой. Определенные нейросекреторные клетки гипоталамуса при стимуляции их нейронами высших отделов мозга выделяют в кровеносные сосуды ножки гипофиза тиреолиберин, который, воздействуя на специфические клетки гипофиза, заставляет их высвобождать тиреотропный гормон (ТТГ). ТТГ током крови транспортируется к щитовидной железе и побуждает ее клетки синтезировать и выделять тиреоидный гормон. Этот гормон стимулирует разнообразные метаболические процессы в большинстве клеток организма. При этом повышение концентрации тиреоидного гормона в крови тормозит секрецию тиреолиберина и ТТГ (на схеме не показано). Эта отрицательная обратная связь препятствует чрезмерному повышению уровня тиреоидного гормона в крови. Подобного рода механизм регулирует секрецию многих гормонов.
Таблица 12-1. Некоторые примеры внеклеточных сигнальных молекул
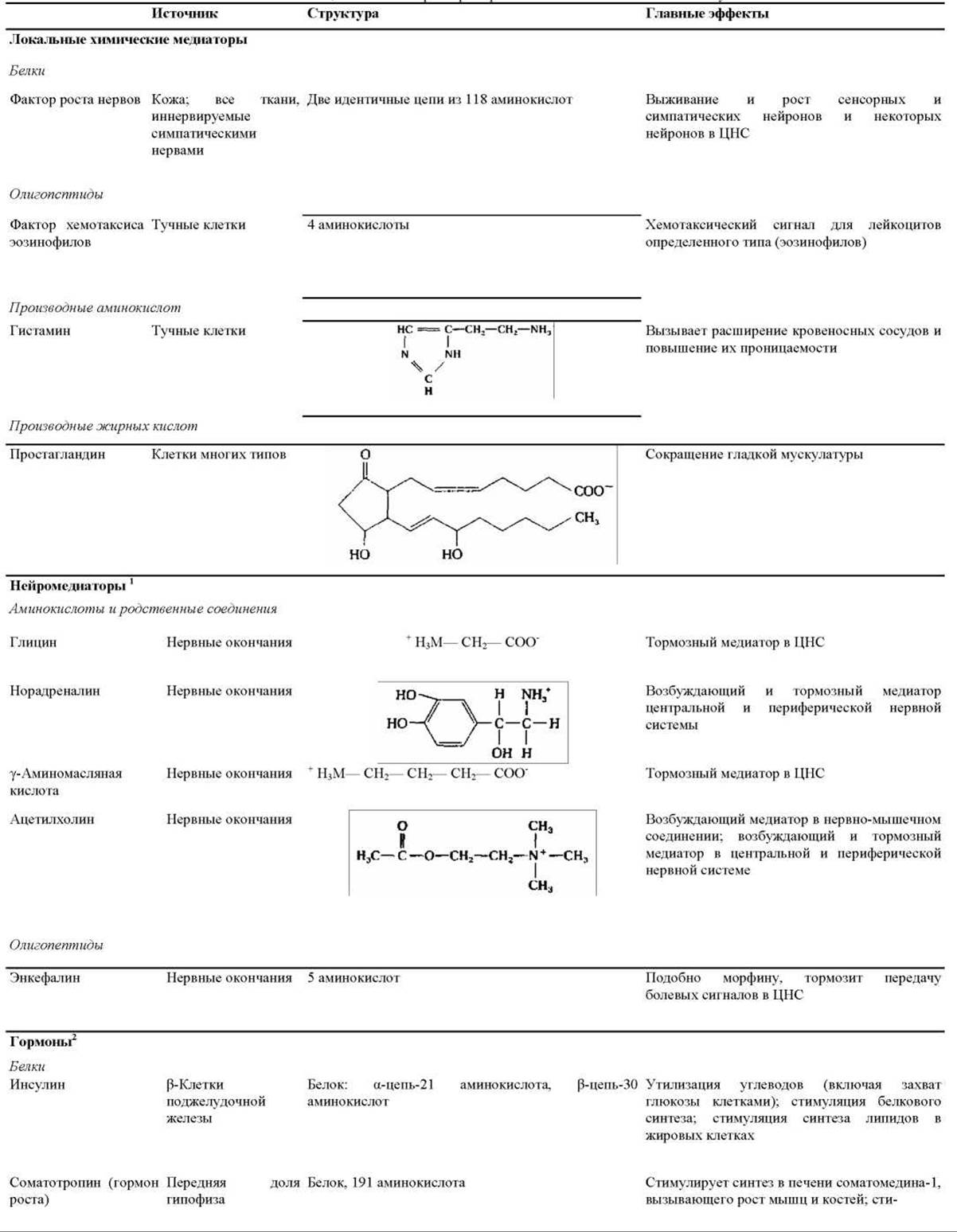
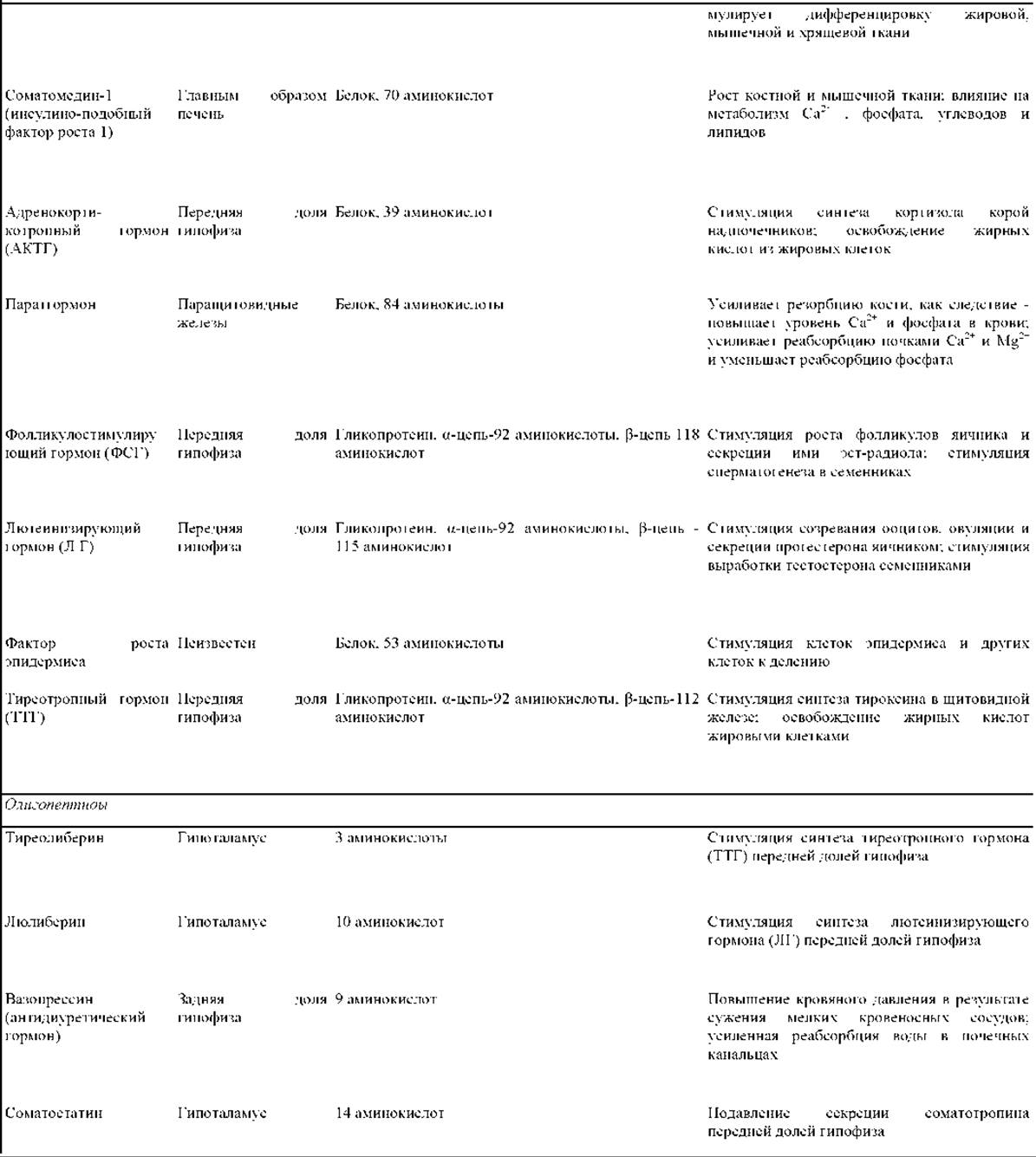
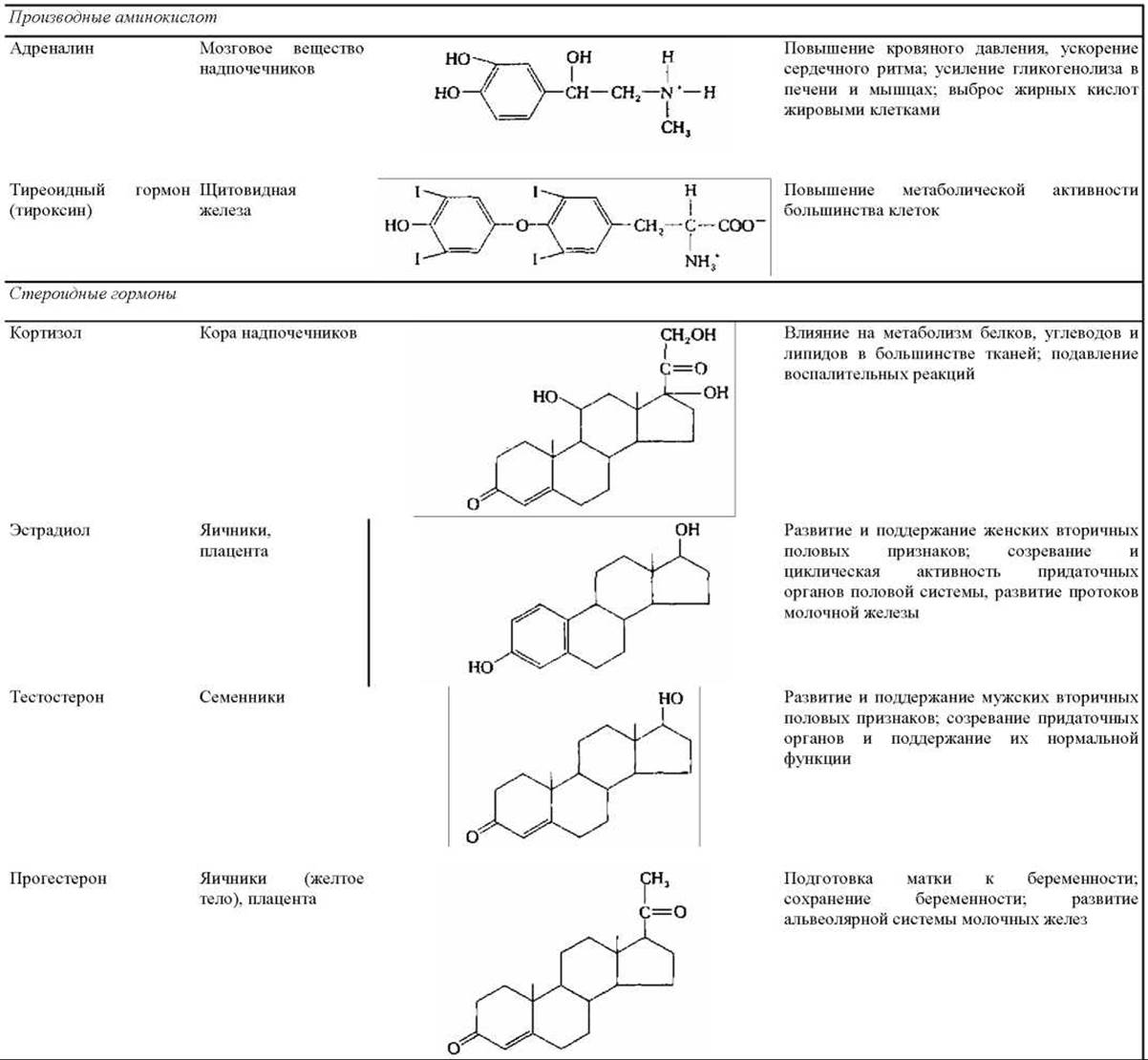
1 Норадреналин и энкефалин чаще действуют по паракринному, чем по синаптическому типу; ацетилхолин может действовать и тем и другим способом. Возбуждающие нейромедиаторы стимулируют активность клеток-мишеней, а тормозные подавляют ее.
