Основы молекулярной биологии. Часть 1: Молекулярная биология клетки - А.Н. Огурцов 2011
Генерация протондвижущей силы и синтез АТФ
Цепь переноса электронов
Свободная энергия, которая выделяется в результате гликолиза и в цитратном цикле, сосредотачивается, главным образом, в молекулах восстановленных коферментов НАДН и ФАДН2. В процессе клеточного дыхания электроны высвобождаются с НАДН и ФАДН2 и передаются на O2, образуя Н2O в суммарных реакциях
![]()
Изменение свободной энергии в этих сильно экзергонических реакциях равно ∆G = -52,6 ккал/моль для НАДН и ∆G = -43,4 ккал/моль для ФАДН2. Суммарно окисление одной молекулы глюкозы в гликолизе и цитратном цикле дает 10 молекул НАДН и 2 молекулы ФАДН2, поэтому окисление этих коферментов даст
∆G = 10х(-52,6) + 2х(-43,4) = — 613 ккал/моль.
Таким образом, из всей свободной энергии, которая может выделится при полном окислении глюкозы (-680 ккал/моль), более 90% переходит в коферменты НАДН и ФАДН2 при их восстановлении.
Количество свободной энергии, которое выделяется при окислении одной молекулы НАДН или ФАДН2 кислородом, достаточно для синтеза нескольких молекул АТФ из АДФ и Рi (эта эндергоническая реакция потребляет всего ∆G = +7,3 ккал/моль). Митохондрия максимально использует эту энергию, перенося электроны от НАДН и ФАДН2 по цепи электронных переносчиков (рисунок 162), все, кроме одного из которых, являются интегральными белками внутренней мембраны митохондрии.
Такой пошаговый перенос электронов по дыхательной цепи (по цепи переноса электронов) обеспечивает контролируемое выделение энергии небольшими порциями и превращение этих порций энергии в протондвижущую силу (рисунок 154(2)).
Главными компонентами дыхательной цепи являются четыре больших мультибелковых комплекса:
|
Комплекс I) |
НАДН-СоQ-редуктаза, |
|
Комплекс II) |
Сукцииат-СоQ-редуктаза, |
|
Комплекс III) |
CoQH2-цитохром с редуктаза, |
|
Комплекс IV) |
Цитохром с оксидаза. |
