Молекулярная биология клетки - Том 3 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
От клеток к многоклеточным организмам
Клеточные механизмы развития
Принципы образования пространственных структур
Различные органы животного возникают не из хаотического скопления клеток; пространственная организация разных типов клеток исходно предопределена. Взаимодействия, координирующие процесс становления пространственной организации, могут быть чрезвычайно разнообразными. В некоторых случаях, например в мозаичных яйцах, пространственная организация может быть заложена в одной клетке, и возникающее внутриклеточное распределение цитоплазматических детерминантов воспроизводится в расположении дочерних клеток эмбриона. В других случаях, например при индукции мезодермы у Xenopus или вульвы у нематод, определенная структура возникает как результат взаимодействия отдельных клеток. Следовательно, в формировании пространственной организации важная роль принадлежит всем видам пространственных сигналов, как внутриклеточных, так и межклеточных. Можно сказать, что эти сигналы обеспечивают клетки позиционной информацией, направляя их дальнейшую специализацию.
В этом разделе мы рассмотрим различные пути создания и использования позиционной информации. Здесь же мы увидим, как в процессе развития многих организмов происходит приобретение отдельными клетками точных молекулярных «адресов» или позиционных значений. Так, клетка раннего эмбриона может «знать» сравнительно немного, например, насколько близко она расположена по отношению к голове или к хвосту. Что же касается ее потомков, существующих в теле взрослого животного, то многие клеточные поколения спустя они могут «знать» о том, что являются костными клетками, расположенными на конце третьего сустава передней конечности. Такая специализация выстраивается постепенно за счет механизмов, основанных на клеточной памяти: ранние пространственные сигналы лишь довольно приблизительно определяют общие координаты, а последующие сигналы местного значения обеспечивают клетки дополнительными уточнениями адреса, привязанными к местным координатам.
16.4.1. Асимметрия, определяемая пространственной организацией среды, может нарастать за счет положительной обратной связи [34, 35]
Формирование пространственной организации начинается с создания асимметрии: будущая голова должна отличаться от хвоста, спина от живота. Мы уже видели, что у животных большинства видов яйцеклетки асимметричны (см. разд. 16.2.2). Асимметрия зачастую возникает в яйцеклетке в процессе ее развития в яичнике. У насекомых, например, ооцит (будущая яйцеклетка) расположен в одном из концов фолликула, где одной стороной он соприкасается с питающими клетками, передающими ему запасаемые вещества по цитоплазматическим мостикам (см. разд. 15.3.4). По всей вероятности, асимметрическое поступление запасаемых веществ в ооцит создает химические различия между разными участками цитоплазмы яйца, которые соответствуют в будущем голове и хвосту эмбриона (см. рис. 16-39).
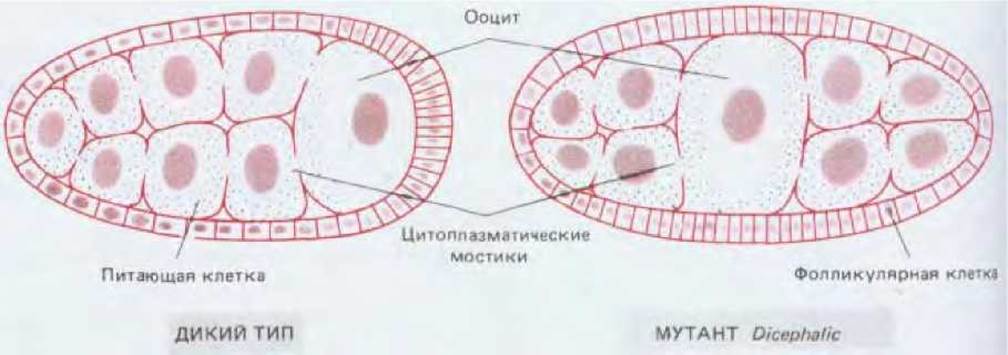
Рис. 16-39. Контроль полярности яйца дрозофилы со стороны среды, окружающей ооцит в яичнике. В норме питающие клетки поставляют запасаемые вещества на один из полюсов ооцита, который соответствует будущей голове. У мутантов Dicephalic ооцит расположен в центре фолликула и симметричен по отношению к питающим клеткам. Вследствие этого возникает симметричное яйцо, дающее начало эмбриону с головами на обоих концах.
Исходно асимметрия может быть не слишком выражена: далее клетки воспользуются своими собственными внутренними механизмами для усиления слабой асимметрии и превратятся в поляризованные. Например, у яйцеклетки морской водоросли Fucus ризоид, или корешок, образуется только на одном конце. В норме ризоид растет вниз и от света. Таким образом, сила тяготения и освещенность вместе определяют, на какой стороне он образуется. Такая полярность обусловлена потоком ионов Са2+ через клетку (см. разд. 20.5.6): полагают, что случайный свет или сила тяготения вызывают появление небольшой асимметрии в распределении белков, участвующих в транспорте ионов Са2+ через плазматическую мембрану. Эта асимметрия позже усиливается за счет механизмов положительной обратной связи (см. рис. 20-66, разд. 20.5.7).
Таким же образом положительная обратная связь может действовать и на других стадиях развития многоклеточных организмов. Например, при делении поляризованной клетки дочерние клетки наследуют слабо выраженную асимметрию и, усилив ее, становятся более поляризованными. Так происходит усиление поляризации в ряду клеточных поколений. Поэтому в принципе для возникновения сложной структуры многоклеточных организмов межклеточный обмен сигналами не обязателен (рис. 16-40). Возможно, что некоторые процессы формирования пространственной организации, зависящие от автономного поведения клетки, можно объяснить действием такого механизма.
