Молекулярная биология клетки - Том 3 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
От клеток к многоклеточным организмам
Половые клетки и оплодотворение
Преимущества полового процесса
Цикл полового размножения включает чередование гаплоидных поколений клеток, каждая из которых имеет одинарный набор хромосом, с диплоидными поколениями, где клетки обладают двойным хромосомным набором (рис. 15-2). Смешивание геномов происходит благодаря слиянию двух гаплоидных клеток, из которых образуется одна диплоидная. В свою очередь новые гаплоидные клетки образуются из диплоидных в результате деления особого типа, называемого мейозом. В процессе генетической рекомбинации в мейозе парные хромосомы обмениваются ДНК, после чего новые их комбинации расходятся в разные клетки, которые теперь содержат одинарные наборы хромосом (см. разд. 15.2.2). В результате каждая клетка нового гаплоидного поколения получает новое сочетание генов, происходящих частично от одной родительской клетки предыдущего гаплоидного поколения и частично от другой. Таким образом, благодаря циклам, включающим гаплоидную фазу, слияние гамет, диплоидную фазу и мейоз, распадаются старые комбинации генов и создаются новые.
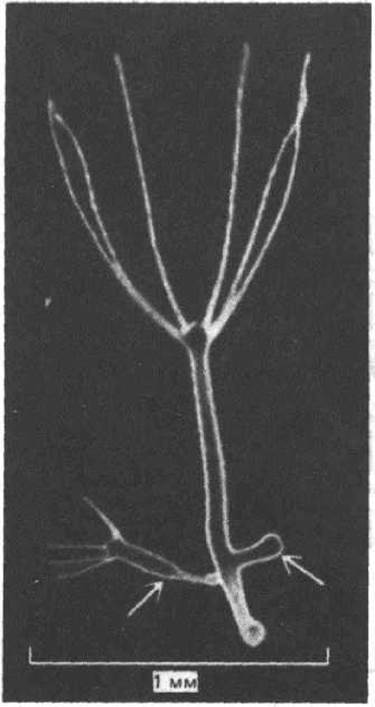
Рис. 15-1. Гидра, от которой отпочковываются две новые особи (указаны стрелками). Потомки генетически идентичны родительскому организму; они в конце концов, отделяются и переходят к независимому существованию. (С любезного разрешения Matai Hornbruch.)
15.1.1. У многоклеточных животных диплоидная фаза бывает сложной и продолжительной, а гаплоидная - простой и кратковременной
В ходе полового цикла клетки размножаются путем обычного митотического деления - чаще всего во время диплоидной фазы (см. разд. 13.5). Исключение составляют некоторые простые организмы, например дрожжи (путем митоза у них размножаются только гаплоидные клетки, диплоидная же клетка, образовавшись, сразу переходит к мейозу), а также растения, хотя и не в столь яркой форме; у последних митотические деления происходят и в гаплоидной, и в диплоидной фазах. При этом у всех растений, за исключением наиболее примитивных, гаплоидная фаза очень короткая и простая, тогда как диплоидная представлена длительным периодом развития и роста. Почти у всех многоклеточных животных, и в том числе у всех позвоночных, гаплоидная фаза еще короче. Практически весь свой жизненный цикл они проводят в диплоидном состоянии; гаплоидные клетки живут очень недолго, они совсем не делятся и специально приспособлены для полового слияния (рис. 15-3).
Гаплоидные клетки, которые сливаются при оплодотворении, называются гаметами. В типичном случае образуются гаметы двух типов: крупные неподвижные яйцеклетки (или яйца) и мелкие, способные передвигаться спермии (или сперматозоиды) (рис. 15-4). Во время диплоидной фазы, начинающейся сразу после слияния гамет, клетки размножаются и специализируются, образуя сложный многоклеточный организм. У большинства животных (но не растений) полезно различать клетки зародышевой линии (зачаткового пути), от которых берет начало следующее поколение гамет, и соматические клетки, образующие весь остальной организм и не оставляющие потомства. В некотором смысле соматические клетки нужны только для того, чтобы способствовать выживанию и размножению клеток зачаткового пути (половых клеток).
