Основы молекулярной биологии. Часть 1: Молекулярная биология клетки - А.Н. Огурцов 2011
Окисление глюкозы и жирных кислот
Цитратный цикл
Финальный этап окисления глюкозы представляет собой цикл из девяти реакций, который имеет несколько названий - цитратный цикл, или цикл трикарбоновых кислот, или цикл Кребса (рисунок 157).
Каждая ацетильная группа, вошедшая в этот цикл в составе ацетил-СоА, превращается в две молекулы СO2.
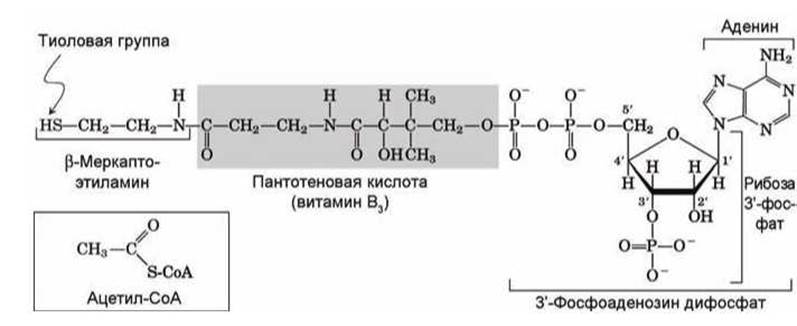
Рисунок 156 - Схема кофермента коэнзим А (СоА). Вставка - схема ацетил-СоА
Цикл начинается с образования шестиуглеродной молекулы лимонной кислоты (citric acid) в результате объединения на ферменте цитрат-синтаза двухуглеродной ацетильной группы ацетил-СоА с четырёхуглеродной молекулой оксалоацетата (рисунок 157).
Двухэтапная конверсия цитрата в изоцитрат (реакции 2 и 3 на рисунке 157) проходят на одном ферменте аконитат-гидратаза, причем промежуточный продукт цис-аконитат остается во время реакции связанным с ферментом.
В реакциях 4 и 5 высвобождается по одной молекуле СО2.
В реакции 5, которая катализируется ферментом а-кетоглутарат-дегидрогеназа, также восстанавливается НАД⊕ до НАДН. Кроме того, в реакциях 4 и 9 также происходит такое же восстановление, в результате в каждом цикле образуется 3 молекулы НАДН.
В реакции 7 два электрона и два протона переносятся на ФАД, восстанавливая кофермент до ФАДН2.
В реакции 6 (фермент сукцинат-СоА-лигаза) гидролиз высокоэнергетичной тиоэфирной связи в сукцинил-СоА сопряжен с синтезом ГТФ (из ГДФ) - второй пример субстратного фосфорилирования.
Субстратное фосфорилирование реализуется только в двух случаях - в реакции 6 образования ГТФ в цитратном цикле (рисунок 157) и в реакциях 7 и 10 гликолиза (рисунок 150).
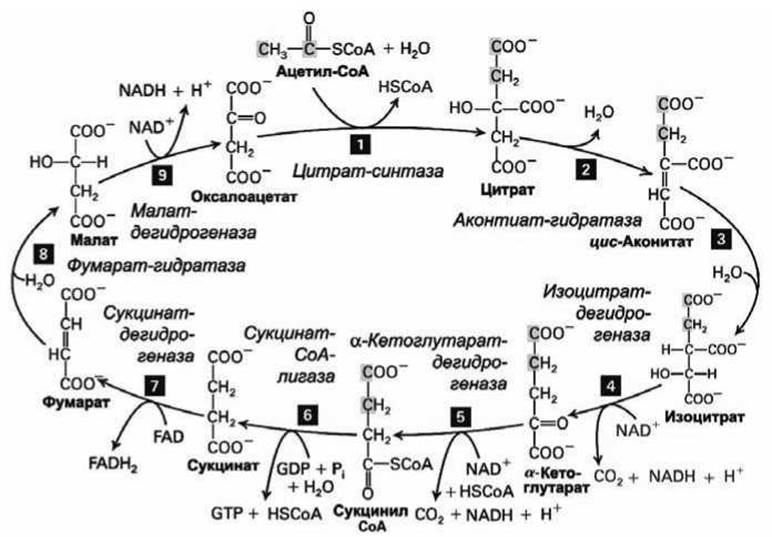
Рисунок 157 - Схема витратного цикла. Метаболиты подписаны жирным шрифтом, а ферменты - курсивом
В последней реакции 9 (фермент малатдегидрогеназа) вновь образуется оксалоацетат и цикл может начаться снова.
Примечательно, что молекулярный кислород О2 не участвует в цитратном цикле.
Все метаболиты и шесть из восьми ферментов цитратного цикла являются водорастворимыми и располагаются в матриксе митохондрии.
Сукцинат дегидрогеназа (реакция 7) и а-кетоглутаратдегидрогеназа (реакция 5) являются мембранными белками внутренней мембраны с активными центрами ориентированными в матрикс митохондрии.
Экспериментально показано, что шесть немембранных ферментов образуют в матриксе большой мулытибелковый комплекс (конвейер ферментов), в котором продукты предыдущего фермента непосредственно передаются следующему ферменту без диффузии в матриксе.
Суммарно, на одну молекулу глюкозы гликолиз и цитратный цикл синтезируют 6 молекул СО2, 10 молекул НАДН и 2 молекулы ФАДН2 (таблица 8). И хотя в этих реакциях формируются ещё четыре высоко-энергетичные фосфоангидридные связи в составе молекул АТФ и ГТФ, на это уходит только незначительная часть энергии, выделяющейся при полном окислении глюкозы. Основная часть этой энергии аккумулируется в восстановленных коферментах НАДН и ФАДН2.
Таблица 8 - Число молекул, суммарно синтезируемых при гликолизе и в цитратном цикле
|
Реакция |
Число синтезируемых молекул |
||
|
СО2 |
НАД⊕ восстановлено до НАДН |
ФАД восстановлено до ФАДН2 |
|
|
1 глюкоза → 2 пирувата (гликолиз) |
0 |
2 |
0 |
|
2 пирувата → 2 ацетил-СоА (пируват-дегидрогеназа) |
2 |
2 |
0 |
|
2 ацетил-СоА → 4 СО2 (цикл Кребса) |
4 |
6 |
2 |
|
Всего |
6 |
10 |
2 |
Синтез же большинства молекул АТФ, которые образуются в процессах аэробного окисления, сопряжен с процессами окисления НАДН и ФАДН2 молекулярным кислородом О2 в пошаговой последовательности реакций, которые в совокупности называются дыхательной цепью или цепью транспорта электронов.
И хотя молекулярный кислород не используется в цитратном цикле, в отсутствии кислорода цитратный цикл замедляется и останавливается по мере исчерпания запаса НАД⊕ и ФАД. Прежде чем рассматривать цепи транспорта электронов, мы ознакомимся с системами регенерации НАД⊕ в цитозоле и окисления жирных кислот до СО2.