Однако, как правило, формирование пространственной организации у многоклеточных организмов зависит и от межклеточных сигналов. И здесь положительная обратная связь также выполняет важную роль усиления исходно слабых асимметрий, создавая локализованные группы клеток, свойства которых очень различны. Такие группы клеток могут в свою очередь испускать сигналы, влияющие на развитие соседних клеток. Простейшим примером может служить примитивный организм Hydra. Тело гидры имеет форму трубки, на одном конце которой расположена «голова», снабженная щупальцами, и на другом конце - «нога» (см. рис. 1-33). Гидра известна способностью восстанавливать целостность организма после удаления какой-либо из его частей. Множество экспериментов по ампутации «головы», выполненных на гидре, позволяют предположить, что процесс регенерации осуществляется благодаря локальному аутокаталитическому процессу, усиливающему слабую тенденцию клеток к приобретению свойств клеток «головы». В создании данной утраченной структуры принимают участие все клетки тела; они создают ингибирующий сигнал, диффундирующий вдоль продольной оси тела, и исключающий условия формирования «головы» в иных участках организма (рис. 16-41).
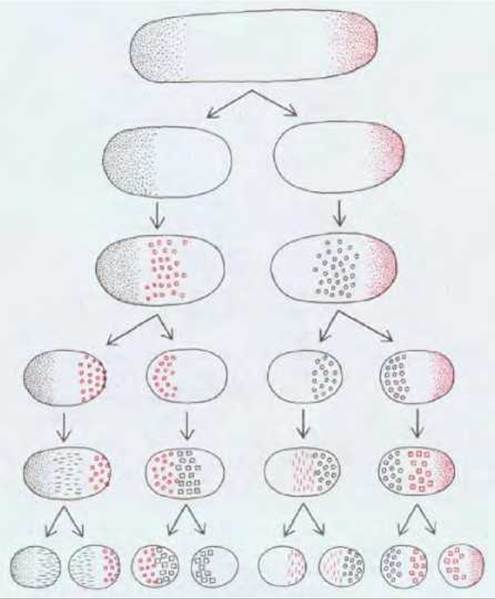
Рис. 16-40. Сложная многоклеточная структура в принципе может возникнуть в результате серии асимметричных клеточных делений без каких-либо межклеточных взаимодействий (помимо обеспечивающих соединение клеток). Внутренней полярности, наследуемой каждой из дочерних клеток в результате последовательных клеточных делений, может оказаться вполне достаточно для управления следующим циклом внутриклеточного структурообразования. Возникающие при последующих делениях дочерние клетки будут опять-таки весьма различными и внутренняя поляризация этих клеток не будет уменьшаться.
16.4.2. Локализованный участок, испускающий сигнал, может часто создавать градиент морфогена [36]
Влияние головы на остальную часть тела гидры, которое характеризуется значительным радиусом действия, представляет собой весьма распространенное явление. Во многих развивающихся системах небольшие участки ткани способны приобретать какие-либо особые свойства, превращающие их в источник сигнала, который распространяется через прилежащие участки ткани, и может контролировать их поведение. В частности, сигналом могут служить диффундирующие молекулы, секретируемые сигнализирующим участком. Предположим, что по мере диффузии сигнального вещества через соседние ткани происходит его разрушение. В этом случае наивысшая концентрация сигнального вещества будет вблизи источника. С увеличением расстояния от источника количество этого вещества будет уменьшаться, вследствие чего в ткани возникнет концентрационный градиент (см. рис. 16-42). На разных расстояниях от источника сигнала клетки будут подвергаться различному действию этого вещества в зависимости от его концентрации и благодаря этому приобретут разные свойства. Гипотетическое вещество, подобное вышеупомянутому, концентрация которого прочитывается клетками и позволяет последним определить расстояние активатора в этом участке вызывает образование ингибитора, диффундирующего на значительные расстояния и подавляющего синтез активатора в других участках. Например, при декапитации гидры оставшаяся ткань в норме регенерирует голову, но если на место ампутированной головы или на некотором расстоянии от него пересадить голову другой гидры, то новой дополнительной головы не образуется.
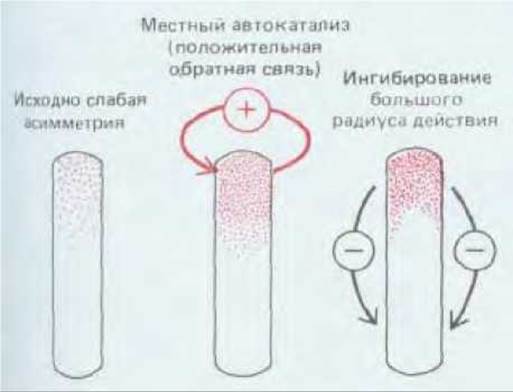
Рис. 16-41. Механизм создания пространственной структуры вследствие автокаталитического процесса.
Вещество «активатор» в результате автокаталитического процесса накапливается до весьма высокого уровня в небольшом участке, расположение которого определяется слабовыраженной исходной асимметрией системы; высокая концентрация относительно неких ориентиров или «маяков», называется морфогеном. Сигнализирующий участок способен с помощью морфогена контролировать формирование пространственной организации больших полей прилежащей ткани. Градиенты морфогена представляют собой простое и эффективное средство обеспечения клеток позиционной информацией. Именно поэтому морфогены и оказались в центре нашего внимания. Здесь необходимо добавить, что клетки эмбриона способны приобретать позиционную информацию иными способами. Рис. 16-21 иллюстрирует одну из таких альтернатив: здесь в основе механизма лежат индукционные взаимодействия короткого радиуса действия.
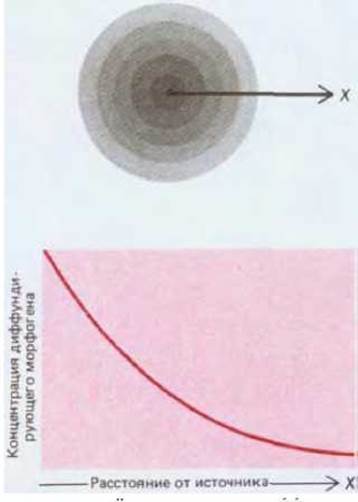
Рис. 16-42. Если какое-то вещество образуется в определенной точке и по мере диффузии из этой точки разрушается, то создается градиент концентрации этого вещества с максимумом в исходной точке. Такое вещество может служить морфогеном, локальная концентрация которого контролирует поведение клеток в соответствии с их расстоянием от источника вещества.