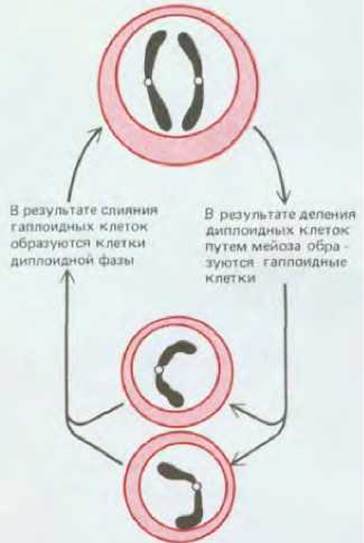
Рис. 15-2. Жизненный цикл организма, размножающегося половым путем, включает чередование диплоидного поколения клеток с гаплоидным.
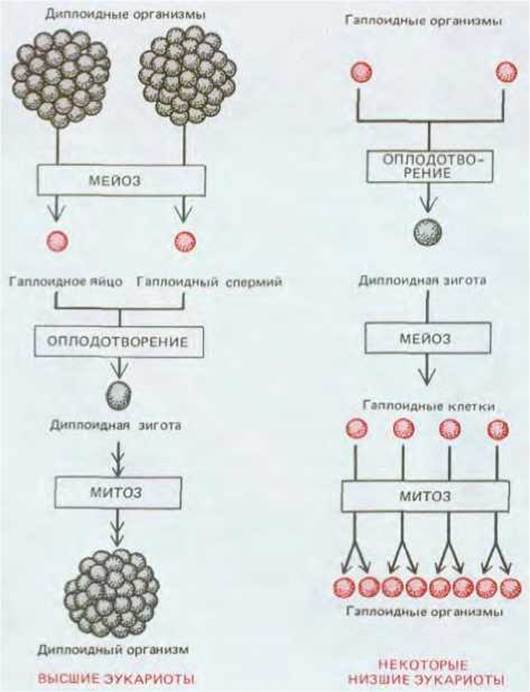
Рис. 15-3. Эта схема показывает, как размножаются в диплоидной фазе клетки высших эукариот, образуя многоклеточный организм, в котором гаплоидными становятся только гаметы. Напротив, у некоторых низших эукариот размножаются именно гаплоидные клетки, а единственной диплоидной клеткой является зигота, которая существует очень недолго после оплодотворения. Гаплоидные клетки выделены цветом
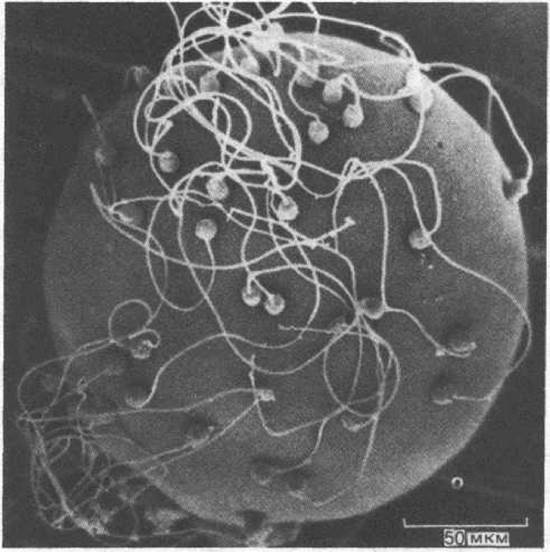
Рис. 15-4. Яйцеклетка двустворчатого моллюска с многочисленными спермиями, прикрепившимися к ее поверхности. Микрофотография получена с помощью сканирующего электронного микроскопа. (С любезного разрешения David Epel.)