2 Многие нестероидные гормоны синтезируются также некоторыми нейронами головного мозга.
В табл. 12-1 приведены сведения о некоторых локальных химических медиаторах, нейромедиаторах и гормонах - указаны места их синтеза, структура и основное действие. Видно, что структура сигнальных молекул так же разнообразна, как и их функции. Среди этих молекул есть короткие пептиды, более крупные белки и гликопротеины, аминокислоты и родственные им соединения, стероиды (вещества, образующиеся из холестерола и очень сходные между собой по структуре) и производные жирных кислот. Каждая сигнальная молекула представлена в табл. 12-1 только в одном разделе, хотя многие из них могут действовать несколькими способами. Например, некоторые пептидные гормоны в головном мозгу позвоночных действуют как нейромедиаторы (в качестве паракринных факторов).
12.1.3. Разные клетки по-разному реагируют на один и тот же сигнал
Большинство клеток у взрослых животных специализировано для выполнения какой-то одной главной функции, и все они имеют характерный набор рецепторов, который позволяет им реагировать на химические сигналы, запускающие или модулирующие эту функцию. Многие сигнальные молекулы действуют в очень низкой концентрации (обычно не более 10-8 М), и связывающие их рецепторы, как правило, имеют к ним высокое сродство (константа сродства Ка≥ 108 л/моль).
Одни и те же сигнальные молекулы часто оказывают различное действие на разные клетки-мишени. Например, ацетилхолин стимулирует сокращение волокон скелетной мускулатуры, но уменьшает частоту и силу сокращения клеток сердечной мышцы. В данном случае рецепторы ацетилхолина в скелетных мышцах отличаются от рецепторов на клетках миокарда. Но не всегда причина состоит в различии рецепторов. Часто одинаковые сигнальные молекулы связываются с идентичными рецепторами, и все же это ведет к совершенно разным реакциям у различных клеток-мишеней (рис. 12-5). Это означает, что ответы клеток-мишеней могут быть запрограммированы двумя способами: либо самим набором рецепторов клеточной поверхности, либо теми внутриклеточными системами, с которыми эти рецепторы сопряжены.