И хотя известно множество примеров сигнализирующих участков, которые (с нашей точки зрения) через градиент морфогена могли бы оказывать воздействие на значительном расстоянии, на практике в настоящее время мы имеем дело всего лишь с несколькими случаями непосредственной химической идентификации морфогена. Как правило, известно лишь, что при изменении местоположения предполагаемого сигнализирующего участка после пересадки или иным способом, происходит изменение структуры прилежащих участков ткани. Наблюдения такого рода не позволяют выяснить, насколько изменения структуры отдельных клеток определяются прямым ответом на действие морфогена и каков вклад от взаимодействия реагирующих клеток. Вероятно, в большинстве случаев исходный градиент морфогена вызывает импульс, обеспечивающий формирование пространственной организации, широкого спектра действия, а локальные межклеточные взаимодействия участвуют в проработке деталей. Накапливаются данные, свидетельствующие о том, что именно таким образом возникает пространственная организация тела насекомых; это будет обсуждаться ниже. Здесь же мы рассмотрим, какой тип пространственной организации может быть обусловлен действием градиента морфогена в поле клеток, каждая из которых реагирует назависимо.
16.4.3. Порог реакции клетки обусловливает строго определенный характер ее детерминации, несмотря на плавный градиент морфогена [37]
При наличии плавного градиента концентрации морфогена можно ожидать, что и свойства клеток в разных участках будут изменяться постепенно. Такие слабо выраженные различия действительно встречаются в некоторых тканях. Но наибольший интерес вызывает возникновение резких качественных различий - таких, например, как различия между хрящевыми и мышечными клетками, не имеющими переходных форм. В популяции исходно однородных клеток благодаря порогу реакции на плавно изменяющийся сигнал могут возникать резкие различия между клетками: в каждой из реагирующих клеток эффект небольшого приращения сигнала может быть усилен по принципу положительной обратной связи так, что клетки, подвергающиеся действию сигнала, интенсивность которого изменяется слабо, выберут различные пути развития в зависимости от того, подверглись ли они действию сигнала над- или подпороговой интенсивности. При этом для каждого из сигналов могут существовать несколько порогов интенсивности, и одна переменная может контролировать несколько выборов. Если под влиянием какого-то фактора клетка определенно вступила на путь, ведущий к одному из стабильных состояний, то она будет развиваться в выбранном направлении даже в отсутствие фактора, исходно контролировавшего выбор. На этом пути временные, зависящие от положения клетки воздействия могут вызывать эффект «запоминания» воздействий, которые претерпела клетка. Выбор клеткой определенного состояния зависит от того, какая позиционная информация сохранилась в ее клеточной памяти. Такую запись, воплощенную в виде определенного свойства клетки, можно назвать ее «позиционным значением».
16.4.4. Эмбриональные поля очень малы, поэтому основные черты строения взрослого животного должны детерминироваться достаточно рано с участием клеточной памяти [38]
Независимо от природы механизмов, обеспечивающих клетки позиционной информацией, зона их действия довольно ограниченна: они обычно эффективны лишь в пределах небольших областей (морфогенетических полей), длина которых составляет около 1 мм, что соответствует примерно 100 или менее клеточным диаметрам. Отсюда следует, что существует предел количества деталей, которые могут быть заложены на столь ограниченном пространстве. Именно поэтому конечное позиционное значение клетки приходится фиксировать в виде последовательных элементов позиционной информации, записываемых на разных стадиях развития. В связи с этим механизм детерминации, основанный на клеточной памяти, совершенно необходим для развития крупных, сложно устроенных животных. Различие между головой и хвостом должно закладываться еще тогда, когда длина соответствующих зачатков не превышает 1 мм. К тому времени, когда длина животного достигнет сантиметра или метра, события, в результате которых возникли эти различия, будут уже «древней историей»; и для того, чтобы данные различия сохранялись, клетки должны обладать хорошей памятью.
Таким образом, общий план строения тела определяется очень рано, а детали, все более и более тонкие, добавляются позже, по мере того, как зачатки отдельных органов достигают размеров, подходящих для записи дополнительной позиционной информации (рис. 16-43).
Как это происходит, мы рассмотрим более подробно на примере развития конечности.
16.4.5. В процессе развития конечности позиционная информация постепенно уточняется [39]
Развитие конечностей и многих других органов (например, зубов, позвонков или кожи) обеспечивается весьма ограниченным числом различных способов дифференцировки. Конечность, например, образована клетками нескольких типов, а именно мышечными, хрящевыми, костными и клетками соединительной ткани. Но эти немногочисленные типы дифференцированных клеток организованы в пространстве весьма сложным образом. При этом различия между передней и задней конечностями не связаны с появлением дополнительных типов тканей; каждая конечность характеризуется иным способом расположения тканей в пространстве. Эксперименты по трансплантации развивающихся конечностей у цыпленка показали, что внутренние различия клеток конечностей, связанные с возникающими структурами, определяются задолго до начала дифференцировки.
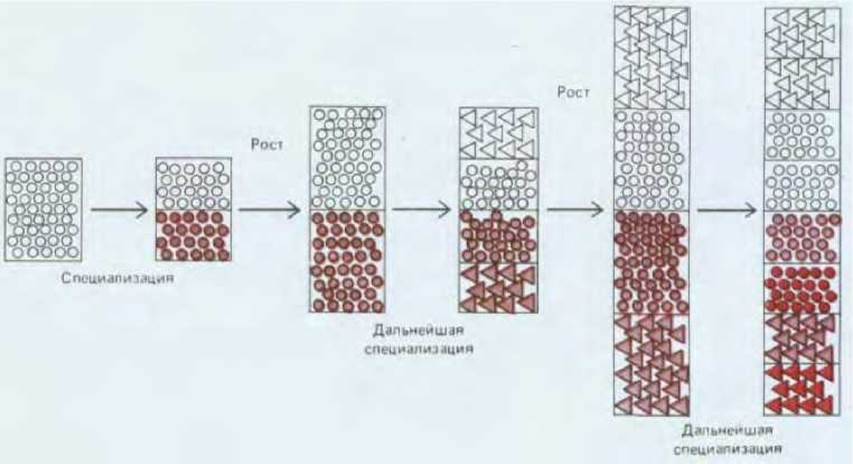
Рис. 16-43. Когда эмбрион мал, он начинает подразделяться на несколько отличных друг от друга по своим свойствам областей, соответствующих основным частям тела взрослого организма. Клетки каждого такого участка, границы которого установить довольно трудно, приобретают достаточно широкие позиционные значения (представленные здесь в виде треугольников или кружочков). По, мере роста эмбриона эти области также растут и в свою очередь в них начинают обособляться подобласти, в результате чего возникает все более детализованная картина позиционных значений.