15.1.2. Половое размножение делает организмы конкурентоспособными в условиях изменчивости окружающей среды
Аппарат полового размножения сложен, и средства, «затрачиваемые» на него, очень велики. Какие же преимущества он дает, и почему он выработался в процессе эволюции? При наличии генетической рекомбинации родительские особи производят потомков, которые будут отличаться от них самым непредсказуемым образом, причем среди новых случайных сочетаний генов, по крайней мере, половина может оказаться хуже родительских генотипов. Но если это так, то почему половое размножение должно быть выгоднее бесполого, при котором потомки будут сохранять все родительские гены? Хотя для специалистов по генетике популяций этот вопрос все еще до конца не ясен, основной вывод, по-видимому, состоит в том, что перетасовка генов в процессе полового размножения способствует выживанию вида при изменении условий среды. Если родительская особь производит много потомков с самыми разнообразными комбинациями генов, имеется больше шансов на то, что хотя бы один потомок окажется хорошо приспособленным для будущих жизненных обстоятельств, какими бы они ни были. Для объяснения преимуществ полового размножения в борьбе за существование было предложено много гипотез. Одна из них дает представление о том, какими могли быть первые этапы эволюции полового размножения. Ход эволюции в значительной мере зависит от мутаций, которые изменяют существующие гены, образуя вместо них новые аллели (варианты) этих генов. Предположим, что у двух особей в некоторой популяции возникли благоприятные мутации, затрагивающие разные генетические локусы, а значит, и разные функции.
У бесполого вида каждая из этих особей даст начало клону мутантных потомков, и два новых клона будут конкурировать до тех пор, пока один из них не одержит верх. Один из благоприятных аллелей, появившихся благодаря мутациям, будет, таким образом, распространяться, тогда как другой в конце концов исчезнет. Теперь представим себе, что один из исходных мутантов обладает генетически обусловленной особенностью, позволяющей ему время от времени включать в свой геном гены из других клеток. В условиях борьбы за существование приобретение генов у клеток конкурирующего клона равносильно созданию клетки, несущей все благоприятные мутации. Такая клетка будет обладать наибольшей приспособленностью, и полученные ею преимущества обеспечат распространение в популяции особенности, позволяющей включать в свой геном гены других клеток. Естественный отбор будет благоприятствовать такому примитивному половому размножению.
Какими бы ни были истоки полового размножения, поразительно то, что эволюция практически всех сложных организмов, доживших до наших дней, протекала благодаря чередованию многих поколений, размножавшихся половым путем. Несмотря на обилие бесполых организмов, они, по-видимому, остались весьма примитивными. Почему? Возможно, ответ состоит в том, что половое размножение создает особые возможности для генетического обновления, ведущего к развитию сложных организмов. Обо всем этом мы будем говорить дальше.
15.1.3. Новые гены появляются в результате дупликаций и дивергенции
Для эволюции сложного организма требуется нечто большее, чем улучшение уже имеющихся генов: нужны новые гены для осуществления новых функций. Как они появляются?
Многие белки многоклеточного животного могут быть сгруппированы в семейства: коллагены, глобины, сериновые протеазы и т. п. Белки одного семейства близки как по своей функции, так и по аминокислотной последовательности. Вряд ли можно сомневаться в том, что гены белков каждого такого семейства произошли от единственного предкового гена в результате процессов дупликации и дивергенции (разд. 10.5.3). Разные члены одного семейства белков часто бывают характерны для различных тканей тела, где они выполняют похожие, но несколько различающиеся функции. Создание новых генов благодаря дивергенции и специализации имеющихся играло, очевидно, решающую роль в эволюции сложных многоклеточных организмов. В этом отношении диплоидные организмы обладают важным преимуществом: у них имеется добавочная копия каждого гена, и эта копия может мутировать и служить исходным материалом для создания чего-то нового. Гаплоидные виды не могут так же легко вступать на путь, ведущий к увеличению и усложнению генома. Чтобы механизм этих процессов стал ясен, нам нужно будет несколько подробнее рассмотреть связь между половым размножением и диплоидией.