12.1.4. Реакция клетки на химический сигнал может быть в одних случаях быстрой и кратковременной, в других - медленной и продолжительной [2, 3]
Химические сигналы, приводящие клеточные функции в соответствие с изменениями среды, обычно вызывают быстрые и кратковременные ответы. Например, повышение уровня глюкозы в крови стимулирует секрецию белкового гормона инсулина эндокринными клетками поджелудочной железы. В считанные минуты повышение концентрации инсулина заставляет клетки печени и мышц усиленно поглощать глюкозу, и ее концентрация в крови падает. Этот ответ состоит из трех частей, ни одна их которых не требует синтеза нового белка:
1) в поджелудочной железе повышение уровня глюкозы заставляет клетки освобождать (путем экзоцитоза) запасенный в них инсулин;
2) в жировых и мышечных клетках во внутриклеточных пузырьках имеется некоторый резерв белков, транспортирующих глюкозу, и повышение уровня инсулина стимулирует включение этих пузырьков с их белками в плазматическую мембрану, что повышает скорость поглощения глюкозы; тогда уровень глюкозы в крови падает, и это снижает секрецию инсулина;
3) поскольку при снижении уровня инсулина добавочные переносчики глюкозы быстро удаляются с клеточной поверхности путем эндоцитоза и снова включаются во внутриклеточный пул, скорость поглощения глюкозы жировыми и мышечными клетками возвращается к исходному уровню. Таким способом поддерживается относительно постоянная концентрация глюкозы в крови.
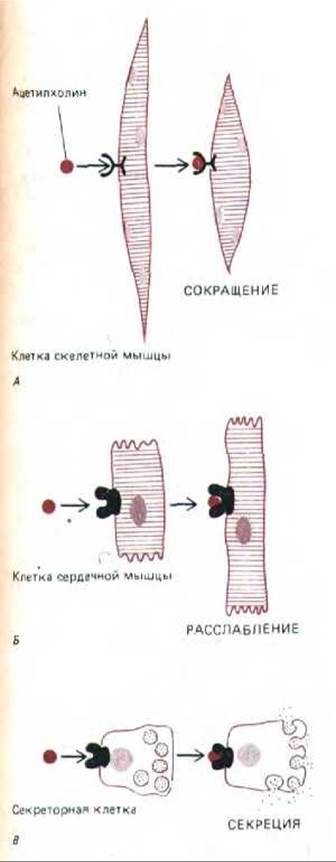
Рис. 12-5. Одни и те же сигнальные молекулы могут вызывать в разных клетках-мишенях различные ответы. В некоторых случаях это обусловлено тем, что сигнальная молекула связывается с разными белками-рецепторами (А и Б). В других случаях такие молекулы связываются с одинаковыми рецепторами, но активируют в разных клетках разные механизмы ответа (В и Г).
Нейромедиаторы вызывают еще более быстрые реакции: в ответ на освобождение ацетилхолина из двигательных нервных окончаний волокна скелетной мышцы сокращаются и вновь расслабляются всего лишь за несколько миллисекунд.
Химические сигналы играют важную роль и в процессах индивидуального развития животных, нередко определяя время и тип дифференцировки тех или иных клеток. Некоторые из вызываемых эффектов проявляются медленно и бывают продолжительными. Например, в период полового созревания клетки яичника начинают секретировать в больших количествах стероидный женский половой гормон эстрадиол. Этот гормон вызывает изменение многих клеток в различных частях организма, что в конце концов приводит к развитию вторичных женских половых признаков, например к увеличению грудных желез. Если секреция эстрадиола прекращается, эти эффекты постепенно исчезают, но некоторые реакции, вызываемые стероидными половыми гормонами на очень ранней стадии развития млекопитающих, необратимы. Сходным образом, десятикратное повышение концентрации тиреоидного гормона в крови головастика стимулирует ряд радикальных и необратимых изменений, приводящих к его превращению в лягушку (рис. 12-6).
12.1.5. Только жирорастворимые сигнальные молекулы могут самостоятельно проникать в клетку
Все известные нейромедиаторы, а также большинство гормонов и локальных химических медиаторов водорастворимы. Есть, однако, исключения, и они образуют отдельный класс сигнальных молекул. Важными примерами служат сравнительно плохо растворимые в воде стероидные и тиреоидные гормоны, которые переносятся кровью в виде растворимых комплексов со специфическими белками-переносчиками. С таким различием в растворимости связаны фундаментальные различия в механизмах действия этих двух классов молекул на клетки-мишени. Водорастворимые молекулы слишком гидрофильны, чтобы прямо приходить через липидный бислой плазматической мембраны; поэтому они связываются со специфическими белковыми рецепторами на клеточной поверхности. Напротив, стероидные и тиреоидные гормоны растворимы в липидах и, отделившись от белка-носителя, могут легко проникать через плазматическую мембрану клетки-мишени. Эти гормоны связываются с белковыми рецепторами внутри клетки (рис. 12-7).