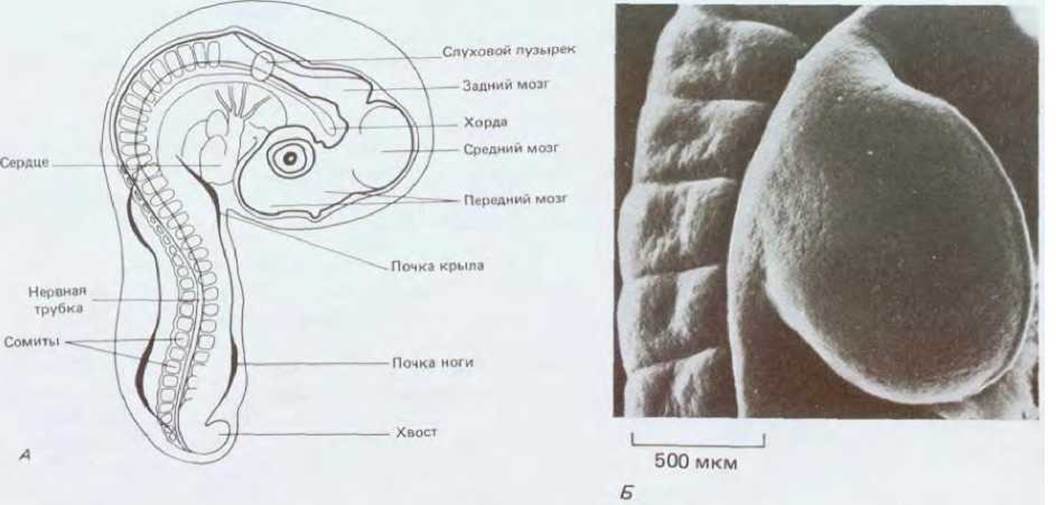
Рис. 16-44. А. Куриный эмбрион после трех дней инкубации. Показано расположение ранних зачатков (почек) конечностей. Б. Зачаток крыла и прилежащие к нему сомиты после еще одного дня развития; зачаток растет, превращаясь в язычкообразный вырост (примерно I мм в длину, 1 мм в ширину и 0,5 мм в толщину). Микрофотография дорсальной стороны крыла, полученная с помощью сканирующего электронного микроскопа. (А - по W. H. Freeman, В. Bracegirdle. An Atlas of Embryology. London, Heinemann, 1967; Б-с любезного разрешения Paul Martin.)
Ноги и крылья у куриного эмбриона возникают почти одновременно в виде небольших вытянутых зачатков, выступающих на боковых поверхностях зародыша (рис. 16-44). Клетки двух пар зачатков конечностей внешне не различаются и в это время не дифференцированы; ни в чем не проявляется также и характер будущего скелета (рис. 14-20, разд. 14.2). На этой стадии из основания зачатка ноги можно вырезать небольшой участок недифференцированной ткани (в норме этот участок образует голень) и пересадить его на верхушку зачатка крыла. В процессе развития трансплантата формируется не голень и не соответствующая часть кончика крыла, а палец (рис. 16-45). Из этого опыта следует, во-первых, что клетки раннего зачатка ноги детерминированы в качестве клеток ноги, и во-вторых, что, будучи детерминированы в этом качестве, они еще не восприняли детализованное позиционное значение вдоль оси зачатка и могут отвечать на информационные сигналы крыла формированием структур, соответствующих не основанию конечности, а ее концу. Можно заключить, что окончательная дифференцировка клеток у позвоночных детерминируется серией последовательных сигналов позиционной информации, регистрируемых клеточной памятью в разное время. Конечное состояние клетки возникает как результат определенной последовательности решений.
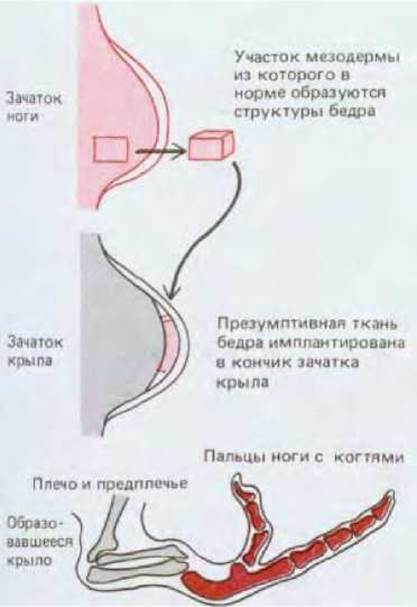
Рис. 16-45. Ткань будущего бедра после пересадки на край почки крыла (у куриного эмбриона) образует пальцы. (J. W. Saunders et al., Dev. Biol., 1, 281-301, 1959.)
16.4.6. После приобретения позиционных значений сходные клетки становятся неэквивалентными [40]
Хотя зачатки передней и задней конечности образованы дифференцированными клетками одинаковых типов, совершенно очевидно, что эти клетки неэквивалентны. Они различаются по своему внутреннему состоянию, определяемому разными позиционными значениями. Клетки зачатков конечностей способны сохранять позиционное значение, отличающее ногу от крыла, даже после дифференцировки; и если это так, то дифференцированные клетки одного и того же типа (например, хрящевые) будут неэквивалентны. Отсюда следует, что на самом деле клетки взрослого организма значительно более разнообразны, чем это учитывает традиционная классификация клеток позвоночных (см. гл. 17, дополнение).