15.1.4. Половое размножение сохраняет диплоидность у диплоидных видов
У диплоидного организма имеются две копии каждого гена - по одной от каждого из родителей; однако для выживания и нормальной жизнедеятельности в большинстве случаев бывает достаточно одной копии. Мутация, нарушающая функцию жизненно важного гена, для гаплоидного организма летальна, но она может оказаться безвредной
для диплоида, если затронута лишь одна из двух копии гена, т. е. если организм гетерозиготен по мутации. Как правило, в геномах диплоидных организмов содержится много рецессивных деталей. Нередко гетерозиготные особи обладают несколько пониженной приспособленностью, поэтому распространение таких аллелей затруднено. Однако даже в том случае, когда приспособленность гетерозигот не снижена, на частоту появления в популяции рецессивных леталей накладывает ограничение половое размножение (рис. 15-5). Если обе родительские особи несут рецессивную летальную мутацию в одном и том же гене, их потомок может унаследовать две мутантные копии этого гена и не получить ни одной нормальной; такой гомозиготный организм погибнет, и вместе с ним будут утрачены мутантные копии гена. Чем больше распространен в популяции летальный аллель, тем быстрее он будет элиминироваться. В результате устанавливается равновесие между скоростью элиминации летального аллеля и скоростью его образования за счет новых мутаций. При равновесии рецессивный летальный аллель встречается в популяции достаточно редко (хотя и значительно чаще, чем это было бы у гаплоидного организма): подавляющее большинство особей будут иметь две функциональные копии гена. Сходным образом обстоит дело и с теми рецессивными мутациями, которые просто вредны (ведут к снижению численности потомства), но не летальны. В общем случае при наличии полового размножения и генетической рекомбинации отбор обеспечивает такое положение, при котором у большинства особей в большей части генных локусов остаются две функционально взаимозаменяемые копии гена, т. е. геном сохраняет диплоидность.
Для сравнения рассмотрим популяцию, первоначально состоящую из диплоидных особей, которые размножаются бесполым способом. В отсутствии генетической рекомбинации ничто не препятствует тому, чтобы две копии каждого гена эволюционировали различными путями. Вредные рецессивные мутации будут накапливаться в геноме до тех пор, пока диплоидность не сменится состоянием, при котором общее количество ДНК остается прежним, но сохраняется лишь одна функционирующая копия каждого из первоначальных необходимых генов. Организм становится «функционально гаплоидным». Представление о промежутках времени, необходимых для подобных эволюционных изменений, можно получить, рассматривая эволюцию чукучановых рыб1. Эти рыбы происходят от предков, у которых около 50 млн. лет назад произошла полная дупликация прежде диплоидного генома и они стали, таким образом, тетраплоидами. Подсчитано, что около 50% «лишних» пар генов, кодирующих белки, утратили с тех пор свое функциональное значение.
1) Чукучановые (Catostomidae) - семейство пресноводных рыб отряда карпообразных. - Прим. ред.
15.1.5. Диплоидный вид обладает лишней копией каждого гена, способной мутировать и выполнять после этого новую функцию
Большинство мутаций являются вредными, поскольку они нарушают функцию гена, которая уже была оптимизирована в ходе естественного отбора. Однако иногда может произойти мутация, которая модифицирует имеющийся ген таким образом, что он приобретает новую полезную функцию. Как правило, такая мутация делает ген неспособным осуществлять свою первоначальную функцию и, если эта функция была жизненно необходимой, гаплоидный организм гибнет. Однако в диплоидном организме подобного рода мутация в одной из двух копий гена не просто терпима - она приносит пользу. Даже небольших преимуществ, которые получит организм благодаря новому мутантному гену, будет достаточно, чтобы перекрыть ущерб, нанесенный потерей одной из двух исходных копий гена: гетерозиготная особь будет извлекать пользу как из старой, так и из новой функции гена. Гомозиготы, имеющие две копии старого аллеля или две копии нового, окажутся менее приспособленными. В подобных случаях, когда имеется преимущество гетерозигот, мутантный ген быстро распространяется в диплоидной популяции с половым размножением - до тех пор, пока не будет достигнуто равновесие, при котором и старые, и новые аллели представлены с высокой частотой и доля гетерозиготных особей велика. Это явление называется сбалансированным полиморфизмом. Однако кое-чем приходится расплачиваться: при скрещивании двух гетерозигот значительная часть потомков в соответствии с обычными законами Менделя окажется гомозиготной и поэтому хуже приспособленной. Но такое положение не будет сохраняться вечно - из него есть выход.