Еще одно важное различие между двумя описанными классами сигнальных молекул - это разная продолжительность жизни в кровотоке или тканевой жидкости. Водорастворимые молекулы, перейдя в кровь, обычно удаляются и/или разрушаются за время, измеряемое минутами, а локальные химические медиаторы и нейромедиаторы после выхода в межклеточное пространство инактивируются еще быстрее - за время порядка секунд или даже миллисекунд. В отличие от этого стероидные гормоны циркулируют в крови часами, а тиреоидные могут сохраняться несколько дней. Поэтому водорастворимые сигнальные молекулы обычно вызывают кратковременные реакции, а водонерастворимые - более продолжительный ответ. Здесь, однако, тоже есть исключения из основного правила: простагландини, например, являются гидрофобными локальными медиаторами, но они связываются с поверхностными клеточными рецепторами и вызывают кратковременный ответ.
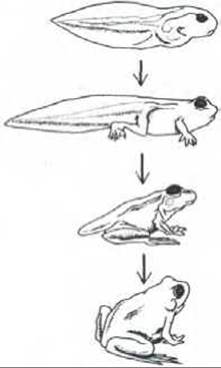
Рис. 12-6. Превращение головастика в лягушку. Показанные радикальные изменения вызываются тиреоидным гормоном. Если у развивающегося эмбриона удалить зачаток щитовидной железы, животное будет продолжать расти как головастик, не претерпевая метаморфоза. При введении тиреоидного гормона этот гигантский головастик превращается в лягушку.
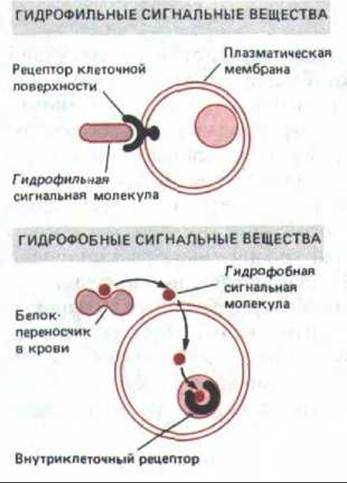
Рис. 12-7. Внеклеточные сигнальные молекулы в зависимости от своей растворимости связываются с поверхностными или внутриклеточными рецепторами. Гидрофильные молекулы не способны прямо проходить через плазматическую мембрану, поэтому они связываются с рецепторами на поверхности клетки-мишени. Многие гидрофобные молекулы могут диффундировать через плазматическую мембрану и связываться с рецепторами внутри клетки. Будучи нерастворимы в водных средах, гидрофобные сигнальные молекулы транспортируются кровью в виде комплексов со специальными белками-переносчиками, от которых они отделяются, перед тем как проникнуть в клетку-мишень.
12-3
12.1.6. Локальные химические медиаторы после их секреции быстро разрушаются, подвергаются обратному захвату или иммобилизуются [4]
Сигнальные молекулы паракринного типа воздействуют только на ближайшее окружение выделяющей их клетки. Такие локальные химические медиаторы столь быстро поглощаются клетками, разрушаются внеклеточными ферментами или иммобилизуются во внеклеточном матриксе, что, как правило, не попадают в кровь в сколько-нибудь значительном количестве.