В распоряжении исследователей имеются данные, свидетельствующие в пользу существования таких слабых различий между дифференцированными клетками одного и того же типа, расположенными в разных участках тела позвоночных. Примеры неэквивалентности клеток кожи и нервной системы будут рассмотрены в этой главе ниже (разд. 16.6.4) и в гл. 19 (разд. 19.7.9). Один из наиболее ярких примеров получен при изучении регенерации конечностей у хвостатых амфибий (Urodeles); в этих исследованиях было показано, что неэквивалентность характерна не только для клеток передних и задних конечностей, но и для клеток, расположенных по оси конечности на разных уровнях.
Если у тритона или аксолотля удалить конечность на любом расстоянии от ее основания, то утраченная часть восстанавливается. На конце культи образуется бугорок из внешне недифференцированных клеток, покрытый эпидермисом; это - так называемая регенерационная бластема. В результате роста и дифференцировки бластемы из нее образуются именно те части конечности, которые должны быть расположены дистально от места ампутации. Если удалена кисть, то образуется кисть, если удалены предплечья и кисть, то - недостающие предплечье и кисть. В данном случае не имеет значения, где развивается бластема: как обычно - на проксимальном конце культи, или (в условиях эксперимента) на конце дистального участка ампутированной конечности (рис. 16-46). В любом случае из клеток бластемы возникают только те части, которые в норме расположены дистально по отношению к уровню разреза; при этом во втором случае возникает зеркальный дубликат имеющегося участка. Вероятно, клетки, расположенные вдоль оси конечности на разных уровнях, запомнили различные позиционные значения и именно этими значениями они руководствуются в выполнении программы, восстанавливающей утраченные части конечности.
16.4.7. Клетки различных областей обеспечиваются одинаковой позиционной информацией, но интерпретируют ее по-разному [36, 41]
Эксперимент, схематически представленный на рис. 16-45, позволяет приблизиться к пониманию процесса возникновения сложной позиционной детализированной информации в клетках конечности цыпленка. Из этого эксперимента следует, что у птиц сигналы, обеспечивающие клетки информацией о положении вдоль оси конечности, в крыле и ноге, принципиально не различаются. Клетки будущей ноги после трансплантации на кончик зачатка крыла точно прочитывают сообщения, указывающие на их дистальное положение, и поэтому участвуют в формировании пальцев. Но Интерпретируя эту информацию по-своему, они вместо пальцев передней конечности образуют пальцы задней конечности. Механизмы, основанные на явлении клеточной памяти, могут быть использованы для обеспечения позиционной информацией клеток различных участков, или полей. Поэтому в каждом из полей возникнет определенная структура, как результат сочетания информации, закодированной в различной предыстории клеток, и информации определенного межклеточного сигнала.
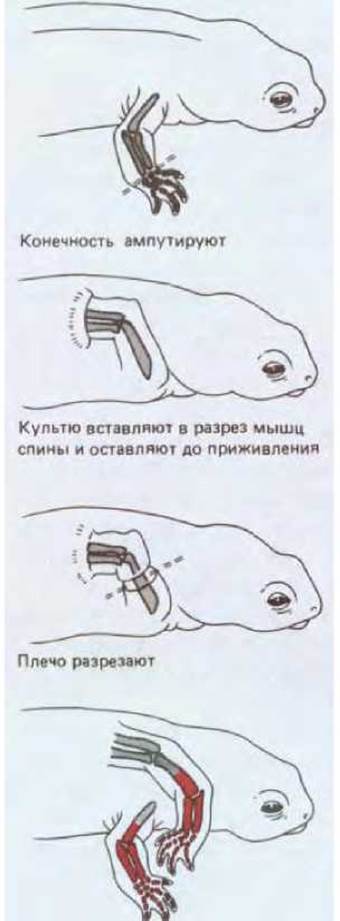
На обеих поверхностях образуется регенерационная бластема; из клеток бластемы в обоих случаях формируются те части конечности, которые в норме расположены дистально от места разреза
Рис. 16-46. Как показывают результаты эксперимента, схема которого приведена на этом рисунке, характер структур (в данном случае у саламандры), образующихся при регенерации бластемы, определяется тем, на каком уровне была отсечена конечность, а не тем, какие из структур в ней сохранились. Предплечье и кисть образуются как из дистального, так и из проксимального отдела исходной конечности.
Эти замечания относятся не только к сигналам, кодирующим формирование пространственной организации вдоль длинной (проксимодистальной) оси конечности, но по оси от большого пальца к мизинцу (переднезадней оси). Две этих оси (рис. 16-47) соответствуют различным компонентам позиционных значений, подобно декартовой системе координат, которые в свою очередь определяются за счет различных способов детализации позиционной информации. Регуляция формирования переднезадней оси имеет особое значение, поскольку это один из немногих примеров, где удалось химически идентифицировать морфоген.
6.4.8 Ретиноевая кислота - вероятный морфоген в зачатке конечности позвоночных [42]
На заднем крае зачатка конечности куриного эмбриона расположена группа клеток, обладающих уникальными свойствами, которые выявляются при пересадке этих клеток в переднюю область зачатка другой конечности (рис. 16-48). Под влиянием трансплантата зачаток-реципиент уже в первые сутки начинает разрастаться вширь и в результате формирует конечность, строение которой очень сильно изменено. В данном случае происходит удвоение всех частей скелета и всех соответствующих этим частям мягких тканей; формируется второй набор элементов, организованный по принципу зеркальной симметрии по отношению к срединной плоскости конечности. Второй набор элементов конечности формируется не из тканей трансплантата, а практически полностью за счет клеток зачатка-реципиента. Можно предположить, что образование второго набора элементов конечности хозяина, расположенных в обратной последовательности, определяется клетками трансплантата. Область зачатка конечности, послужившая источником трансплантата, получила название зоны поляризующей активности или поляризующей области.
