Основы молекулярной биологии. Часть 2: Молекулярные генетические механизмы - А.Н. Огурцов 2011
Вирусы
Строение вирусного капсида
Нуклеиновая кислота вириона заключена в белковую оболочку - капсид - которая состоит из тождественных белков или же из множественных повторений небольшого числа разных белков, которые кодируются одним виральным геном.
Именно такая организация позволяет вирусу кодировать информацию для создания относительно большого капсида с помощью очень небольшого числа генов.
Такая эффективность использования генетической информации критически важна, поскольку очень ограниченный объём капсида может вместить в себя очень ограниченный набор генов.
Капсид вместе с заключенной в него нуклеиновой кислотой (РНК или ДНК) называют нуклеокапсидом.
Существует два основных способа организации нуклеокапсида.
Первый способ состоит в том, что идентичные капсидные белки формируют спиральную структуру, которая окружает и защищает вирусную РНК (или ДНК), которая расположена вдоль спиральной впадины внутри белковой трубки (рисунок 63).
Вирусы, имеющие такую спиралевидную структуру (как, например, вирус табачной мозаики), имеют стержнеобразную форму.
Второй основной способ формирования капсида - икосаэдрический.
Икосаэдр - это сфероподобная поверхность, построенная из 20 идентичных равносторонних треугольников. Число и расположение белков на гранях икосаэдрических или квази-сферических вирусных капсидов различается в зависимости от размера вириона.
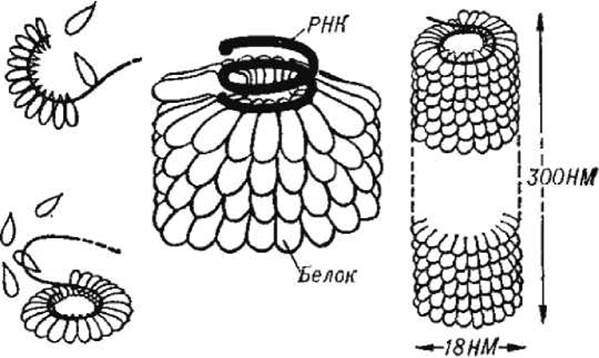
Рисунок 63 - Структура и самосборка нуклеокапсида вируса табачной мозаики из белковых субъединиц и молекул РНК
В малых квази-сферических вирусах каждая из 20 треугольных граней построена тремя идентичными копиями капсидных белков. Весь капсид, таким образом, построен из 60 белков. И каждый капсидный белок находится в эквивалентном положении и имеет эквивалентное окружение (рисунок 64). Каждая вершина икосаэдра имеет ось симметрии пятого порядка.
В больших квази-сферических вирусах каждая грань икосаэдра образована более чем тремя белковыми субъединицами.
В результате число соседей у каждой белковой субъединицы зависит от того, расположена она у вершины икосаэдра или нет. Положения белков в капсиде уже не эквивалентны, а квазиэквивалентны.
Так, например, каждая грань икосаэдра аденовируса образована четырьмя субъединицами (рисунок 65). Поэтому вершины икосаэдра имеют симметрию пятого порядка, а места соединения граней — симметрию шестого порядка.
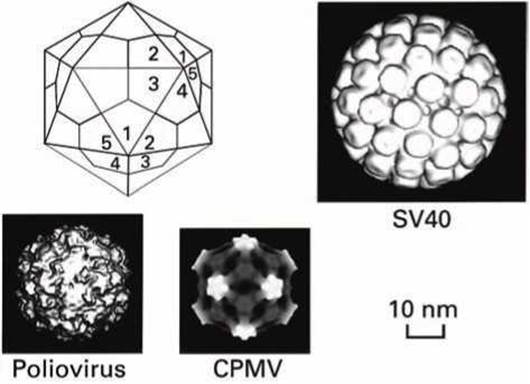
Рисунок 64 - Структура малых икосаэдрическнх вирусов - полиовирус (РНК- вирус человека), мозаичный вирус вигны (cowpea mosaic virus, CPMV; РНК-вирус; вигна - род одно- и многолетних растений семейства бобовых), малый вирус 40 (small virus 40, SV40, ДНК-вирус обезьян)
В малых вирусах, таких как полиовирус, впадина, которая окружает каждую вершину икосаэдра, взаимодействует с рецепторами на поверхности инфицируемой клетки.
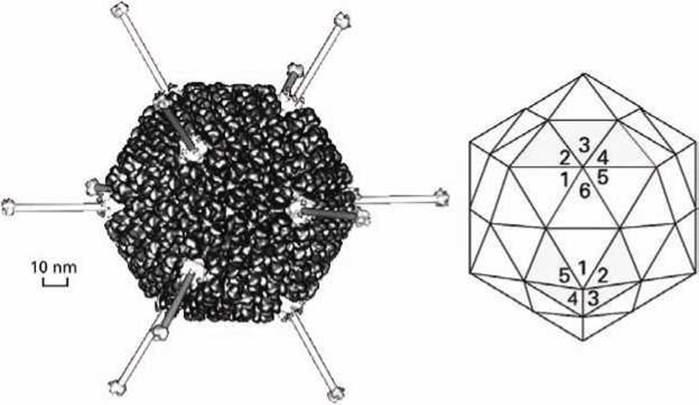
Рисунок 65 - Структура большого икосаэдрического аденовируса
В больших вирусах (например, в аденовирусе) с поверхностными рецепторами инфицируемой клетки взаимодействуют длинные фибриллярные белки, выступающие над поверхностью нуклеокапсида.