Некоторые локальные медиаторы вырабатываются специально приспособленными для этого клетками. Например, гистамин (производное аминокислоты гистидина, см. табл. 13-1) выделяют главным образом тучные клетки. Эти клетки, встречающиеся в соединительной ткани всех частей тела, накапливают гистамин в больших секреторных пузырьках и в случае повреждения ткани, при местной инфекции или при некоторых иммунных реакциях быстро освобождают его путем экзоцитоза (разд. 18.2.5). Гистамин вызывает местное расширение кровеносных сосудов и увеличивает их проницаемость, что облегчает доступ к поврежденному участку фагоцитирующим лейкоцитам и белкам сыворотки (например, антителам и компонентам системы комплемента-см. гл. 18). Тучные клетки выделяют также два тетрапептида, привлекающих к месту своей секреции лейкоциты из группы эозинофилов; эозинофилы же содержат разнообразные ферменты, участвующие в инактивации гистамина и других освобождаемых тучными клетками медиаторов, что способствует прекращению реакции.
Некоторые локальные химические медиаторы после их секреции не уничтожаются, а быстро иммобилизуются. В эту группу входят фибронектин, протеогликаны и ряд других макромолекул внеклеточного матрикса. Эти секретируемые клетками макромолекулы можно рассматривать как специальную группу локальных медиаторов, так как они оказывают влияние только на соседние клетки (разд. 14.2). В отличие от других локальных медиаторов они объединяются во внеклеточном пространстве в нерастворимую сеть, утрачивают подвижность и не могут поэтому диффундировать из того места, где образовались. Таким образом, хотя их эффект и локален, он может быть продолжительным. Внеклеточный матрикс тоже иногда связывает растворимые сигнальные молекулы, иммобилизуя их так, что они действуют только в определенном участке. Например, фактор роста фибробластов (fibroblast growth factor, FGF), - небольшой белок, стимулирующий деление самых разнообразных клеток в культуре, - in vitro прочно связывается с протеогликаном внеклеточного матрикса и, возможно, таким же способом иммобилизуется в тканях.
12.1.7. У млекопитающих клетки всех тканей непрерывно выделяют простагландины [5]
Для синтеза многих химических медиаторов существуют специальные клетки, но есть и такие медиаторы, источники которых более разнообразны. Важным примером служат простагландины - семейство производных жирной кислоты с 20 атомами углерода, образующихся во всех тканях млекопитающих. Эти локальные медиаторы непрерывно синтезируются в мембранах клеток из предшественников, отщепляемых от мембранных фосфолипидов фосфолипазами (рис. 12-8), и столь же непрерывно разрушаются ферментами во внеклеточной жидкости. Существует не менее 16 различных простагландинов, подразделяемых на 9 классов (PGA, PGB, PGC, ... PGI), которые, связываясь со специфическими рецепторами клеточной поверхности, вызывают разнообразные биологические эффекты.
В отличие от большинства других сигнальных молекул простагландины не накапливаются в клетках, а по мере их синтеза все время освобождаются во внеклеточное пространство. Однако, когда клетки активируются при повреждении ткани или под действием некоторых химических факторов, скорость синтеза простагландинов возрастает; повышение локальной концентрации простагландинов оказывает воздействие не только на клетку, которая их выделяет (аутокринная стимуляция), но и на соседние клетки. Аутокринная стимуляция, возникающая в ответ на другой химический сигнал, может усиливать эффект последнего и (или) его продолжительность, а также его распространение на большую локальную популяцию однотипных клеток.
С простагландинами связывают множество разнообразных биологических эффектов. Они вызывают сокращение гладкой мускулатуры, агрегацию тромбоцитов, участвуют в воспалительной реакции. Некоторые простагландини, образующиеся в матке во время родов, по-видимому, важны для стимуляции сокращений гладкой мускулатуры матки. Эти простагландины сейчас широко используют в качестве фармакологических агентов, вызывающих аборт. С другой стороны, действие таких противовоспалительных средств, как аспирин, основано на подавлении биосинтеза прост агландинов в очагах воспаления.