Поляризующая область присутствует и в зачатке ноги, и в зачатке крыла; необходимо отметить, что поляризующие области двух пар конечностей и даже конечностей разных видов функционально взаимозаменяемы. Независимо от происхождения трансплантата (нога или крыло, куриный это зародыш или зародыш мыши) он во всех случаях приводит к образованию в крыле цыпленка дополнительного зачатка крыла, возникающего из тканей куриного зародыша, и из этого зачатка крыла формируются дополнительные «пальцы» цыпленка. Следовательно, сигнал, испускаемый поляризующими областями разных конечностей у разных видов, один и тот же. Клетки конечности интерпретируют сигнал согласно своему геному и предыстории.
В нормальном крыле три «пальца» соответствуют трем средним пальцам пятипалой руки и могут быть обозначены цифрами 2, 3, 4. Палец 4 образуется вблизи поляризующей области, палец 3-несколько дальше, а палец 2-еще дальше. Такое же правило справедливо в отношении тканей, расположенных в непосредственной близости к трансплантату поляризующей области, как если бы характер формирующихся пальцев соответствовал характеру некоего морфогена, секретируемого поляризующей тканью (рис. 16-49). Градуальный характер сигнала, исходящего от клеток поляризующей области, можно продемонстрировать, пересаживая меньшее число клеток. При пересадке примерно 30 клеток образуется дополнительный палец 2, при пересадке 80 клеток - дополнительный палец 3 и при пересадке 130 клеток - дополнительный палец 4.
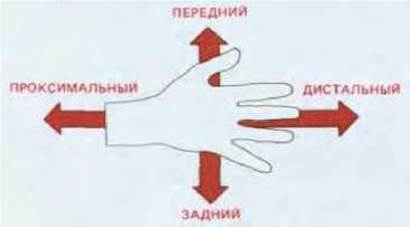
Рис. 16-47. Проксимодистальная и переднезадняя оси конечности позвоночных.
Действие пересаженных клеток поляризующей области можно воспроизвести путем имплантации инертного носителя, пропитанного ретиноевой кислотой. Ретиноевая кислота диффундирует из носителя, и при этом вдоль зачатка конечности создается градиент концентрации, высшая точка которого по крайней мере в три раза превышает минимальную концентрацию; дополнительные пальцы начинают возникать при концентрации около 20 нМ. Благодаря применению высокочувствительных методов хроматографии удалось показать, что в нормальном зачатке конечности существует естественный градиент ретиноевой кислоты именно такого порядка, причем высшая концентрация характерна для клеток поляризующей области. Весьма вероятно, что природным морфогеном является именно ретиноевая кислота (рис. 16-50).
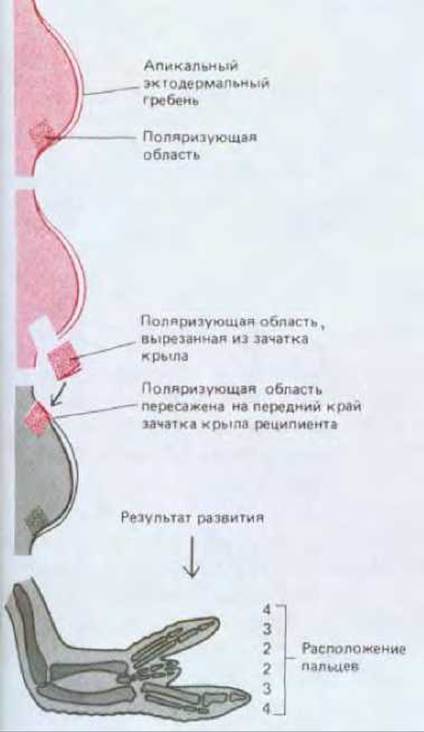
Рис. 16-48. Трансплантат из зоны поляризующей активности вызывает зеркальное удвоение структур крыла у реципиента.
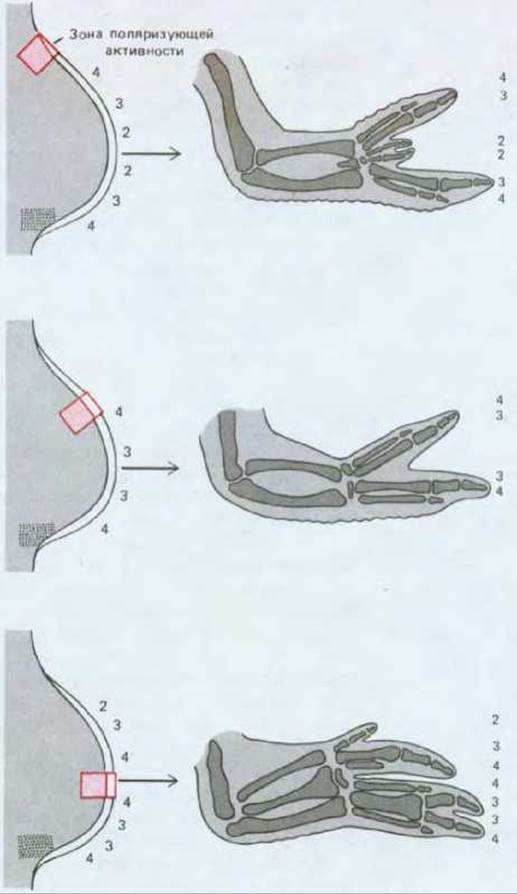
Рис. 16-49. Расстояние между трансплантированным участком зоны поляризующей активности и участком такой же зоны хозяина определяет тип структур, формируемых клетками зачатка крыла. Числа 2, 3 и 4 показывают, какой палец развивается из каждой части зачатка.