Рис. 15-5. Из этой схемы видно, каким образом при половом размножении диплоидные организмы сохраняют диплоидность в ходе эволюции. Для простоты рассматриваются только летальные рецессивные мутации. Аналогичным образом обстоит дело и с вредными рецессивными мутациями.
15.1.6. Диплоидный вид может быстро обогащать свой геном, приобретая новые гены
Время от времени у всех организмов происходит спонтанное удвоение генов: хромосома, содержащая одну копию гена G, в результате ошибки в репликации ДНК дает начало хромосоме, в которую входят уже две копии этого гена, расположенные одна за другой. Такие дупликации сами по себе не дают никаких преимуществ и встречаются, как правило, у очень немногих особей. Предположим, однако, что дупликация произошла в локусе, содержащем полезный мутантный аллель G*, который с высокой частотой присутствует в популяции в связи с отбором в пользу гетерозигот и сосуществует в геноме с исходным аллелем G (рис. 15-6). Тогда велика вероятность того, что в диплоидной клетке, содержащей хромосому GG (несущую дупликацию), ее гомолог будет содержать аллель G*, так что получится генотип GG/G*. Затем в результате генетической рекомбинации в мейозе (см. ниже) могут образоваться гаметы с генотипом GG*. В этих гаметах исходный ген G и мутантный G*. расположенные один за другим, не будут уже двумя аллелями, конкурирующими за один и тот же локус; теперь это два отдельных гена, каждый из которых занимает собственный локус. Такая комбинация выгодна, и она станет быстро распространяться, пока, наконец, вся популяция не будет состоять из гомозигот GG*/GG* (см. рис. 15-6). Преимущество особей с таким генотипом состоит не только в обладании обоими генами - старым G и новым G*, но и в том, что они могут передавать это преимущество всем своим потомкам.
Таким образом, у диплоидного вида с половым размножением могут возникать новые гены в результате мутаций в добавочных копиях имеющихся генов; эти новые гены могут распространиться в популяции благодаря отбору в пользу гетерозигот, причем не будут потеряны и исходные гены; и наконец, новые гены могут дополнительно включаться в геном в результате процессов дупликации генов и генетической рекомбинации. Такая последовательность событий возможна только у диплоидных видов. Обогащение генома у гаплоидного вида связано с большими трудностями. Если в процессе приобретения нового гена вид должен сохранить и старый ген, то ему придется ждать возникновения нужной мутации у одной из очень немногих особей, у которых уже произошла дупликация соответствующего локуса. А поскольку и мутации, и дупликации в определенном локусе происходят очень редко, гаплоидному виду приходится дожидаться совпадения этих событий чрезвычайно долго (рис. 15-7). Детальные расчеты показывают, что обычно (за исключением ситуаций, когда частота дупликаций генов очень велика) диплоидный организм способен расширять свой геном и добавлять к нему новые гены с новыми функциями в сотни или даже тысячи раз быстрее, чем это происходит у гаплоидного организма. Наиболее ярко различие выражено в отношении тех мутаций, которые происходят с низкой частотой, а именно такие редкие мутации необходимы для обновления генома.