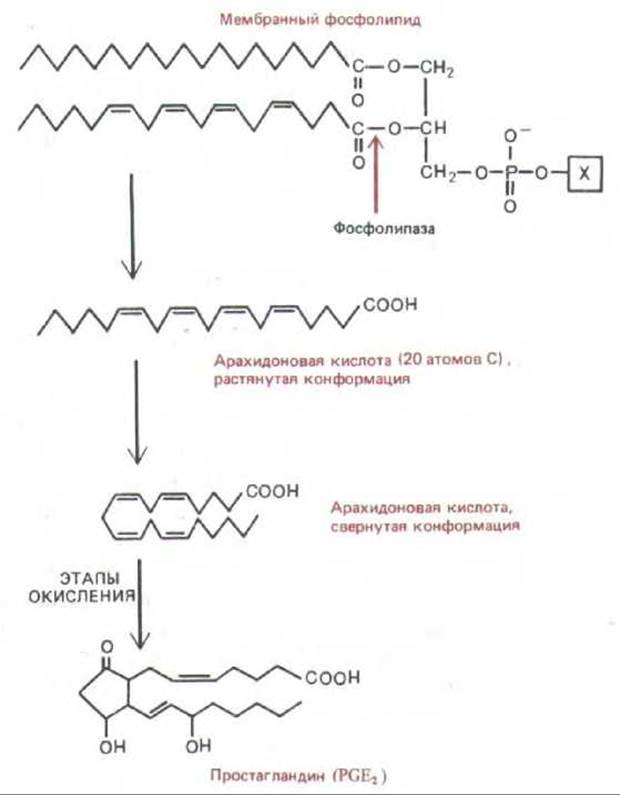
Рис. 12-8. Простагландины непрерывно синтезируются в мембранах из жирных кислот с длинной цепью, состоящих из 20 углеродных атомов и содержащих не менее трех двойных связей. На схеме показан синтез РОЕ2. Индекс указывает на две двойные углерод-углеродные связи вне кольца PGE2. Простагландины вместе с химически родственными им сигнальными молекулами - тромбоксонами, лейкотриенами и липоксинами-образуются главным образом из арахидоновой кислоты и носят общее название эйкозаноидов. Этот метаболический путь служит важной мишенью для лекарственных препаратов, так как эйкозаноиды играют большую роль в развитии воспаления. Кортикостероидные гормоны, такие как кортизон, широко применяются при лечении неинфекционных воспалительных заболеваний, например некоторых форм артрита. Один из возможных механизмов их действия - индукция синтеза и (или) секреции лейкоцитами локальных химических медиаторов, называемых липокортными (или кальпактинами). Это белки, каким-то образом подавляющие активность фосфолипазы на первом этапе показанного здесь пути синтеза эйкозаноидов. Такие нестероидные противовоспалительные препараты, как аспирин, блокируют окислительные этапы синтеза простагландинов. Для лечения артритов применяют как кортикостероиды, так и аспирин.
Заключение
Внеклеточные сигнальные молекулы можно разделить по степени их «дальнодействия» на три основных класса: 1) локальные химические медиаторы, которые быстро поглощаются или разрушаются и поэтому оказывают влияние только на соседние клетки; 2) гормоны, которые переносятся к своим мишеням, распределенным нередко по всему организму, с кровотоком; 3) нейро медиаторы, действующие только на постсинаптическую клетку. Каждый тип клеток организма имеет свойственный ему набор белков-рецепторов, позволяющий запрограммированным и характерным образом реагировать на соответствующий набор сигнальных молекул.
Сигнальные молекулы можно также классифицировать по их растворимости в воде. Небольшие гидрофобные молекулы, такие как стероидные гормоны или гормоны щитовидной железы, свободно проходят через плазматическую мембрану клетки-мишени и активируют рецепторний белок в ее цитоплазме. В отличие от этого гидрофильные молекулы, в частности нейро медиаторы и большинство гормонов и локальных химических медиаторов, активируют белковый рецептор на поверхности клетки-мишени.