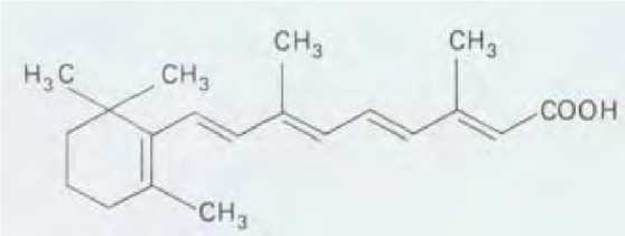
Рис. 16-50. Химическая структура ретиноевой кислоты.
Недавно удалось идентифицировать рецептор ретиноевой кислоты. Им оказался белок, гомологичный рецепторам стероидных и тиреоидных гормонов; он связывается с определенными последовательностями ДНК и регулирует транскрипцию определенных генов. Была выделена и секвенирована кДНК, кодирующая рецептор. В настоящее время для выявления рецепторов в тканях, подобных зачаткам конечностей, используются как ДНК-зонды, так и соответствующие антитела.
16.4.9. Процесс роста контролируется характером позиционных значений, которые могут изменяться при интеркаляции [43]
До сих пор при обсуждении механизмов формирования пространственной организации и позиционных значений мы пренебрегали одним важным аспектом этого процесса: мы не обсуждали регуляцию роста, которая исключительно важна для приобретения различными частями и органами зародыша соответствующих размеров. В некоторых случаях эти механизмы основаны на автономных клеточных программах, запускаемых на ранних стадиях закладки зачатков определенных органов. Однако во многих других случаях рост и структура позиционных значений определяются как результат постоянных межклеточных взаимодействий и тесно связаны с ними. Изучая процессы регуляции, происходящие у многих организмов при замене фрагментов ткани, обладающих различными позиционными значениями, исследователи обнаружили, что клетки этих фрагментов растут и приспосабливаются к локальным условиям. Из этих экспериментов следует простое правило общего характера. Его лучшей иллюстрацией являются результаты исследований по регенерации конечностей у тараканов.
Тараканы относятся к насекомым, у которых в процессе развития от личинки до взрослой формы не наблюдается радикального метаморфоза; развитие протекает постепенно путем смены нескольких ювенильных форм. Конечности молодых тараканов хорошо дифференцированы, но в отличие от клеток конечностей человека здесь дифференцированные клетки способны реагировать на сигналы, управляющие формированием пространственной организации конечности. При нарушении такой организации происходит ее восстановление. Благодаря этим особенностям объекта можно исследовать деятельность систем, контролирующих становление пространственной организации. Для этого по завершении процесса эмбрионального развития на молодых особях проводят необходимые операции.
Мы опишем эксперименты, в которых изучалось поведение клеток эпидермального слоя и кутикулы, покрывающей тело таракана и образующей наружную поверхность конечности. Рост наружного покрова связан с последовательными линьками, когда молодая особь сбрасывает старую кутикулу и вместо нее возникает новая, более просторная. Материал для построения кутикулы выделяют лежащие под ней клетки эпидермиса, образующие один клеточный слой. Позиционные значения этих клеток определяют структуру образуемой ими кутикулы, и если эти значения в результате экспериментального воздействия изменяются, то такой результат можно выявить, исследовав кутикулу после линьки. Регенерация наблюдается только у молодых особей, так как взрослые насекомые не растут и не линяют.
Нога таракана состоит из нескольких частей, расположенных (от основания к концу) в следующем порядке: тазик, вертлуг, бедро, голень, лапка. Лапка в свою очередь состоит из нескольких мелких члеников и оканчивается парой коготков (рис. 16-51). Если удалить две ноги, перерезанные в голени на разных уровнях, то можно трансплантировать дистальную часть одной ноги на проксимальную культю другой. В результате получается «составная» нога без средней части голени. Тем не менее после линьки насекомого образуется внешне нормальная конечность: утраченная средняя часть голени регенерирует (рис. 16-52, А). Операцию можно видоизменить, так что ее результат будет еще более удивительным. Голень одной ноги перерезают около проксимального конца, а голень другой - около дистального. Длинную отрезанную голень одной ноги присоединяют к длинной культе другой, в результате чего образуется удлиненная конечность с удвоенной средней частью (рис. 16-52, Б). После линьки оказывается, что эта нога удлинилась еще больше: между двумя имевшимися срединными частями голени образовалась третья срединная часть. Как показано на рис. 16-52, Б, щетинки на этом вновь образовавшемся участке направлены противоположно по отношению к щетинкам на остальных частях голени.
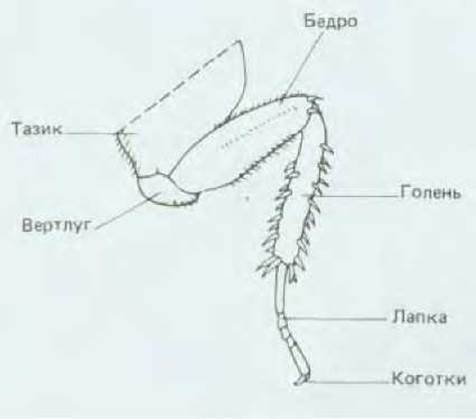
Рис. 16-51. Нога таракана. При каждой из последовательных линек нога растет, но ее строение не меняется.