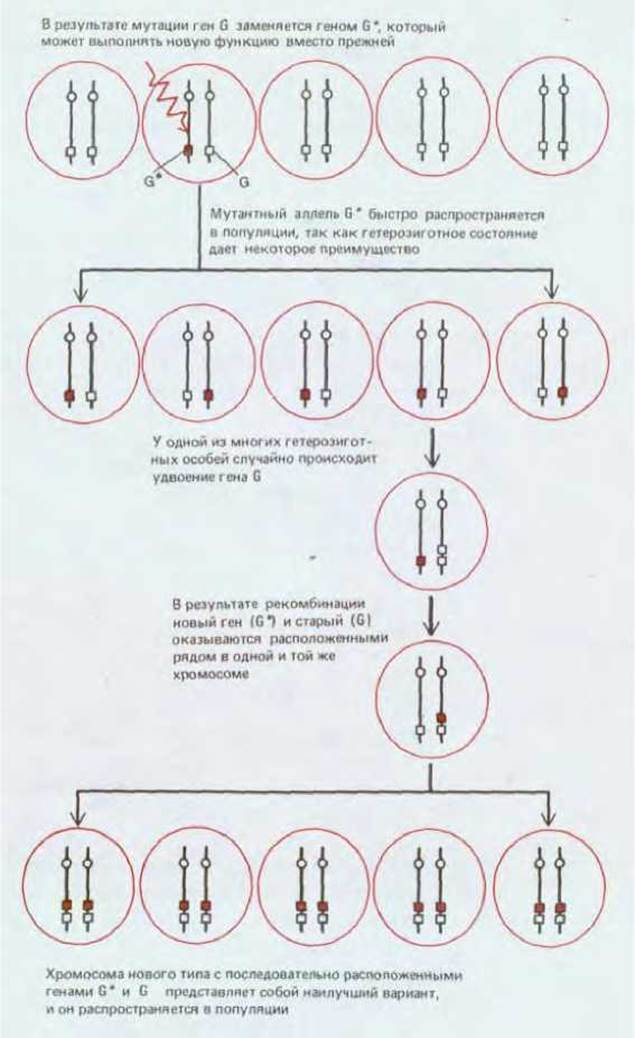
Рис. 15-6. Возникновение нового гена (G*) по схеме «мутация → распространение → дупликация» в процессе полового размножения диплоидного организма.
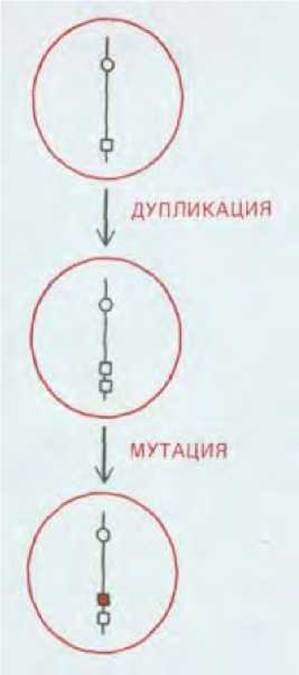
Рис. 15-7. Образование нового гена у гаплоидного организма. Такая последовательность событий кажется гораздо более простой, чем та, которая показана на рис. 15-6, но требует для реализации значительно больше времени.
Итак, половое размножение идет «рука об руку» с диплоидностью, которая в свою очередь обеспечивает особо благоприятные условия для создания более крупного, более сложного и более гибкого генома. Конечно, эволюция может протекать по-разному, и указанный нами путь дупликаций и дивергенции генов, безусловно, далеко не единственный. Тем не менее, половое размножение, по-видимому, оказало огромное влияние на истоки и способы распространения в популяции генетических изменений, сделав возможным появление столь сложных организмов, как мы сами.
Теперь можно перейти к детальному описанию клеточных механизмов полового процесса. В последующих разделах сначала будет рассмотрен мейоз, в ходе которого осуществляется генетическая рекомбинация и из диплоидных клеток образуются гаплоидные гаметы; затем мы обратимся к самим гаметам и, наконец, познакомимся с процессом оплодотворения, при котором гаметы сливаются, образуя новый диплоидный организм.
Заключение
При половом размножении происходит циклическое чередование диплоидного и гаплоидного состояний: диплоидная клетка делится путем мейоза, порождая гаплоидные клетки, а гаплоидные клетки попарно сливаются при оплодотворении и образуют новые диплоидные клетки. Во время этого процесса происходит перемешивание и рекомбинация геномов, в результате чего появляются особи с новыми наборами генов. Высшие растения и животные большую часть жизненного цикла проводят в диплоидной фазе, а гаплоидная фаза у них очень короткая. Вероятно, процесс эволюции благоприятствовал половому размножению, так как случайная генетическая рекомбинация увеличивала шансы организмов на то, что хотя бы некоторые из их потомков выживут в непредсказуемо изменчивом мире. Половой процесс необходим также для поддержания диплоидности; он способствует созданию условий для быстрой выработки новых генов у высших растений и животных.