Кофермент Q (CoQ) и цитохром с переносят электроны между этими комплексами. Той энергии, которая выделяется при релаксации электрона внутри комплексов I, III и IV, хватает для переноса протонов через мембрану белками этих комплексов (их называют протонными насосами или протонными помпами), и в результате такого трансмембранного транспорта создается градиент концентрации протонов на внутренней мембране митохондрии.
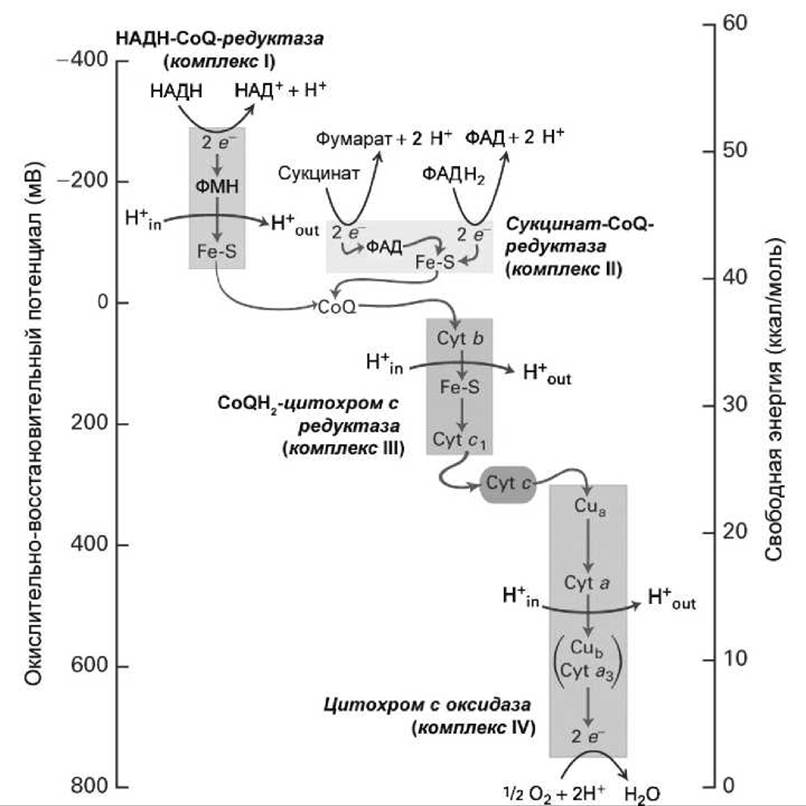
Рисунок 162 - Изменение окислительно-восстановительного потенциала и свободной энергии в ходе последовательного переноса электронов по дыхательной цепи
Поскольку внешняя мембрана митохондрии проницаема для протонов, а внутренняя — нет, то в результате такого трансмембранного транспорта протонов pH митохондриального матрикса растет (а концентрация протонов в матриксе снижается) по отношению к цитозолю и межмембранному пространству митохондрии. Матрикс становится отрицательно заряженным по отношению к цитозолю и на внутренней мембране митохондрии формируется мембранный потенциал.
В результате свободная энергия, которая выделяется в ходе окисления НАДН и ФАДН2, запасается и в виде градиента концентрации протонов, и в форме электрического потенциала на внутренней мембране митохондрии (суммарно это и называют протондвижущей силой).
Движение протонов обратно через внутреннюю мембрану митохондрии под действием этой протондвижущей силы сопряжено с синтезом АТФ из АДФ и Pi ферментом АТФ-синтаза (рисунок 154(3)).
В состав комплекса I входит флавиновый кофермент (простетическая группа) флавинмононуклеотид, ФМН (FMN), в котором, так же как и в ФАД (рисунок 163(a)), активной группой является флавин (изоаллоксазин) (см. рисунок 155).
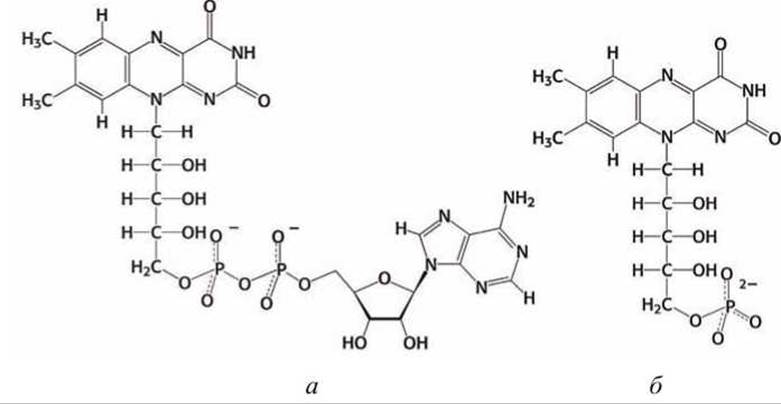
Рисунок 163 - Флавиновые коферменты: а - флавинадениндинуклеотид (ФАД), б - флавинмононуклеотид (ФМН)
Флавин имеет сопряженную систему из трех колец, которая может при восстановлении принимать два электрона и два протона. В флавин-мононуклеотиде (ФМН) к флавину присоединен фосфорилированный полиол рибит (рисунок 163(6)).
Окислительным фосфорилированием называется процесс синтеза АТФ из АДФ и Рi за счет релаксации энергии электронов при их переносе от НАДН и ФАДН2 к О2. Окислительное фосфорилирование является главным источником АТФ в аэробных нефотосинтетических клетках.