Во многих ДНК-бактериофагах виральная ДНК расположена в икосаэдрической "головке", которая соединена с трубчатым каналом ("хвостом") (рисунок 62(д)). В процессе инфицирования вирусные белки на конце хвоста связываются с рецепторами на поверхности инфицируемой клетки, а затем ДНК вируса спускается из головки через трубчатый канал хвоста в цитоплазму инфицируемой клетки.
У некоторых вирусов симметричный нуклеокапсид окружен внешней оболочкой (envelope) - липопротеидной мембраной, которая состоит в основном из фосфолипидного бислоя и одного-двух типов вирусных белков (рисунок 66).
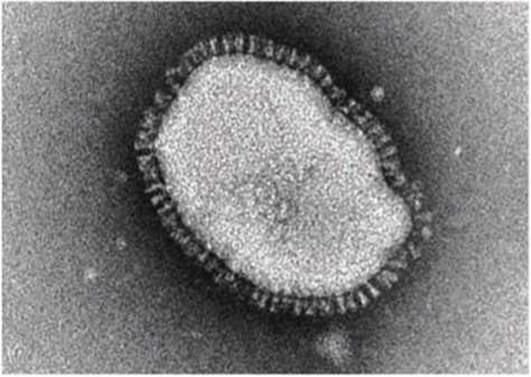
Рисунок 66 - Электронная микрофотография вируса гриппа
Фосфолипиды липопротеидной оболочки вируса идентичны тем фосфолипидам, из которых состоит плазматическая мембрана инфицируемой клетки, а сама вирусная оболочка, на самом деле, формируется из плазматической мембраны инфицируемой клетки во время цикла размножения вируса, но при этом ещё и содержит большое число вирусных белков.
На рисунке 66 представлена электронная микрофотография вируса гриппа, на которой видны выступающие над поверхностью оболочки вируса белки: нейраминидаза (тетрамерный белок) или гемаглуттинин.
Исследование вирусов. Число вирусов в образце может быть определено с использованием метода бляшек. Разбавленный раствор, содержащий вирусы помещают на поверхность чашки Петри, содержащей инфицируемые клетки. После инфицирования покрывают слой клеток агаром для фиксации и инкубируют. Затем подсчитывают количество повреждений, называемых дисками или бляшками (plaque) (или негативными колониями), которые вырастают через определённое время (рисунок 67).
Бляшка развивается в чашке Петри в месте, где один вирион первоначально инфицировал одну клетку. Вирус размножается в этой клетке, а затем разрушает её, высвобождая много дочерних вирионов, которые инфицируют соседние клетки в чашке. После нескольких циклов инфицирования образуется достаточно разрушенных клеток, которые образуют видимое пятно (бляшку) в слое оставшихся неинфицированных клеток.
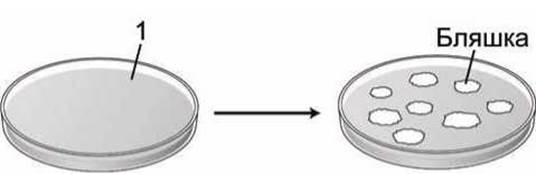
Рисунок 67 - Определение числа инфицированных частиц методом бляшек; 1 - посев клеток
Поскольку дочерние вирионы в бляшке развиваются от единственного родительского вируса, они формируют вирусный клон. Такая методика является стандартной при исследовании вирусов бактерий и животных (рисунок 68). Вирусы растений исследуют аналогично, считая количество локальных повреждений на листьях растений пораженных вирусом. Именно анализ вирусных мутантов, которые изолированы в бляшках, внес большой вклад в современный уровень понимания молекулярных клеточных процессов. Кроме того, метод бляшек является важным компонентом изоляции клонов бактериофага X, который используют для транспортировки сегментов клеточной ДНК в технологии рекомбинантных ДНК.
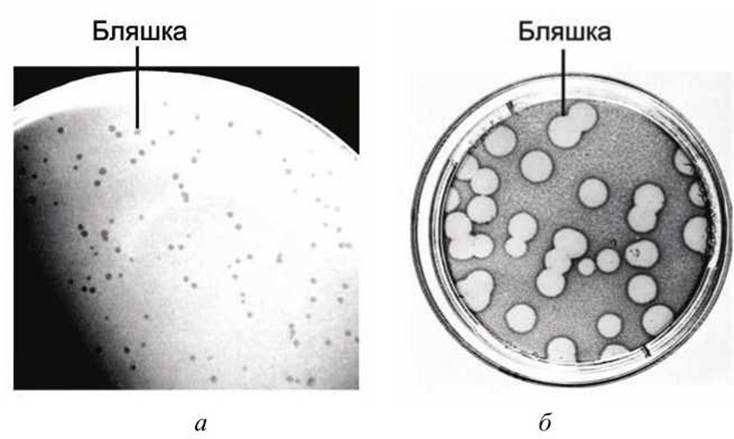
Рисунок 68 - Вирусные клоны: а - бляшки образованные бактериофагом X в клетках Е. coli; б - бляшки образованные полиовирусом в HeLa-клетках