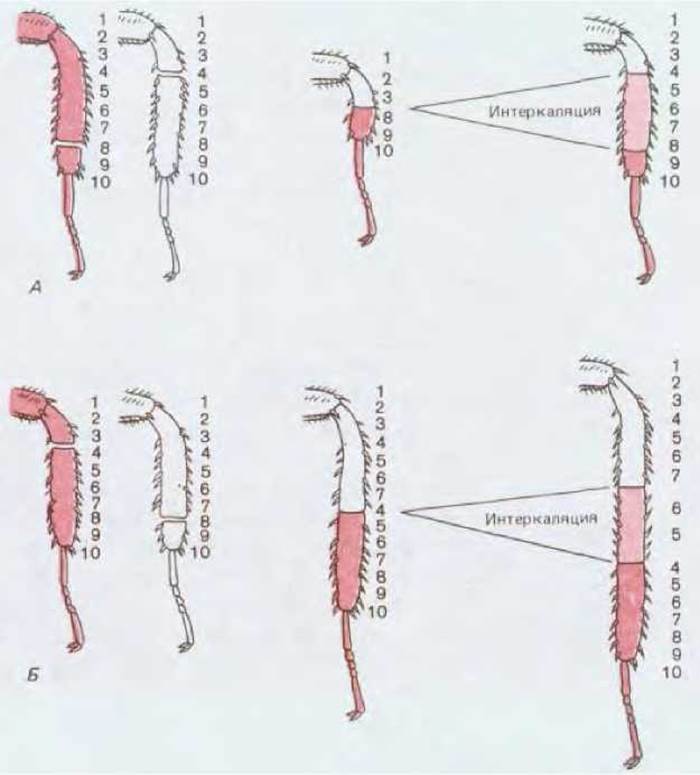
Рис. 16-52. После соединения несовпадающих частей голени таракана происходит интеркаляция новой ткани (показана светлой краской), заполняющей разрыв в ряду позиционных значений (числа от 1 до 10). В первом случае (А) в результате интеркаляции восстанавливается недостающий участок, а во втором (Б) образуется третья, средняя часть голени между двумя уже имеющимися. Направление щетинок указывает на полярность интеркалирующей ткани. В обоих случаях восстанавливается непрерывность ряда позиционных значений.
Было проведено много операций такого типа. Все они указывают на существование системы позиционных значений, наличие которых делает клетки, расположенные в различных участках, неэквивалентными. Характеристика позиционного значения тесно связана с контролем пролиферации клеток. Принято определять позиционное значение числом, изменяющимся от максимума на одном конце участка конечности до минимума на другом. При описанных выше операциях рядом оказывались эпидермальные клетки, резко различающиеся по своим позиционным значениям. Поэтому в месте соединения двух отрезков начиналась пролиферация клеток и новые клетки при этом приобретали значения, которые плавно, без скачков, заполняли разрыв между позиционными значениями клеток, сближенных при операции (рис. 16-52). Такой результат можно обобщить, сформулировав правило интеркаляции: разрывы в плавном ряду позиционных значений вызывают местную пролиферацию клеток и вновь образующиеся клетки приобретают промежуточные позиционные значения, восстанавливая таким образом непрерывность структуры. Пролиферация прекращается только после заполнения промежутка клетками, обладающими всеми утраченными позиционными значениями. При этом восстанавливается нормальное распределение клеток в пространстве. Весь этот процесс получил название интеркалярной (или вставочной) регенерации.
Правило интеркаляции и вытекающее из него следствие, согласно которому рост продолжается до приобретения определенного распределения позиционных значений, - это важный организующий принцип для систем, где он действует. Начиная со структуры, определяемой приблизительно и в миниатюрном масштабе (например, за счет градиента морфогена), это правило предопределяет создание полного набора позиционных значений и регулирует рост каждой из частей данной структуры до определенных размеров. Все это необходимо для того, чтобы возникающая структура качественно, а следовательно, топологически, была верна. Если применить правило интеркаляции к двух- и трехмерным структурам, то оно окажется справедливым для широкого спектра явлений, включая как нормальную регенерацию ампутированных частей тела, так и аномальные эффекты (возникновение дополнительных конечностей при некоторых трансплантациях). По всей вероятности, это правило справедливо по отношению ко многим процессам органогенеза и регенерации не только у насекомых, но и у ракообразных и амфибий. Например, у дрозофилы в соответствии с этим правилом происходит коррекция ранних ошибок спецификации структуры (рис. 16-60). Даже у млекопитающих, утраченные структуры которых во взрослом состоянии не регенерируют, правилу интеркаляции может подчиняться регуляция роста и пространственной организации в эмбриональный период. Молекулярные механизмы, лежащие в основе этого важного способа контроля роста, пока неизвестны.
Заключение
Возникновение различных типов клеток эмбриона строго упорядочено в пространстве. Этот процесс обычно начинается с возникновения асимметрии яйцеклетки и продолжается благодаря межклеточным взаимодействиям в эмбриогенезе. На каждом из этапов этого процесса асимметрия, изначально выраженная очень слабо, усиливается положительной обратной связью и приводит к появлению четко выраженной пространственной организации. Можно сказать, что сигналы, координирующие формирование пространственной организации, снабжают клетки позиционной информацией. В простейшем случае градуальная концентрация диффундирующего морфогена может контролировать свойства клеток в зависимости от их расстояния по отношению к источнику морфогена; вероятно, при развитии конечности у куриного зародыша роль морфогена выполняет ретиноевая кислота, контролирующая возникновение элементов по оси «большой палец-мизинец». Дискретные различия клеток по своим свойствам могут соответствовать пороговым значениям реакции на морфоген.
Пространственная характеристика клетки определяется позиционной информацией, поступившей к ней в разное время. Пока эмбрион мал, эта характеристика весьма приблизительна, однако по мере роста эмбриона к ней добавляются уточняющие детали. Клетки ранних зачатков передней и задней конечностей эмбрионов позвоночных приобретают различные позиционные значения, вследствие чего свойства клеток разных конечностей становятся неэквивалентными задолго до дифференцировки. Уточнения структуры каждого органа возникают после появления в каждом из зачатков этих органов более детализированной внутренней сети позиционной информации. Вероятно, эта система детализированной позиционной информации в гомологичных органах, каковыми являются передние и задние конечности, может быть весьма сходной. Обладая памятью, клетки различных полей интерпретируют уточняющую позиционную информацию по-разному, в соответствии со своей предысторией.
Структура позиционных значений у многих животных тесно связана с контролем клеточной пролиферации по простому правилу интеркаляции. Это правило было сформулировано при изучении регенерации конечностей у насекомых и амфибий. Оно гласит, что нарушение непрерывности позиционных значений вызывает местную пролиферацию клеток, и вновь возникающие клетки приобретают промежуточные позиционные значения, восстанавливающие непрерывность нарушенной структуры. Этот же механизм может функционировать при нормальном развитии зародыша, восстанавливая неточности исходных характеристик позиционной информации.